


Mitigating Cardiotoxicity of Dendrimers: Angiotensin-(1-7) via Its Mas Receptor Ameliorates PAMAM-Induced Cardiac Dysfunction in the Isolated Mammalian Heart

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Procedures
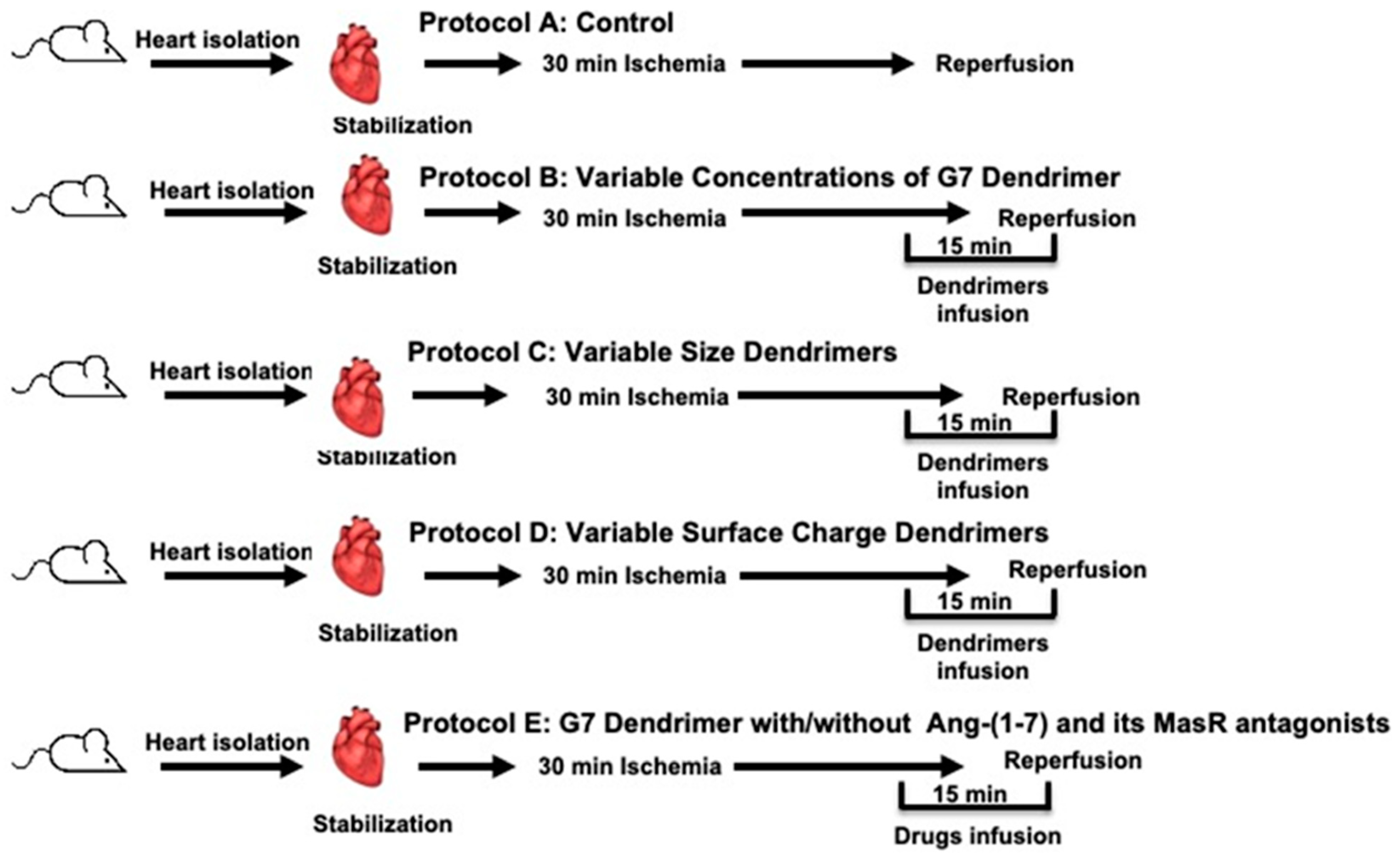
2.3. Experimental Study Protocols
2.4. Assessment of Heart Function
2.5. Assessment of Cardiac Damage through Infarct Size Measurement and Determination of Cardiac Enzyme Levels
2.6. Data Analysis
3. Results
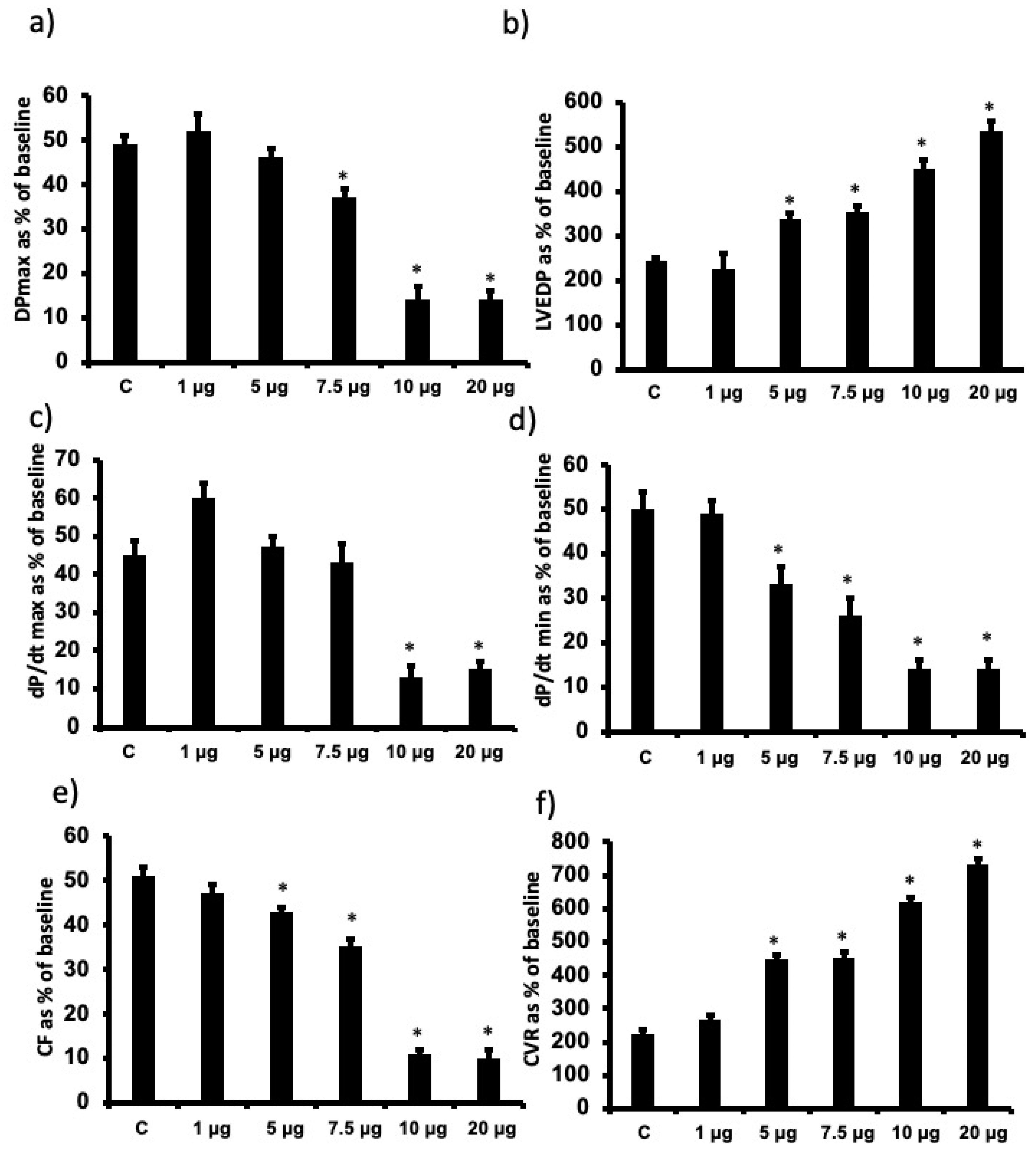
3.1. The Effects of Increasing Doses of Cationic G7 PAMAM Dendrimers on Cardiac Function Recovery following I/R Injury in Isolated Rat Hearts
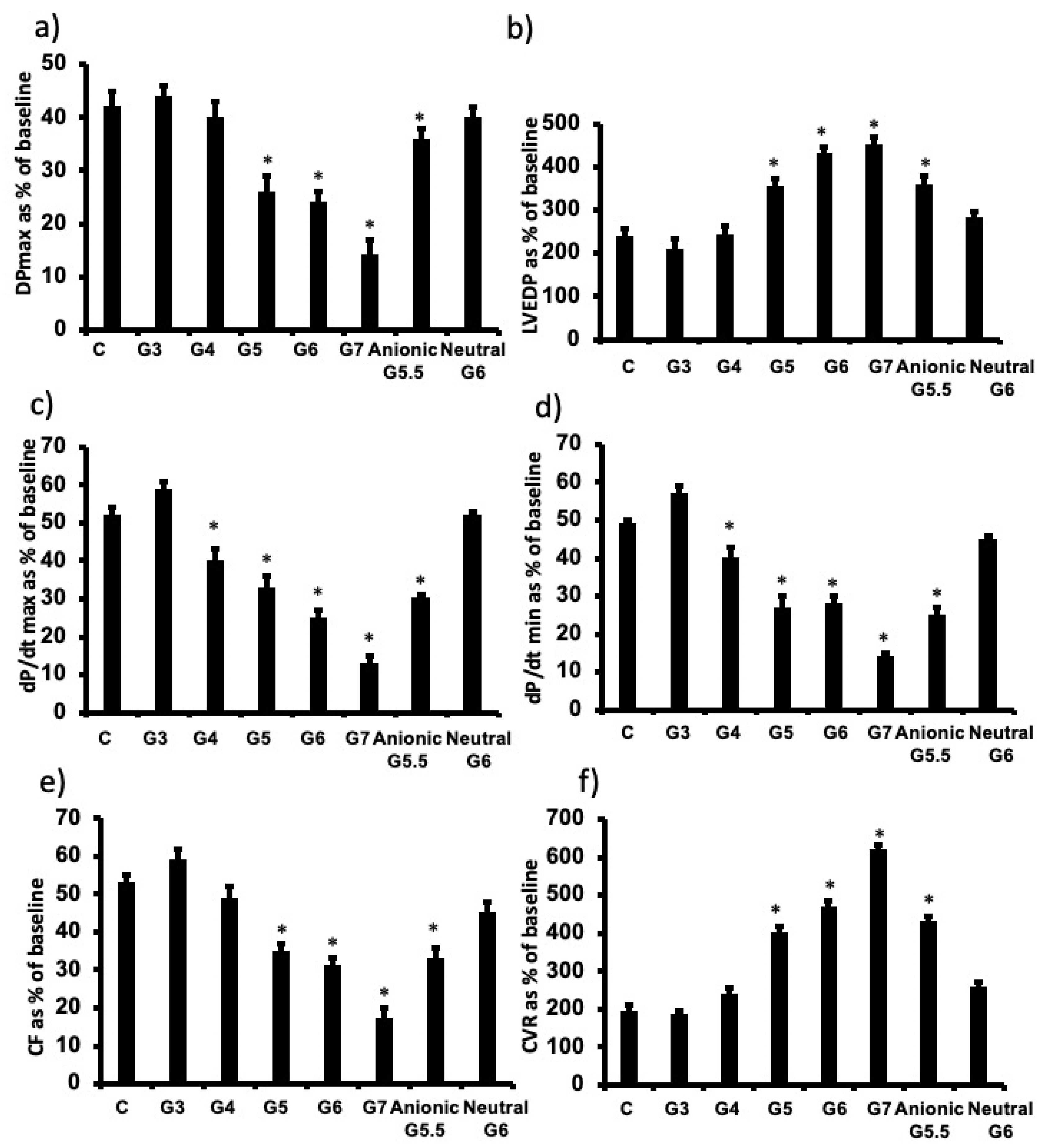
3.2. The Influence of Cationic Dendrimer Generation on Cardiac Function Recovery following I/R Injury in Isolated Rat Hearts
3.3. The influence of PAMAM Dendrimer Surface Chemistry on Cardiac Function Recovery after I/R Injury in Isolated Rat Hearts
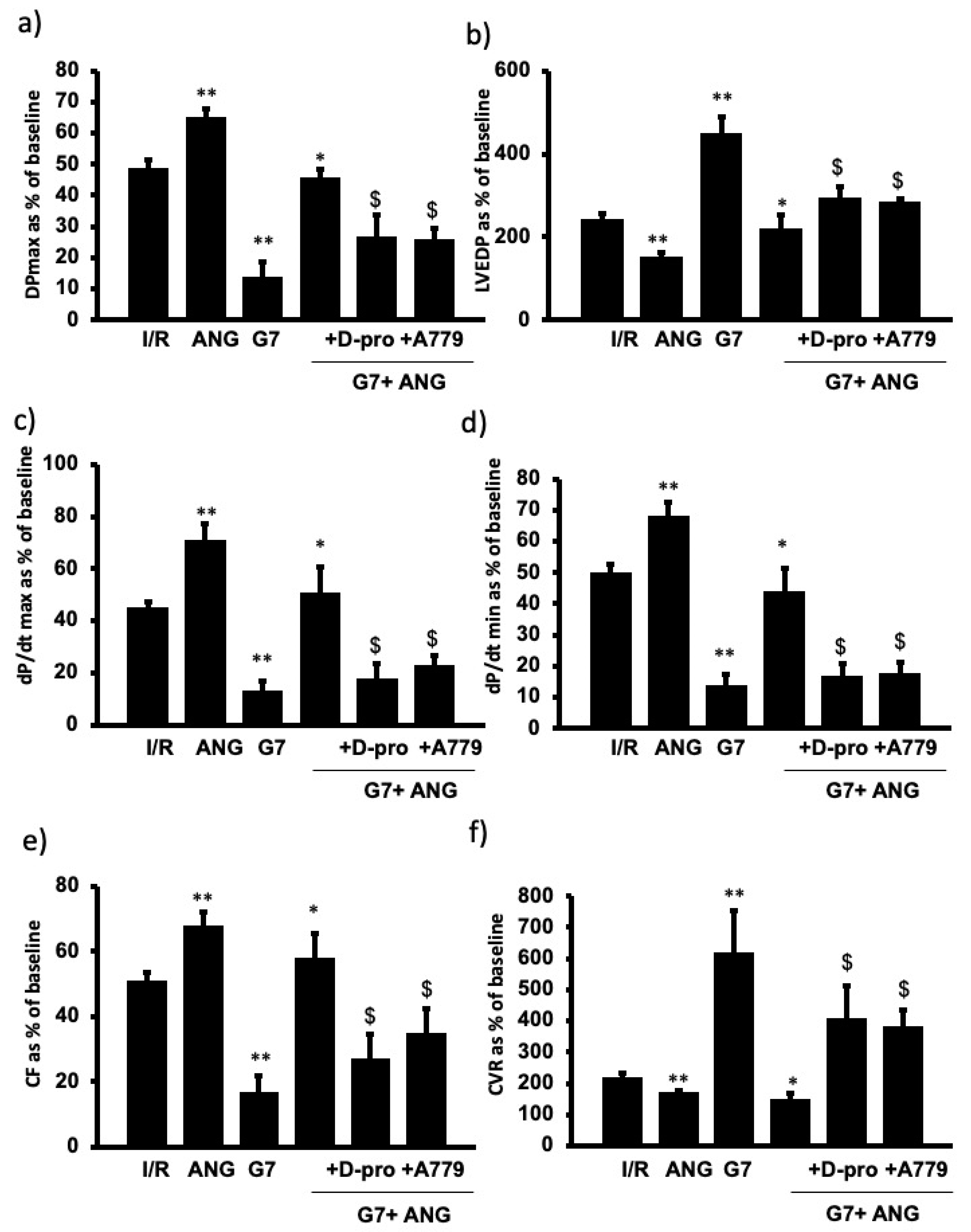
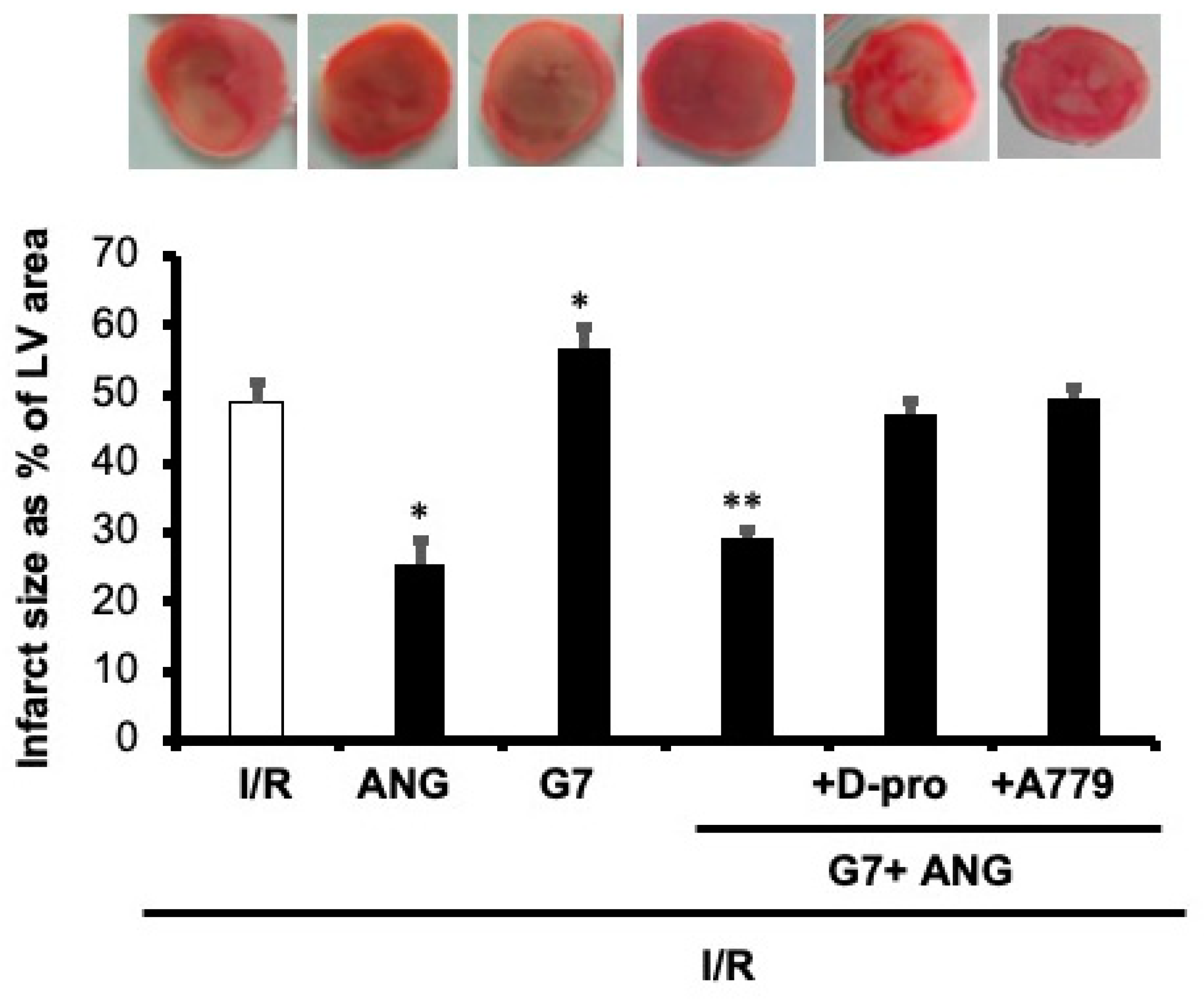
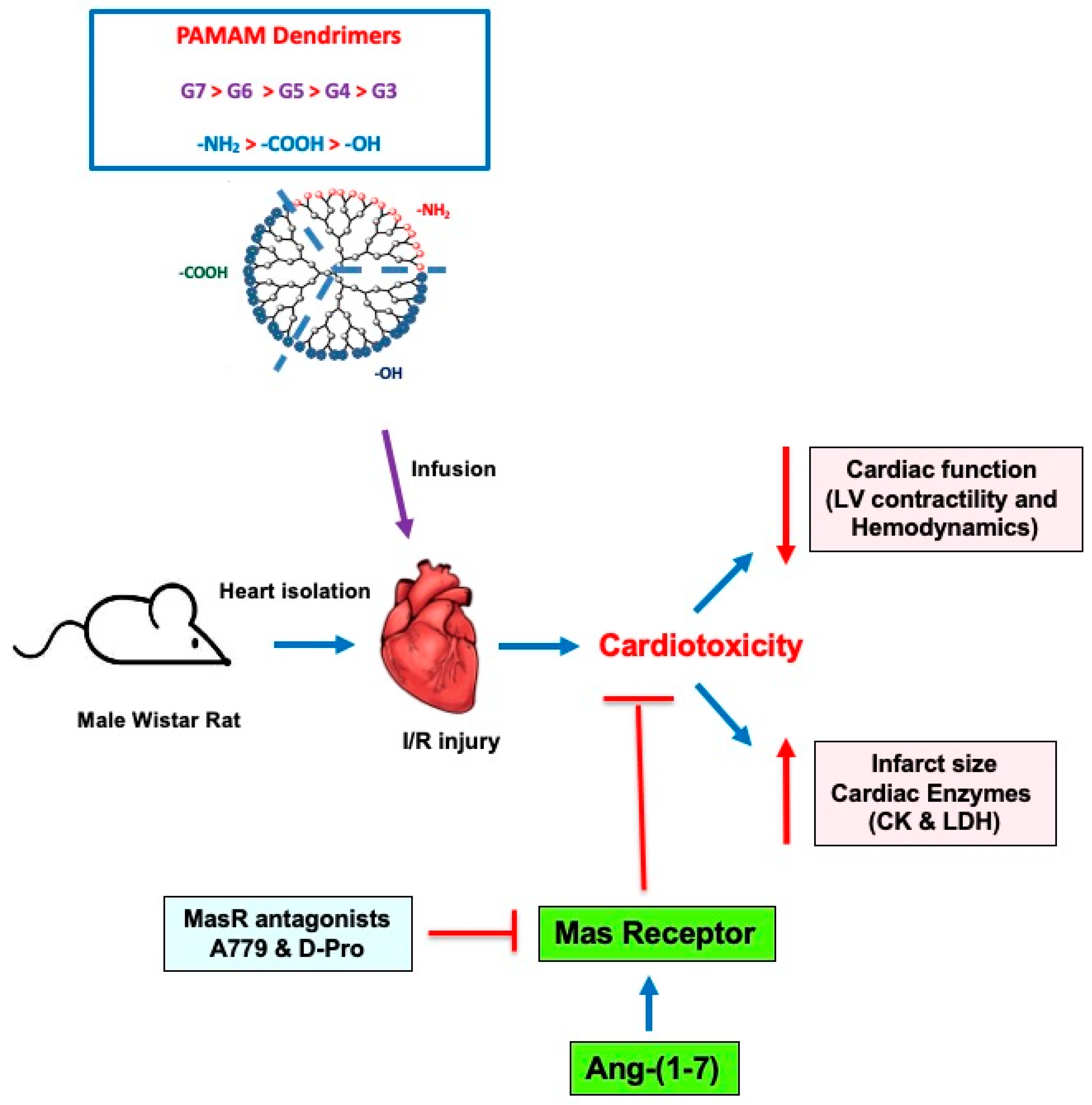
3.4. PAMAM-Induced Cardiac Dysfunction Can Be Rescued by Ang-(1-7) in a Mas Receptor-Dependent Mechanism of Action
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abedi-Gaballu, F.; Dehghan, G.; Ghaffari, M.; Yekta, R.; Abbaspour-Ravasjani, S.; Baradaran, B.; Ezzati Nazhad Dolatabadi, J.; Hamblin, M.R. PAMAM dendrimers as efficient drug and gene delivery nanosystems for cancer therapy. Appl. Mater. Today 2018, 12, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Kannan, R.M.; Nance, E.; Kannan, S.; Tomalia, D.A. Emerging concepts in dendrimer-based nanomedicine: From design principles to clinical applications. J. Intern. Med. 2014, 276, 579–617. [Google Scholar] [CrossRef] [PubMed]
- Kheraldine, H.; Rachid, O.; Habib, A.M.; Al Moustafa, A.-E.; Benter, I.F.; Akhtar, S. Emerging innate biological properties of nano-drug delivery systems: A focus on PAMAM dendrimers and their clinical potential. Adv. Drug Deliv. Rev. 2021, 178, 113908. [Google Scholar] [CrossRef]
- Li, J.; Liang, H.; Liu, J.; Wang, Z. Poly (amidoamine) (PAMAM) dendrimer mediated delivery of drug and pDNA/siRNA for cancer therapy. Int. J. Pharm. 2018, 546, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Tomalia, D.; Reyna, L.; Svenson, S. Dendrimers as multi-purpose nanodevices for oncology drug delivery and diagnostic imaging. Biochem. Soc. Trans. 2007, 35, 61–67. [Google Scholar] [CrossRef]
- Kheraldine, H.; Gupta, I.; Alhussain, H.; Jabeen, A.; Cyprian, F.S.; Akhtar, S.; Al Moustafa, A.-E.; Rachid, O. Substantial cell apoptosis provoked by naked PAMAM dendrimers in HER2-positive human breast cancer via JNK and ERK1/ERK2 signalling pathways. Comput. Struct. Biotechnol. J. 2021, 19, 2881–2890. [Google Scholar] [CrossRef]
