
Effects of Intranasal Administration of Oxytocin and Vasopressin on Social Cognition and Potential Routes and Mechanisms of Action


{kind=link}
Abstract
:1. Introduction
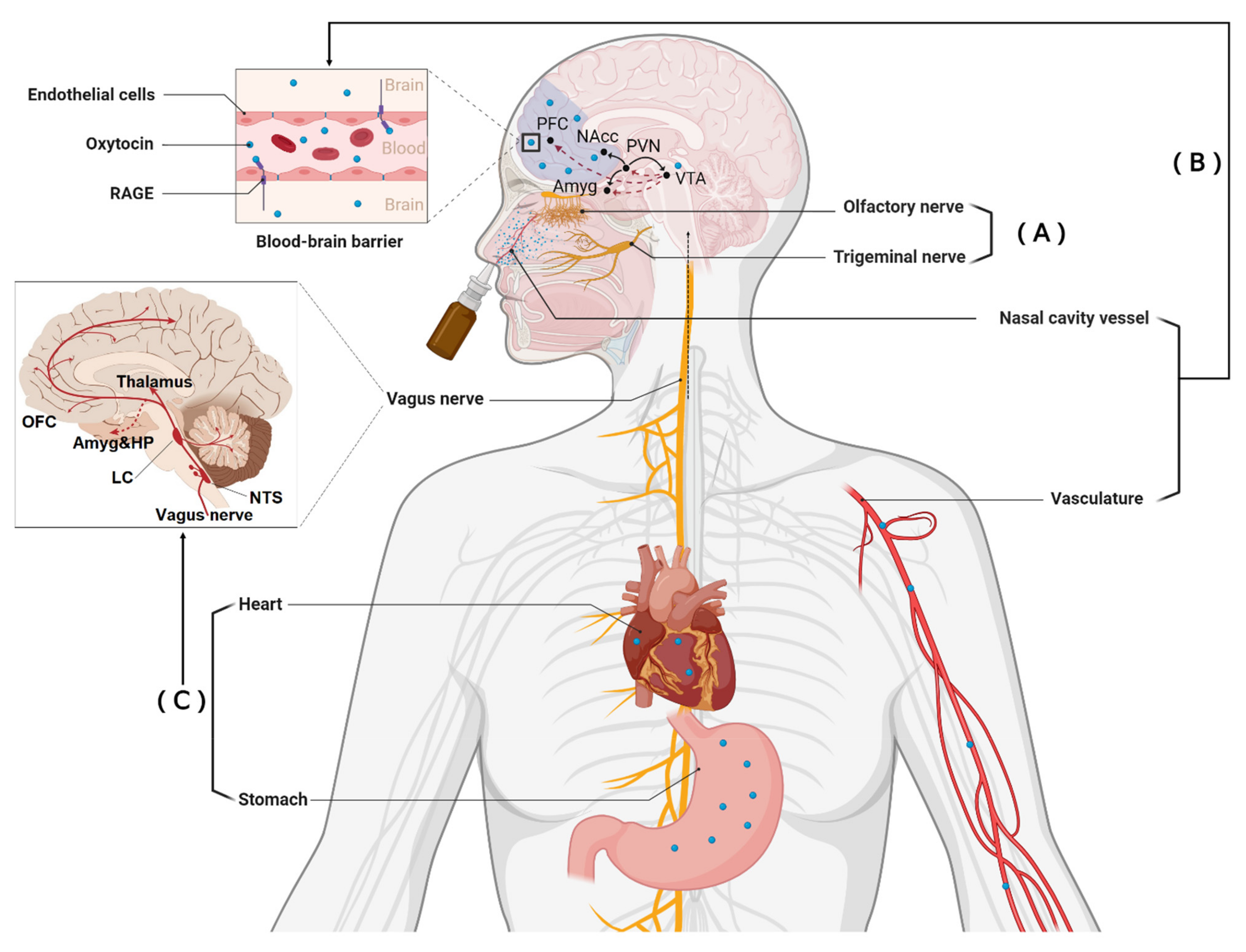
2. Different Routes Whereby Intranasal Oxytocin and Vasopressin May Produce Functional Effects
2.1. Direct Entry into the Brain
2.2. Indirect Entry into the Brain from the Peripheral Circulation
3. Acting on Receptors in Peripheral Organs to Influence the Brain via the Vagus
4. How Can We Distinguish between Functional Effects of Intranasal Administration Mediated via These Three Different Routes?