- Akhtar, S.; Al-Zaid, B.; El-Hashim, A.Z.; Chandrasekhar, B.; Attur, S.; Benter, I.F. Impact of PAMAM delivery systems on signal transduction pathways in vivo: Modulation of ERK1/2 and p38 MAP kinase signaling in the normal and diabetic kidney. Int. J. Pharm. 2016, 514, 353–363. [Google Scholar] [CrossRef]
- Akhtar, S.; Chandrasekhar, B.; Yousif, M.H.; Renno, W.; Benter, I.F.; El-Hashim, A.Z. Chronic administration of nano-sized PAMAM dendrimers in vivo inhibits EGFR-ERK1/2-ROCK signaling pathway and attenuates diabetes-induced vascular remodeling and dysfunction. Nanomed. Nanotechnol. Biol. Med. 2019, 18, 78–89. [Google Scholar] [CrossRef]
- Akhtar, S.; El-Hashim, A.Z.; Chandrasekhar, B.; Attur, S.; Benter, I.F. Naked Polyamidoamine Polymers Intrinsically Inhibit Angiotensin II-Mediated EGFR and ErbB2 Transactivation in a Dendrimer Generation- and Surface Chemistry-Dependent Manner. Mol. Pharm. 2016, 13, 1575–1586. [Google Scholar] [CrossRef]
- Chanyshev, B.; Shainberg, A.; Isak, A.; Litinsky, A.; Chepurko, Y.; Tosh, D.K.; Phan, K.; Gao, Z.-G.; Hochhauser, E.; Jacobson, K.A. Anti-ischemic effects of multivalent dendrimeric A3 adenosine receptor agonists in cultured cardiomyocytes and in the isolated rat heart. Pharmacol. Res. 2012, 65, 338–346. [Google Scholar] [CrossRef][Green Version]
- Johnson, T.A.; Stasko, N.A.; Matthews, J.L.; Cascio, W.E.; Holmuhamedov, E.; Johnson, C.B.; Schoenfisch, M.H. Reduced ischemia/reperfusion injury via glutathione-initiated nitric oxide-releasing dendrimers. Nitric Oxide 2010, 22, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Sayed, N.; Tambe, P.; Kumar, P.; Jadhav, S.; Paknikar, K.M.; Gajbhiye, V. miRNA transfection via poly(amidoamine)-based delivery vector prevents hypoxia/reperfusion-induced cardiomyocyte apoptosis. Nanomedicine 2020, 15, 163–181. [Google Scholar] [CrossRef] [PubMed]
- Wan, T.C.; Tosh, D.K.; Du, L.; Gizewski, E.T.; Jacobson, A.K.; Auchampach, A.J. Polyamidoamine (PAMAM) dendrimer conjugate specifically activates the A3 adenosine receptor to improve post-ischemic/reperfusion function in isolated mouse hearts. BMC Pharmacol. 2011, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Babiker, F.; Benter, I.F.; Akhtar, S. Nanotoxicology of Dendrimers in the Mammalian Heart: Ex vivo and in vivo Administration of G6 PAMAM Nanoparticles Impairs Recovery of Cardiac Function Following Ischemia-Reperfusion Injury. Int. J. Nanomed. 2020, 15, 4393–4405. [Google Scholar] [CrossRef]
- Hausenloy, D.J.; Baxter, G.; Bell, R.; Bøtker, H.E.; Davidson, S.M.; Downey, J.; Heusch, G.; Kitakaze, M.; Lecour, S.; Mentzer, R.; et al. Translating novel strategies for cardioprotection: The Hatter Workshop Recommendations. Basic Res. Cardiol. 2010, 105, 677–686. [Google Scholar] [CrossRef]