5. Characteristics of Oxytocin and Vasopressin Receptors
Effects of Dose Magnitude and Dose Frequency
6. Influence of Different Types of Intranasal Administration Device
7. Conclusion and Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kendrick, K.M.; Guastella, A.J.; Becker, B. Overview of human oxytocin research. Curr. Top. Behav. Neurosci. 2017, 35, 321–348. [Google Scholar]
- Patin, A.; Scheele, D.; Hurlemann, R. Oxytocin and interpersonal relationships. Behav. Pharmacol. Neuropept. Oxytocin 2017, 35, 389–420. [Google Scholar]
- Rae, M.; Lemos Duarte, M.; Gomes, I.; Camarini, R.; Devi, L.A. Oxytocin and vasopressin: Signalling, behavioural modulation and potential therapeutic effects. Br. J. Pharmacol. 2021. [Google Scholar] [CrossRef]
- Erdő, F.; Bors, L.A.; Farkas, D.; Bajza, Á.; Gizurarson, S. Evaluation of intranasal delivery route of drug administration for brain targeting. Brain Res. Bull. 2018, 143, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Lochhead, J.J.; Thorne, R.G. Intranasal delivery of biologics to the central nervous system. Adv. Drug Deliv. Rev. 2012, 64, 614–628. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.A.; Merkel, O.; Popp, A. Intranasal drug delivery: Opportunities and toxicologic challenges during drug development. Drug Deliv. Transl. Res. 2021, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Lindenberg, A.; Domes, G.; Kirsch, P.; Heinrichs, M. Oxytocin and vasopressin in the human brain: Social neuropeptides for translational medicine. Nat. Rev. Neurosci. 2011, 12, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Ma, X.; Geng, Y.; Zhao, Z.; Zhou, F.; Zhao, W.; Yao, S.; Yang, S.; Zhao, Z.; Becker, B.; et al. Intrinsic, dynamic and effective connectivity among large-scale brain networks modulated by oxytocin. Neuroimage 2021, 227, 117668. [Google Scholar] [CrossRef]
- Quintana, D.S.; Rokicki, J.; van der Meer, D.; Alnæs, D.; Kaufmann, T.; Córdova-Palomera, A.; Dieset, I.; Andreassen, O.A.; Westlye, L.T. Oxytocin pathway gene networks in the human brain. Nat. Commun. 2019, 10, 668. [Google Scholar] [CrossRef] [Green Version]
- Auyeung, B.; Lombardo, M.V.; Heinrichs, M.; Chakrabarti, B.; Sule, A.; Deakin, J.B.; Bethlehem, R.A.I.; Dickens, L.; Mooney, N.; Sipple, J.A.N.; et al. Oxytocin increases eye contact during a real-time, naturalistic social interaction in males with and without autism. Transl. Psychiatry 2015, 5, e507. [Google Scholar] [CrossRef]
- Le, J.; Kou, J.; Zhao, W.; Fu, M.; Zhang, Y.; Becker, B.; Kendrick, K.M. Oxytocin biases eye-gaze to dynamic and static social images and the eyes of fearful faces: Associations with trait autism. Transl. Psychiatry 2020, 10, 142. [Google Scholar] [CrossRef]
- Gordon, I.; Jack, A.; Pretzsch, C.M.; Vander Wyk, B.; Leckman, J.F.; Feldman, R.; Pelphrey, K.A. Intranasal oxytocin enhances connectivity in the neural circuitry supporting social motivation and social perception in children with autism. Sci. Rep. 2016, 6, 35054. [Google Scholar] [CrossRef] [Green Version]
- Greene, R.K.; Spanos, M.; Alderman, C.; Walsh, E.; Bizzell, J.; Mosner, M.G.; Kinard, J.L.; Stuber, G.D.; Chandrasekhar, T.; Politte, L.C.; et al. The effects of intranasal oxytocin on reward circuitry responses in children with autism spectrum disorder. J. Neurodev. Disord. 2018, 10, 12. [Google Scholar] [CrossRef]
- Kruppa, J.A.; Gossen, A.; Weiß, E.O.; Kohls, G.; Großheinrich, N.; Cholemkery, H.; Freitag, C.M.; Karges, W.; Wölfle, E.; Sinzig, J.; et al. Neural modulation of social reinforcement learning by intranasal oxytocin in male adults with high-functioning autism spectrum disorder: A randomized trial. Neuropsychopharmacology 2019, 44, 749–756. [Google Scholar]