- Wereski, R.; Kimenai, D.M.; Bularga, A.; Taggart, C.; Lowe, D.J.; Mills, N.L.; Chapman, A.R. Risk factors for type 1 and type 2 myocardial infarction. Eur. Heart J. 2021, 43, 127–135. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, L.; Wang, S.; Cheng, H.; Xu, L.; Pei, G.; Wang, Y.; Fu, C.; Jiang, Y.; He, C.; et al. Signaling pathways and targeted therapy for myocardial infarction. Signal Transduct. Target. Ther. 2022, 7, 78. [Google Scholar] [CrossRef]
- Schäfer, A.; König, T.; Bauersachs, J.; Akin, M. Novel Therapeutic Strategies to Reduce Reperfusion Injury After Acute Myocardial Infarction. Curr. Probl. Cardiol. 2022, 47, 101398. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Y.; Li, G. Preclinical multi-target strategies for myocardial ischemia-reperfusion injury. Front. Cardiovasc. Med. 2022, 9. [Google Scholar] [CrossRef]
- Babiker, A.F.; Elkhalifa, A.L.; Moukhyer, E.M. Awareness of hypertension and factors associated with uncontrolled hypertension in Sudanese adults: Cardiovascular topic. Cardiovasc. J. Afr. 2013, 24, 208–212. [Google Scholar] [CrossRef]
- Ribichini, F. ACUTE MYOCARDIAL INFARCTION: REPERFUSION TREATMENT. Heart 2002, 88, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.; Ambrose, A.J. Understanding myocardial infarction. F1000Research 2018, 7, 1378. [Google Scholar] [CrossRef] [PubMed]
- Vanagt, W.Y.; Cornelussen, R.N.; Poulina, Q.P.; Blaauw, E.; Vernooy, K.; Cleutjens, J.P.; van Bilsen, M.; Delhaas, T.; Prinzen, F.W. Pacing-Induced Dys-Synchrony Preconditions Rabbit Myocardium Against Ischemia/Reperfusion Injury. Circulation 2006, 114, I-264–I-269. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.-Q.; Corvera, J.S.; Halkos, M.E.; Kerendi, F.; Wang, N.-P.; Guyton, R.A.; Vinten-Johansen, J. Inhibition of myocardial injury by ischemic postconditioning during reperfusion: Comparison with ischemic preconditioning. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H579–H588. [Google Scholar] [CrossRef]
- Reed, G.W.; Rossi, E.J.; Cannon, C.P. Acute myocardial infarction. Lancet 2016, 389, 197–210. [Google Scholar] [CrossRef]
- Babiker, F.; Al-Jarallah, A.; Al-Awadi, M. Effects of Cardiac Hypertrophy, Diabetes, Aging, and Pregnancy on the Cardioprotective Effects of Postconditioning in Male and Female Rats. Cardiol. Res. Pract. 2019, 2019, 3403959. [Google Scholar] [CrossRef]
- Babiker, F.A.; Al-Jarallah, A.; Joseph, S. Understanding pacing postconditioning-mediated cardiac protection: A role of oxidative stress and a synergistic effect of adenosine. J. Physiol. Biochem. 2016, 73, 175–185. [Google Scholar] [CrossRef]
- Hausenloy, D.J.; Yellon, D.M. Preconditioning and postconditioning: Underlying mechanisms and clinical application. Atherosclerosis 2009, 204, 334–341. [Google Scholar] [CrossRef]
- Akhtar, S.; Benter, I.F.; Danjuma, M.; Doi, S.A.R.; Hasan, S.S.; Habib, A.M. Pharmacotherapy in COVID-19 patients: A review of ACE2-raising drugs and their clinical safety. J. Drug Target. 2020, 28, 683–699. [Google Scholar] [CrossRef]