- Le, J.; Zhang, L.; Zhao, W.; Zhu, S.; Lan, C.; Kou, J.; Zhang, Q.; Zhang, Y.; Li, Q.; Chen, Z.; et al. Infrequent intranasal oxytocin followed by positive social interaction improves symptoms in autistic children: A randomized clinical trial. medRxiv 2022. [Google Scholar] [CrossRef]
- Parker, K.J.; Oztan, O.; Libove, R.A.; Sumiyoshi, R.D.; Jackson, L.P.; Karhson, D.S.; Summers, J.E.; Hinman, K.E.; Motonaga, K.S.; Phillips, J.M.; et al. Intranasal oxytocin treatment for social deficits and biomarkers of response in children with autism. Proc. Natl. Acad. Sci. USA 2017, 114, 8119–8124. [Google Scholar] [CrossRef] [Green Version]
- Yatawara, C.J.; Einfeld, S.L.; Hickie, I.B.; Davenport, T.A.; Guastella, A.J. The effect of oxytocin nasal spray on social interaction deficits observed in young children with autism: A randomized clinical crossover trial. Mol. Psychiatry 2016, 21, 1225–1231. [Google Scholar] [CrossRef] [Green Version]
- Parker, K.J.; Oztan, O.; Libove, R.A.; Mohsin, N.; Karhson, D.S.; Sumiyoshi, R.D.; Summers, J.E.; Hinman, K.E.; Motonaga, K.S.; Phillips, J.M.; et al. A randomized placebo-controlled pilot trial shows that intranasal vasopressin improves social deficits in children with autism. Sci. Transl. Med. 2019, 11, 491. [Google Scholar] [CrossRef]
- MacDonald, E.; Dadds, M.R.; Brennan, J.L.; Williams, K.; Levy, F.; Cauchi, A.J. A review of safety, side-effects and subjective reactions to intranasal oxytocin in human research. Psychoneuroendocrinology 2011, 36, 1114–1126. [Google Scholar] [CrossRef]
- Pardridge, W.M. The blood-brain barrier: Bottleneck in brain drug development. NeuroRx 2005, 2, 3–14. [Google Scholar] [CrossRef]
- Mittal, D.; Ali, A.; Md, S.; Baboota, S.; Sahni, J.K.; Ali, J. Insights into direct nose to brain delivery: Current status and future perspective. Drug Deliv. 2014, 21, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Ermisch, A.; Barth, T.; Rühle, H.J.; Skopkova, J.; Hrbas, P.; Landgraf, R. On the blood-brain barrier to peptides: Accumulation of labelled vasopressin, DesGlyNH2-vasopressin and oxytocin by brain regions. Endocrinol. Exp. 1985, 19, 29–37. [Google Scholar] [PubMed]
- Mens, W.B.; Witter, A.; Greidanus, T.B.V.W. Penetration of neurohypophyseal hormones from plasma into cerebrospinal fluid (CSF): Half-times of disappearance of these neuropeptides from CSF. Brain Res. 1983, 262, 143–149. [Google Scholar] [CrossRef]
- Bowen, M.T. Does peripherally administered oxytocin enter the brain? Compelling new evidence in a long-running debate. Pharmacol. Res. 2019, 146, 104325. [Google Scholar] [CrossRef]
- Leng, G.; Ludwig, M. Intranasal oxytocin: Myths and delusions. Biol. Psychiatry 2016, 79, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Quintana, D.S.; Lischke, A.; Grace, S.; Scheele, D.; Ma, Y.; Becker, B. Advances in the field of intranasal oxytocin research: Lessons learned and future directions for clinical research. Mol. Psychiatry 2021, 26, 80–91. [Google Scholar] [CrossRef]
- Lee, M.R.; Scheidweiler, K.B.; Diao, X.X.; Akhlaghi, F.; Cummins, A.; Huestis, M.A.; Leggio, L.; Averbeck, B.B. Oxytocin by intranasal and intravenous routes reaches the cerebrospinal fluid in rhesus macaques: Determination using a novel oxytocin assay. Mol. Psychiatry 2018, 23, 115–122. [Google Scholar] [CrossRef]
- Kou, J.; Lan, C.; Zhang, Y.; Wang, Q.; Zhou, F.; Zhao, Z.; Montag, C.; Yao, S.; Becker, B.; Kendrick, K.M. In the nose or on the tongue? Contrasting motivational effects of oral and intranasal oxytocin on arousal and reward during social processing. Transl. Psychiatry 2021, 11, 94. [Google Scholar] [CrossRef]
- Zhuang, Q.; Zheng, X.; Yao, S.; Zhao, W.; Becker, B.; Xu, X.; Kendrick, K.M. Oral, similar to intranasal, administration of oxytocin decreases top-down social attention, but additionally enhances bottom-up control. medRxiv 2021. [Google Scholar] [CrossRef]
- Bharadwaj, V.N.; Tzabazis, A.Z.; Klukinov, M.; Manering, N.A.; Yeomans, D.C. Intranasal Administration for Pain: Oxytocin and Other Polypeptides. Pharmaceutics 2021, 13, 1088. [Google Scholar] [CrossRef]
- Zheng, X.; Kendrick, K.M. Neural and molecular contributions to pathological jealousy and a potential therapeutic role for intranasal oxytocin. Front. Pharmacol. 2021, 12, 652473. [Google Scholar] [CrossRef]
- Modi, M.E.; Connor-Stroud, F.; Landgraf, R.; Young, L.J.; Parr, L.A. Aerosolized oxytocin increases cerebrospinal fluid oxytocin in rhesus macaques. Psychoneuroendocrinology 2014, 45, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Born, J.; Lange, T.; Kern, W.; McGregor, G.P.; Bickel, U.; Fehm, H.L. Sniffing neuropeptides: A transnasal approach to the human brain. Nat. Neurosci. 2002, 5, 514–516. [Google Scholar] [CrossRef]
- Striepens, N.; Kendrick, K.M.; Hanking, V.; Landgraf, R.; Wüllner, U.; Maier, W.; Hurlemann, R. Elevated cerebrospinal fluid and blood concentrations of oxytocin following its intranasal administration in humans. Sci. Rep. 2013, 3, 3440. [Google Scholar] [CrossRef]
- Beard, R.; Singh, N.; Grundschober, C.; Gee, A.D.; Tate, E.W. High-yielding 18 F radiosynthesis of a novel oxytocin receptor tracer, a probe for nose-to-brain oxytocin uptake in vivo. Chem. Commun. 2018, 54, 8120–8123. [Google Scholar] [CrossRef] [Green Version]
- Neumann, I.D.; Maloumby, R.; Beiderbeck, D.I.; Lukas, M.; Landgraf, R. Increased brain and plasma oxytocin after nasal and peripheral administration in rats and mice. Psychoneuroendocrinology 2013, 38, 1985–1993. [Google Scholar] [CrossRef]
- Smith, A.S.; Korgan, A.C.; Young, W.S. Oxytocin delivered nasally or intraperitoneally reaches the brain and plasma of normal and oxytocin knockout mice. Pharmacol. Res. 2019, 146, 104324. [Google Scholar] [CrossRef]
- Galbusera, A.; De Felice, A.; Girardi, S.; Bassetto, G.; Maschietto, M.; Nishimori, K.; Chini, B.; Papaleo, F.; Vassanelli, S.; Gozzi, A. Intranasal oxytocin and vasopressin modulate divergent brainwide functional substrates. Neuropsychopharmacology 2017, 42, 1420–1434. [Google Scholar] [CrossRef]
- Lee, M.R.; Shnitko, T.A.; Blue, S.W.; Kaucher, A.V.; Winchell, A.J.; Erikson, D.W.; Grant, K.; Leggio, L. Labeled oxytocin administered via the intranasal route reaches the brain in rhesus macaques. Nat. Commun. 2020, 11, 2783. [Google Scholar] [CrossRef]
- Martins, D.A.; Mazibuko, N.; Zelaya, F.; Vasilakopoulou, S.; Loveridge, J.; Oates, A.; Maltezos, S.; Mehta, M.; Wastling, S.; Howard, M.; et al. Effects of route of administration on oxytocin-induced changes in regional cerebral blood flow in humans. Nat. Commun. 2020, 11, 1160. [Google Scholar] [CrossRef] [Green Version]
- Paloyelis, Y.; Doyle, O.M.; Zelaya, F.O.; Maltezos, S.; Williams, S.C.; Fotopoulou, A.; Howard, M.A. A spatiotemporal profile of in vivo cerebral blood flow changes following intranasal oxytocin in humans. Biol. Psychiatry 2016, 79, 693–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habets, P.C.; Mclain, C.; Meijer, O.C. Brain areas affected by intranasal oxytocin show higher oxytocin receptor expression. Eur. J. Neurosci. 2021, 54, 6374–6381. [Google Scholar] [CrossRef] [PubMed]
- Quintana, D.S.; Westlye, L.T.; Rustan, Ø.G.; Tesli, N.; Poppy, C.L.; Smevik, H.; Tesli, M.; Røine, M.; Mahmoud, R.A.; Smerud, K.T.; et al. Low-dose oxytocin delivered intranasally with Breath Powered device affects social-cognitive behavior: A randomized four-way crossover trial with nasal cavity dimension assessment. Transl. Psychiatry 2015, 5, e602. [Google Scholar] [CrossRef] [PubMed]