- Shraim, B.A.; Moursi, M.O.; Benter, I.F.; Habib, A.M.; Akhtar, S. The Role of Epidermal Growth Factor Receptor Family of Receptor Tyrosine Kinases in Mediating Diabetes-Induced Cardiovascular Complications. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef]
- Xie, J.X.; Hu, J.; Cheng, J.; Liu, C.; Wei, X. The function of the ACE2/Ang(1-7)/Mas receptor axis of the renin-angiotensin system in myocardial ischemia reperfusion injury. Eur Rev Med Pharmacol Sci. Mar 2022, 26, 1852–1859. [Google Scholar] [CrossRef]
- Vickers, C.; Hales, P.; Kaushik, V.; Dick, L.; Gavin, J.; Tang, J.; Godbout, K.; Parsons, T.; Baronas, E.; Hsieh, F.; et al. Hydrolysis of Biological Peptides by Human Angiotensin-converting Enzyme-related Carboxypeptidase. J. Biol. Chem. 2002, 277, 14838–14843. [Google Scholar] [CrossRef] [PubMed]
- Al-Maghrebi, M.; Benter, I.F.; Diz, D.I. Endogenous angiotensin-(1-7) reduces cardiac ischemia-induced dysfunction in diabetic hypertensive rats. Pharmacol. Res. 2009, 59, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Benter, I.F.; Yousif, M.; Al-Saleh, F.M.; Raghupathy, R.; Chappell, R.R.M.C.; Diz, D.I. Angiotensin-(1-7) Blockade Attenuates Captopril- or Hydralazine-induced Cardiovascular Protection in Spontaneously Hypertensive Rats Treated With NG-nitro-l-Arginine Methyl Ester. J. Cardiovasc. Pharmacol. 2011, 57, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Benter, I.F.; Yousif, M.; Anim, J.T.; Cojocel, C.; Diz, D.I. Angiotensin-(1-7) prevents development of severe hypertension and end-organ damage in spontaneously hypertensive rats treated with l-NAME. Am. J. Physiol. Circ. Physiol. 2006, 290, H684–H691. [Google Scholar] [CrossRef]
- Yousif, M.; Dhaunsi, G.S.; Makki, B.M.; Qabazard, B.A.; Akhtar, S.; Benter, I.F. Characterization of Angiotensin-(1-7) effects on the cardiovascular system in an experimental model of Type-1 diabetes. Pharmacol. Res. 2012, 66, 269–275. [Google Scholar] [CrossRef]
- Abwainy, A.; Babiker, F.; Akhtar, S.; Benter, I.F. Endogenous angiotensin-(1-7)/Mas receptor/NO pathway mediates the cardioprotective effects of pacing postconditioning. Am. J. Physiol. Circ. Physiol. 2016, 310, H104–H112. [Google Scholar] [CrossRef]
- Khalaf, A.; Babiker, F. Discrepancy in calcium release from the sarcoplasmic reticulum and intracellular acidic stores for the protection of the heart against ischemia/reperfusion injury. J. Physiol. Biochem. 2016, 72, 495–508. [Google Scholar] [CrossRef]
- Babiker, F.A.; Joseph, S.; Juggi, J. The protective effects of 17beta-estradiol against ischemia–reperfusion injury and its effect on pacing postconditioning protection to the heart. J. Physiol. Biochem. 2013, 70, 151–162. [Google Scholar] [CrossRef]
- Babiker, A.F.; Hoteit, L.J.; Joseph, S.; Mustafa, A.S.; Juggi, J.S. The role of 17-beta estradiol in ischemic preconditioning protection of the heart. Exp. Clin. Cardiol. 2012, 17, 95–100. [Google Scholar]