- Pietrowsky, R.; Strüben, C.; Mölle, M.; Fehm, H.L.; Born, J. Brain potential changes after intranasal vs. intravenous administration of vasopressin: Evidence for a direct nose-brain pathway for peptide effects in humans. Biol. Psychiatry 1996, 39, 332–340. [Google Scholar] [CrossRef]
- Quintana, D.S.; Westlye, L.T.; Smerud, K.T.; Mahmoud, R.A.; Andreassen, O.A.; Djupesland, P.G. Saliva oxytocin measures do not reflect peripheral plasma concentrations after intranasal oxytocin administration in men. Horm. Behav. 2018, 102, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Daughters, K.; Manstead, A.S.; Hubble, K.; Rees, A.; Thapar, A.; van Goozen, S.H. Salivary oxytocin concentrations in males following intranasal administration of oxytocin: A double-blind, cross-over study. PLoS ONE 2015, 10, e0145104. [Google Scholar] [CrossRef] [Green Version]
- Van IJzendoorn, M.H.; Bhandari, R.; Van der Veen, R.; Grewen, K.; Bakermans-Kranenburg, M.J. Elevated salivary levels of oxytocin persist more than 7 h after intranasal administration. Front. Neurosci. 2012, 6, 174. [Google Scholar] [CrossRef] [Green Version]
- Maejima, Y.; Horita, S.; Otsuka, A.; Hidema, S.; Nishimori, K.; Shimomura, K. Oral oxytocin delivery with proton pump inhibitor pretreatment decreases food intake. Peptides 2020, 128, 170312. [Google Scholar] [CrossRef]
- De Groot, A.N.; Vree, T.B.; Hekster, Y.A.; Pesman, G.J.; Sweep, F.C.; Van Dongen, P.J.; Van Roosmalen, J. Bioavailability and pharmacokinetics of sublingual oxytocin in male volunteers. J. Pharm. Pharmacol. 1995, 47, 571–575. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Higashida, H. RAGE regulates oxytocin transport into the brain. Commun. Biol. 2020, 3, 70. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Liang, M.; Munesue, S.; Deguchi, K.; Harashima, A.; Furuhara, K.; Yuhi, T.; Zhong, J.; Akther, S.; Goto, H.; et al. Vascular RAGE transports oxytocin into the brain to elicit its maternal bonding behaviour in mice. Commun. Biol. 2019, 2, 76. [Google Scholar] [CrossRef]
- Ring, R.H.; Malberg, J.E.; Potestio, L.; Ping, J.; Boikess, S.; Luo, B.; Schechter, L.E.; Rizzo, S.; Rahman, Z.; Rosenzweig-Lipson, S. Anxiolytic-like activity of oxytocin in male mice: Behavioral and autonomic evidence, therapeutic implications. Psychopharmacology 2006, 185, 218–225. [Google Scholar] [CrossRef]
- Sakamoto, T.; Sugimoto, S.; Uekita, T. Effects of intraperitoneal and intracerebroventricular injections of oxytocin on social and emotional behaviors in pubertal male mice. Physiol. Behav. 2019, 212, 112701. [Google Scholar] [CrossRef]
- Hollander, E.; Bartz, J.; Chaplin, W.; Phillips, A.; Sumner, J.; Soorya, L.; Anagnostou, E.; Wasserman, S. Oxytocin increases retention of social cognition in autism. Biol. Psychiatry 2007, 61, 498–503. [Google Scholar] [CrossRef]
- Hollander, E.; Novotny, S.; Hanratty, M.; Yaffe, R.; DeCaria, C.M.; Aronowitz, B.R.; Mosovich, S. Oxytocin infusion reduces repetitive behaviors in adults with autistic and Asperger’s disorders. Neuropsychopharmacology 2003, 28, 193–198. [Google Scholar] [CrossRef]
- Ferris, C.F.; Yee, J.R.; Kenkel, W.M.; Dumais, K.M.; Moore, K.; Veenema, A.H.; Kulkarni, P.; Perkybile, A.M.; Carter, C.S. Distinct BOLD activation profiles following central and peripheral oxytocin administration in awake rats. Front. Behav. Neurosci. 2015, 9, 245. [Google Scholar] [CrossRef]
- Quintana, D.S.; Westlye, L.T.; Alnæs, D.; Rustan, Ø.G.; Kaufmann, T.; Smerud, K.T.; Mahmoud, R.A.; Djupesland, P.G.; Andreassen, O.A. Low dose intranasal oxytocin delivered with Breath Powered device dampens amygdala response to emotional stimuli: A peripheral effect-controlled within-subjects randomized dose-response fMRI trial. Psychoneuroendocrinology 2016, 69, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Kou, J.; Zhang, Y.; Zhou, F.; Sindermann, C.; Montag, C.; Becker, B.; Kendrick, K.M. A randomized trial shows dose-frequency and genotype may determine the therapeutic efficacy of intranasal oxytocin. Psychol. Med. 2020, 1–10. [Google Scholar] [CrossRef]