- Babiker, F.; Al-Kouh, A.; Kilarkaje, N. Lead exposure induces oxidative stress, apoptosis, and attenuates protection of cardiac myocytes against ischemia–reperfusion injury. Drug Chem. Toxicol. 2018, 42, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Al-Herz, W.; Babiker, F. Acute Intravenous Infusion of Immunoglobulins Protects Against Myocardial Ischemia-Reperfusion Injury Through Inhibition of Caspase-3. Cell. Physiol. Biochem. 2017, 42, 2295–2306. [Google Scholar] [CrossRef] [PubMed]
- Nigavekar, S.S.; Sung, L.Y.; Llanes, M.; El-Jawahri, A.; Lawrence, T.S.; Becker, C.W.; Balogh, L.; Khan, M.K. 3H Dendrimer Nanoparticle Organ/Tumor Distribution. Pharm. Res. 2004, 21, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Magruder, J.T.; Crawford, T.C.; Lin, Y.-A.; Zhang, F.; Grimm, J.C.; Kannan, R.M.; Kannan, S.; Sciortino, C.M. Selective Localization of a Novel Dendrimer Nanoparticle in Myocardial Ischemia-Reperfusion Injury. Ann. Thorac. Surg. 2017, 104, 891–898. [Google Scholar] [CrossRef]
- Shcharbin, D.; Janaszewska, A.; Klajnert-Maculewicz, B.; Ziemba, B.; Dzmitruk, V.; Halets, I.; Loznikova, S.; Shcharbina, N.; Milowska, K.; Ionov, M.; et al. How to study dendrimers and dendriplexes III. Biodistribution, pharmacokinetics and toxicity in vivo. J. Control. Release 2014, 181, 40–52. [Google Scholar] [CrossRef]
- Akhtar, S. Cationic nanosystems for the delivery of small interfering ribonucleic acid therapeutics: A focus on toxicogenomics. Expert Opin. Drug Metab. Toxicol. 2010, 6, 1347–1362. [Google Scholar] [CrossRef]
- Akhtar, S.; Benter, I. Toxicogenomics of non-viral drug delivery systems for RNAi: Potential impact on siRNA-mediated gene silencing activity and specificity. Adv. Drug Deliv. Rev. 2007, 59, 164–182. [Google Scholar] [CrossRef]
- Akhtar, S.; Chandrasekhar, B.; Attur, S.; Dhaunsi, G.S.; Yousif, M.H.M.; Benter, I.F. Transactivation of ErbB Family of Receptor Tyrosine Kinases Is Inhibited by Angiotensin-(1-7) via Its Mas Receptor. PLoS ONE 2015, 10, e0141657. [Google Scholar] [CrossRef]
- Akhtar, S.; Chandrasekhar, B.; Attur, S.; Yousif, M.; Benter, I.F. On the nanotoxicity of PAMAM dendrimers: Superfect® stimulates the EGFR–ERK1/2 signal transduction pathway via an oxidative stress-dependent mechanism in HEK 293 cells. Int. J. Pharm. 2013, 448, 239–246. [Google Scholar] [CrossRef]
- Hotta, H.; Miura, T.; Miki, T.; Togashi, N.; Maeda, T.; Kim, S.J.; Tanno, M.; Yano, T.; Kuno, A.; Itoh, T.; et al. Short Communication: Angiotensin II Type 1 Receptor–Mediated Upregulation of Calcineurin Activity Underlies Impairment of Cardioprotective Signaling in Diabetic Hearts. Circ. Res. 2010, 106, 129–132. [Google Scholar] [CrossRef]
- Omidi, Y.; Barar, J.; Heidari, H.R.; Ahmadian, S.; Yazdi, H.A.; Akhtar, S. Microarray Analysis of the Toxicogenomics and the Genotoxic Potential of a Cationic Lipid-Based Gene Delivery Nanosystem in Human Alveolar Epithelial A549 Cells. Toxicol. Mech. Methods 2008, 18, 369–378. [Google Scholar] [CrossRef]
- Omidi, Y.; Hollins, A.J.; Benboubetra, M.; Drayton, R.; Benter, I.F.; Akhtar, S. Toxicogenomics of Non-viral Vectors for Gene Therapy: A Microarray Study of Lipofectin- and Oligofectamine-induced Gene Expression Changes in Human Epithelial Cells. J. Drug Target. 2003, 11, 311–323. [Google Scholar] [CrossRef]
- Omidi, Y.; Hollins, A.; Drayton, R.; Akhtar, S. Polypropylenimine dendrimer-induced gene expression changes: The effect of complexation with DNA, dendrimer generation and cell type. J. Drug Target. 2005, 13, 431–443. [Google Scholar] [CrossRef]
- Kumagai, S.; Koyama, S.; Nishikawa, H. Antitumour immunity regulated by aberrant ERBB family signalling. Nat. Rev. Cancer 2021, 21, 181–197. [Google Scholar] [CrossRef]
- Hajjo, R.; Sweidan, K. Review on Epidermal Growth Factor Receptor (EGFR) Structure, Signaling Pathways, Interactions, and Recent Updates of EGFR Inhibitors. Curr. Top. Med. Chem. 2020, 20, 815–834. [Google Scholar] [CrossRef]
- Sharifi, J.; Khirehgesh, M.R.; Safari, F.; Akbari, B. EGFR and anti-EGFR nanobodies: Review and update. J. Drug Target. 2020, 29, 387–402. [Google Scholar] [CrossRef]
- Benter, I.F.; Juggi, J.S.; Khan, I.; Yousif, M.; Canatan, H.; Akhtar, S. Signal transduction mechanisms involved in cardiac preconditioning: Role of Ras-GTPase, Ca2 +/calmodulin-dependent protein kinase II and epidermal growth factor receptor. Mol. Cell. Biochem. 2005, 268, 175–183. [Google Scholar] [CrossRef]
- Akhtar, S.; Yousif, M.H.M.; Chandrasekhar, B.; Benter, I.F. Activation of EGFR/ERBB2 via Pathways Involving ERK1/2, P38 MAPK, AKT and FOXO Enhances Recovery of Diabetic Hearts from Ischemia-Reperfusion Injury. PLoS ONE 2012, 7, e39066. [Google Scholar] [CrossRef]
- Benter, I.F.; Juggi, J.S.; Khan, I.; Akhtar, S. Inhibition of Ras-GTPase, but not tyrosine kinases or Ca2+/calmodulin-dependent protein kinase II, improves recovery of cardiac function in the globally ischemic heart. Mol. Cell. Biochem. 2004, 259, 35–42. [Google Scholar] [CrossRef]
- Jones, C.F.; Campbell, R.A.; Brooks, A.E.; Assemi, S.; Tadjiki, S.; Thiagarajan, G.; Mulcock, C.; Weyrich, A.S.; Brooks, B.D.; Ghandehari, H.; et al. Cationic PAMAM Dendrimers Aggressively Initiate Blood Clot Formation. ACS Nano 2012, 6, 9900–9910. [Google Scholar] [CrossRef]
- Chauhan, A.S.; Diwan, P.V.; Jain, N.K.; Tomalia, D.A. Unexpected In Vivo Anti-Inflammatory Activity Observed for Simple, Surface Functionalized Poly(amidoamine) Dendrimers. Biomacromolecules 2009, 10, 1195–1202. [Google Scholar] [CrossRef]
- Labieniec-Watala, M.; Watala, C. PAMAM Dendrimers: Destined for Success or Doomed to Fail? Plain and Modified PAMAM Dendrimers in the Context of Biomedical Applications. J. Pharm. Sci. 2015, 104, 2–14. [Google Scholar] [CrossRef]