- Caldwell, H.K.; Lee, H.J.; Macbeth, A.H.; Young, W.S. 3rd Vasopressin: Behavioral roles of an “original” neuropeptide. Prog. Neurobiol. 2008, 84, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Carter, C.S. Oxytocin pathways and the evolution of human behavior. Annu. Rev. Psychol. 2014, 65, 17–39. [Google Scholar] [CrossRef] [Green Version]
- Liard, J.F. Vasopressin in cardiovascular control: Role of circulating vasopressin. Clin. Sci. 1984, 67, 473–481. [Google Scholar] [CrossRef]
- Share, L. Role of vasopressin in cardiovascular regulation. Physiol. Rev. 1988, 68, 1248–1284. [Google Scholar] [CrossRef] [PubMed]
- Hupf, H.; Grimm, D.; Riegger, G.A.; Schunkert, H. Evidence for a vasopressin system in the rat heart. Circ. Res. 1999, 84, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Monstein, H.J.; Truedsson, M.; Ryberg, A.; Ohlsson, B. Vasopressin receptor mRNA expression in the human gastrointestinal tract. Eur. Surg. Res. 2008, 40, 34–40. [Google Scholar] [CrossRef]
- Chae, J.H.; Nahas, Z.; Lomarev, M.; Denslow, S.; Lorberbaum, J.P.; Bohning, D.E.; George, M.S. A review of functional neuroimaging studies of vagus nerve stimulation (VNS). J. Psychiatr. Res. 2003, 37, 443–455. [Google Scholar] [CrossRef]
- Zhu, S.; Zhang, X.; Qing, Y.; Zhang, Y.; Yao, S.; Kendrick, K.; Zhao, W. Transcutaneous auricular vagus nerve stimulation increases eye-gaze to salient facial features and oxytocin release. Brain Stimul. Basic Transl. Clin. Res. Neuromodul. 2021, 14, 1639. [Google Scholar] [CrossRef]
- Everett, N.A.; Turner, A.J.; Costa, P.A.; Baracz, S.J.; Cornish, J.L. The vagus nerve mediates the suppressing effects of peripherally administered oxytocin on methamphetamine self-administration and seeking in rats. Neuropsychopharmacology 2021, 46, 297–304. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Maejima, Y.; Suyama, S.; Yoshida, M.; Arai, T.; Katsurada, K.; Kumari, P.; Nakabayashi, H.; Kakei, M.; Yada, T. Peripheral oxytocin activates vagal afferent neurons to suppress feeding in normal and leptin-resistant mice: A route for ameliorating hyperphagia and obesity. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2015, 308, R360–R369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamer, M.; Büchel, C. Oxytocin specifically enhances valence-dependent parasympathetic responses. Psychoneuroendocrinology 2012, 37, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Kemp, A.H.; Quintana, D.S.; Kuhnert, R.L.; Griffiths, K.; Hickie, I.B.; Guastella, A.J. Oxytocin increases heart rate variability in humans at rest: Implications for social approach-related motivation and capacity for social engagement. PLoS ONE 2012, 7, e44014. [Google Scholar]
- Martins, D.; Davies, C.; De Micheli, A.; Oliver, D.; Krawczun-Rygmaczewska, A.; Fusar-Poli, P.; Paloyelis, Y. Intranasal oxytocin increases heart-rate variability in men at clinical high risk for psychosis: A proof-of-concept study. Transl. Psychiatry 2020, 10, 227. [Google Scholar] [CrossRef]
- Norman, G.J.; Cacioppo, J.T.; Morris, J.S.; Malarkey, W.B.; Berntson, G.G.; DeVries, A.C. Oxytocin increases autonomic cardiac control: Moderation by loneliness. Biol. Psychol. 2011, 86, 174–180. [Google Scholar] [CrossRef]
- Schoormans, D.; Kop, W.J.; Kunst, L.E.; Riem, M.M.E. Oxytocin effects on resting-state heart rate variability in women: The role of childhood rearing experiences. Compr. Psychoneuroendocrinol. 2020, 3, 100007. [Google Scholar] [CrossRef]