- Benter, I.F.; Yousif, M.; Cojocel, C.; Al-Maghrebi, M.; Diz, D.I. Angiotensin-(1-7) prevents diabetes-induced cardiovascular dysfunction. Am. J. Physiol. Circ. Physiol. 2007, 292, H666–H672. [Google Scholar] [CrossRef]
- El-Hashim, A.Z.; Renno, W.M.; Raghupathy, R.; Abduo, H.T.; Akhtar, S.; Benter, I.F. Angiotensin-(1-7) inhibits allergic inflammation, via the MAS1 receptor, through suppression of ERK1/2- and NF-κB-dependent pathways. J. Cereb. Blood Flow Metab. 2012, 166, 1964–1976. [Google Scholar] [CrossRef]
- Rahimi, O.; Melo, A.C.; Westwood, B.; Grier, R.D.; Tallant, E.A.; Gallagher, P.E. Angiotensin-(1-7) reduces doxorubicin-induced aortic arch dysfunction in male and female juvenile Sprague Dawley rats through pleiotropic mechanisms. Peptides 2022, 152, 170784. [Google Scholar] [CrossRef]
- Pacheco, L.F.; de Castro, C.H.; Dutra, J.B.R.; Lino, R.D.S.; Ferreira, P.M.; dos Santos, R.A.S.; Ulhoa, C.J. Oral Treatment with Angiotensin-(1-7) Attenuates the Kidney Injury Induced by Gentamicin in Wistar Rats. Protein Pept. Lett. 2021, 28, 1425–1433. [Google Scholar] [CrossRef]
- Chi, L.A.; Asgharpour, S.; Correa-Basurto, J.; Bandala, C.R.; Martínez-Archundia, M. Unveiling the G4-PAMAM capacity to bind and protect Ang-(1-7) bioactive peptide by molecular dynamics simulations. J. Comput. Mol. Des. 2022, 36, 653–675. [Google Scholar] [CrossRef]
- Magalhães, G.S.; Gregório, J.F.; Ramos, K.E.; Cançado-Ribeiro, A.T.P.; Baroni, I.F.; Barcelos, L.S.; Pinho, V.; Teixeira, M.M.; Santos, R.A.S.; Rodrigues-Machado, M.G.; et al. Treatment with inhaled formulation of angiotensin-(1-7) reverses inflammation and pulmonary remodeling in a model of chronic asthma. Immunobiology 2020, 225, 151957. [Google Scholar] [CrossRef]
- Márquez-Miranda, V.; Abrigo, J.; Rivera, J.C.; Araya-Duran, I.; Aravena, J.; Simon, F.; Pacheco, N.; Gonzalez-Nilo, F.D.; Cabello-Verrugio, C. The complex of PAMAM-OH dendrimer with Angiotensin (1-7) prevented the disuse-induced skeletal muscle atrophy in mice. Int. J. Nanomed. 2017, 12, 1985–1999. [Google Scholar] [CrossRef]
- Rodgers, K.E.; Oliver, J.; Dizerega, G.S. Phase I/II dose escalation study of angiotensin 1-7 [A(1-7)] administered before and after chemotherapy in patients with newly diagnosed breast cancer. Cancer Chemother. Pharmacol. 2005, 57, 559–568. [Google Scholar] [CrossRef]
- Savage, P.D.; Lovato, J.; Brosnihan, K.B.; Miller, A.A.; Petty, W.J. Phase II Trial of Angiotensin-(1-7) for the Treatment of Patients with Metastatic Sarcoma. Sarcoma 2016, 2016, 4592768. [Google Scholar] [CrossRef] [PubMed]
- Boarescu, P.M.; Boarescu, I.; Bocșan, I.C.; Gheban, D.; Bulboacă, A.E.; Nicula, C.; Pop, R.M.; Râjnoveanu, R.-M.; Bolboacă, S.D. Antioxidant and Anti-Inflammatory Effects of Curcumin Nanoparticles on Drug-Induced Acute Myocardial Infarction in Diabetic Rats. Antioxidants 2019, 8, 504. [Google Scholar] [CrossRef] [PubMed]