- Tanaka, A.; Furubayashi, T.; Arai, M.; Inoue, D.; Kimura, S.; Kiriyama, A.; Kusamori, K.; Katsumi, H.; Yutani, R.; Sakane, T.; et al. Delivery of oxytocin to the brain for the treatment of autism spectrum disorder by nasal application. Mol. Pharm. 2018, 15, 1105–1111. [Google Scholar] [CrossRef]
- Quintana, D.S.; Westlye, L.T.; Alnæs, D.; Kaufmann, T.; Mahmoud, R.A.; Smerud, K.T.; Djupesland, P.G.; Andreassen, O.A. Low-dose intranasal oxytocin delivered with breath powered device modulates pupil diameter and amygdala activity: A randomized controlled pupillometry and fMRI study. Neuropsychopharmacology 2019, 44, 306–313. [Google Scholar] [CrossRef] [Green Version]
- Jurek, B.; Neumann, I.D. The oxytocin receptor: From intracellular signaling to behavior. Physiol. Rev. 2018, 98, 1805–1908. [Google Scholar] [CrossRef]
- Busnelli, M.; Chini, B. Molecular basis of oxytocin receptor signalling in the brain: What we know and what we need to know. In Behavioral Pharmacology of Neuropeptides: Oxytocin; Springer: Cham, Switzerland, 2017; pp. 3–29. [Google Scholar] [CrossRef]
- Spengler, F.B.; Schultz, J.; Scheele, D.; Essel, M.; Maier, W.; Heinrichs, M.; Hurlemann, R. Kinetics and dose dependency of intranasal oxytocin effects on amygdala reactivity. Biol. Psychiatry 2017, 82, 885–894. [Google Scholar] [CrossRef]
- Martins, D.; Brodmann, K.; Veronese, M.; Dipasquale, O.; Mazibuko, N.; Schuschnig, U.; Zelaya, F.; Fotopoulou, A.; Paloyelis, Y. Less is more: A dose-response mechanistic account of intranasal oxytocin pharmacodynamics in the human brain. bioRxiv 2021. [Google Scholar] [CrossRef]
- Price, D.; Burris, D.; Cloutier, A.; Thompson, C.B.; Rilling, J.K.; Thompson, R.R. Dose-dependent and lasting influences of intranasal vasopressin on face processing in men. Front. Endocrinol. 2017, 8, 220. [Google Scholar] [CrossRef]
- Borland, J.M.; Rilling, J.K.; Frantz, K.J.; Albers, H.E. Sex-dependent regulation of social reward by oxytocin: An inverted U hypothesis. Neuropsychopharmacology 2019, 44, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Becker, B.; Luo, L.; Geng, Y.; Zhao, W.; Yin, Y.; Hu, J.; Gao, Z.; Gong, Q.; Hurlemann, R.; et al. Oxytocin, the peptide that bonds the sexes also divides them. Proc. Natl. Acad. Sci. USA 2016, 113, 7650–7654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieberz, J.; Scheele, D.; Spengler, F.B.; Matheisen, T.; Schneider, L.; Stoffel-Wagner, B.; Hurlemann, R. Kinetics of oxytocin effects on amygdala and striatal reactivity vary between women and men. Neuropsychopharmacology 2020, 45, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Becker, B.; Geng, Y.; Zhao, Z.; Gao, S.; Zhao, W.; Yao, S.; Zheng, X.; Ma, X.; Gao, Z.; et al. Sex-dependent neural effect of oxytocin during subliminal processing of negative emotion faces. Neuroimage 2017, 162, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, S.; Shenoy, S.K. GPCR desensitization: Acute and prolonged phases. Cell. Signal. 2018, 41, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Michetti, C.; Busnelli, M.; Manago, F.; Sannino, S.; Scheggia, D.; Giancardo, L.; Sona, D.; Murino, V.; Chini, B.; et al. Chronic and acute intranasal oxytocin produce divergent social effects in mice. Neuropsychopharmacology 2014, 39, 1102–1114. [Google Scholar] [CrossRef] [Green Version]
- Freeman, S.M.; Ngo, J.; Singh, B.; Masnaghetti, M.; Bales, K.L.; Blevins, J.E. Effects of chronic oxytocin administration and diet composition on oxytocin and vasopressin 1a receptor binding in the rat brain. Neuroscience 2018, 392, 241–251. [Google Scholar] [CrossRef]