- Bulboacă, A.E.; Boarescu, P.M.; Bolboacă, S.D.; Blidaru, M.; Feștilă, D.; Dogaru, G.; Nicula, C.A. Comparative effect of curcumin versus liposomal curcumin on systemic pro-inflammatory cytokines profile, MCP-1 and RANTES in experimental diabetes mellitus. Int. J. Nanomed. 2019, 14, 8961–8972. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Terminal Surface Chemistry $ | Surface Charge | Generation $ | Molecular Weight (Da) $ | Diameter (nm) $ | No. of Surface Groups $ | Dose (s) Administered in Isolated Rat Heart |
|---|---|---|---|---|---|---|
| -NH2 | Cationic | 3 | 6909 | 3.6 | 32 | 100 nM |
| -NH2 | Cationic | 4 | 14,215 | 4.5 | 64 | 100 nM |
| -NH2 | Cationic | 5 | 28,826 | 5.4 | 128 | 100 nM |
| -NH2 | Cationic | 6 | 58,048 | 6.7 | 256 | 100 nM |
| -NH2 | Cationic | 7 | 116,493 | 8.1 | 512 | 100 nM and for dose-dependent studies: 1 µg, 5 µg, 7.5 µg, 10 µg or 20 µg/mL |
| -OH | Neutral | 6 | 58,304 | NA | 256 | 100 nM |
| -COOH | Anionic | 5.5 | 52,913 | NA | 256 | 100 nM |
| Treatment | CK (IU/L) | p Value | LDH (IU/L) | p Value |
|---|---|---|---|---|
| I/R | 35.77 ± 0.46 | - | 28.27 ± 1.12 | - |
| Ang-(1-7) | 26.97 ± 1.43 * | 0.001 | 20.24 ± 0.39 * | 0.001 |
| G7 | 46.26 ± 1.39 * | 0.022 | 40.41 ± 0.99 * | 0.01 |
| G7 + Ang-(1-7) | 28.34 ± 0.86 * | 0.002 | 21.73 ± 0.73 * | 0.001 |
| G7 + Ang-(1-7) + DPro | 35.22 ± 1.21 | 0.709 | 29.40 ± 0.76 | 0.294 |
| G7 + Ang-(1-7) + A779 | 35.32 ± 0.96 | 0.633 | 29.33 ± 0.39 | 0.383 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhtar, S.; Babiker, F.; Akhtar, U.A.; Benter, I.F. Mitigating Cardiotoxicity of Dendrimers: Angiotensin-(1-7) via Its Mas Receptor Ameliorates PAMAM-Induced Cardiac Dysfunction in the Isolated Mammalian Heart. Pharmaceutics 2022, 14, 2673. https://doi.org/10.3390/pharmaceutics14122673
Akhtar S, Babiker F, Akhtar UA, Benter IF. Mitigating Cardiotoxicity of Dendrimers: Angiotensin-(1-7) via Its Mas Receptor Ameliorates PAMAM-Induced Cardiac Dysfunction in the Isolated Mammalian Heart. Pharmaceutics. 2022; 14(12):2673. https://doi.org/10.3390/pharmaceutics14122673
Chicago/Turabian StyleAkhtar, Saghir, Fawzi Babiker, Usman A. Akhtar, and Ibrahim F. Benter. 2022. "Mitigating Cardiotoxicity of Dendrimers: Angiotensin-(1-7) via Its Mas Receptor Ameliorates PAMAM-Induced Cardiac Dysfunction in the Isolated Mammalian Heart" Pharmaceutics 14, no. 12: 2673. https://doi.org/10.3390/pharmaceutics14122673
APA StyleAkhtar, S., Babiker, F., Akhtar, U. A., & Benter, I. F. (2022). Mitigating Cardiotoxicity of Dendrimers: Angiotensin-(1-7) via Its Mas Receptor Ameliorates PAMAM-Induced Cardiac Dysfunction in the Isolated Mammalian Heart. Pharmaceutics, 14(12), 2673. https://doi.org/10.3390/pharmaceutics14122673