- Bales, K.L.; Perkeybile, A.M.; Conley, O.G.; Lee, M.H.; Guoynes, C.D.; Downing, G.M.; Yun, C.R.; Solomon, M.; Jacob, S.; Mendoza, S.P. Chronic intranasal oxytocin causes long-term impairments in partner preference formation in male prairie voles. Biol. Psychiatry 2013, 74, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Winter, J.; Meyer, M.; Berger, I.; Royer, M.; Bianchi, M.; Kuffner, K.; Peters, S.; Stang, S.; Langgartner, D.; Hartmann, F.; et al. Chronic oxytocin-driven alternative splicing of CRFR2α induces anxiety. Mol. Psychiatry 2021, 1–14. [Google Scholar] [CrossRef]
- Kou, J.; Zhang, Y.; Zhou, F.; Gao, Z.; Yao, S.; Zhao, W.; Li, H.; Lei, Y.; Gao, S.; Kendrick, K.M.; et al. Anxiolytic effects of chronic intranasal oxytocin on neural responses to threat are dose-frequency dependent. bioRxiv 2021. [Google Scholar] [CrossRef]
- Dal Monte, O.; Noble, P.L.; Turchi, J.; Cummins, A.; Averbeck, B.B. CSF and blood oxytocin concentration changes following intranasal delivery in macaque. PLoS ONE 2014, 9, e103677. [Google Scholar] [CrossRef] [Green Version]
- Guastella, A.J.; Hickie, I.B.; McGuinness, M.M.; Otis, M.; Woods, E.A.; Disinger, H.M.; Chan, H.K.; Chen, T.F.; Banati, R.B. Recommendations for the standardisation of oxytocin nasal administration and guidelines for its reporting in human research. Psychoneuroendocrinology 2013, 38, 612–625. [Google Scholar] [CrossRef]
- Quintana, D.S.; Westlye, L.T.; Hope, S.; Naerland, T.; Elvsåshagen, T.; Dørum, E.; Rustan, Ø.; Valstad, M.; Rezvaya, L.; Lishaugen, H.; et al. Dose-dependent social-cognitive effects of intranasal oxytocin delivered with novel Breath Powered device in adults with autism spectrum disorder: A randomized placebo-controlled double-blind crossover trial. Transl. Psychiatry 2017, 7, e1136. [Google Scholar] [CrossRef] [Green Version]
- Djupesland, P.G. Nasal drug delivery devices: Characteristics and performance in a clinical perspective—A review. Drug Deliv. Transl. Res. 2013, 3, 42–62. [Google Scholar] [CrossRef] [Green Version]
- Djupesland, P.G.; Skretting, A.; Winderen, M.; Holand, T. Breath actuated device improves delivery to target sites beyond the nasal valve. Laryngoscope 2006, 116, 466–472. [Google Scholar] [CrossRef]
- Moffa, A.; Costantino, A.; Rinaldi, V.; Sabatino, L.; Trecca EM, C.; Baptista, P.; Campisi, P.; Cassano, M.; Casale, M. Nasal delivery devices: A comparative study on cadaver model. BioMed. Res. Int. 2019, 2019, 4602651. [Google Scholar] [CrossRef] [Green Version]
- Farzal, Z.; Basu, S.; Burke, A.; Fasanmade, O.O.; Lopez, E.M.; Bennett, W.D.; Ebert, C.S., Jr.; Zanation, A.M.; Senior, B.A.; Kimbell, J.S. Comparative study of simulated nebulized and spray particle deposition in chronic rhinosinusitis patients. Int. Forum Allergy Rhinol. 2019, 9, 746–758. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, S.; Kendrick, K.M. Effects of Intranasal Administration of Oxytocin and Vasopressin on Social Cognition and Potential Routes and Mechanisms of Action. Pharmaceutics 2022, 14, 323. https://doi.org/10.3390/pharmaceutics14020323
Yao S, Kendrick KM. Effects of Intranasal Administration of Oxytocin and Vasopressin on Social Cognition and Potential Routes and Mechanisms of Action. Pharmaceutics. 2022; 14(2):323. https://doi.org/10.3390/pharmaceutics14020323
Chicago/Turabian StyleYao, Shuxia, and Keith Maurice Kendrick. 2022. "Effects of Intranasal Administration of Oxytocin and Vasopressin on Social Cognition and Potential Routes and Mechanisms of Action" Pharmaceutics 14, no. 2: 323. https://doi.org/10.3390/pharmaceutics14020323
APA StyleYao, S., & Kendrick, K. M. (2022). Effects of Intranasal Administration of Oxytocin and Vasopressin on Social Cognition and Potential Routes and Mechanisms of Action. Pharmaceutics, 14(2), 323. https://doi.org/10.3390/pharmaceutics14020323