
Effect of Selected Antidepressants on Placental Homeostasis of Serotonin: Maternal and Fetal Perspectives

 , , ,
, , , 
Abstract
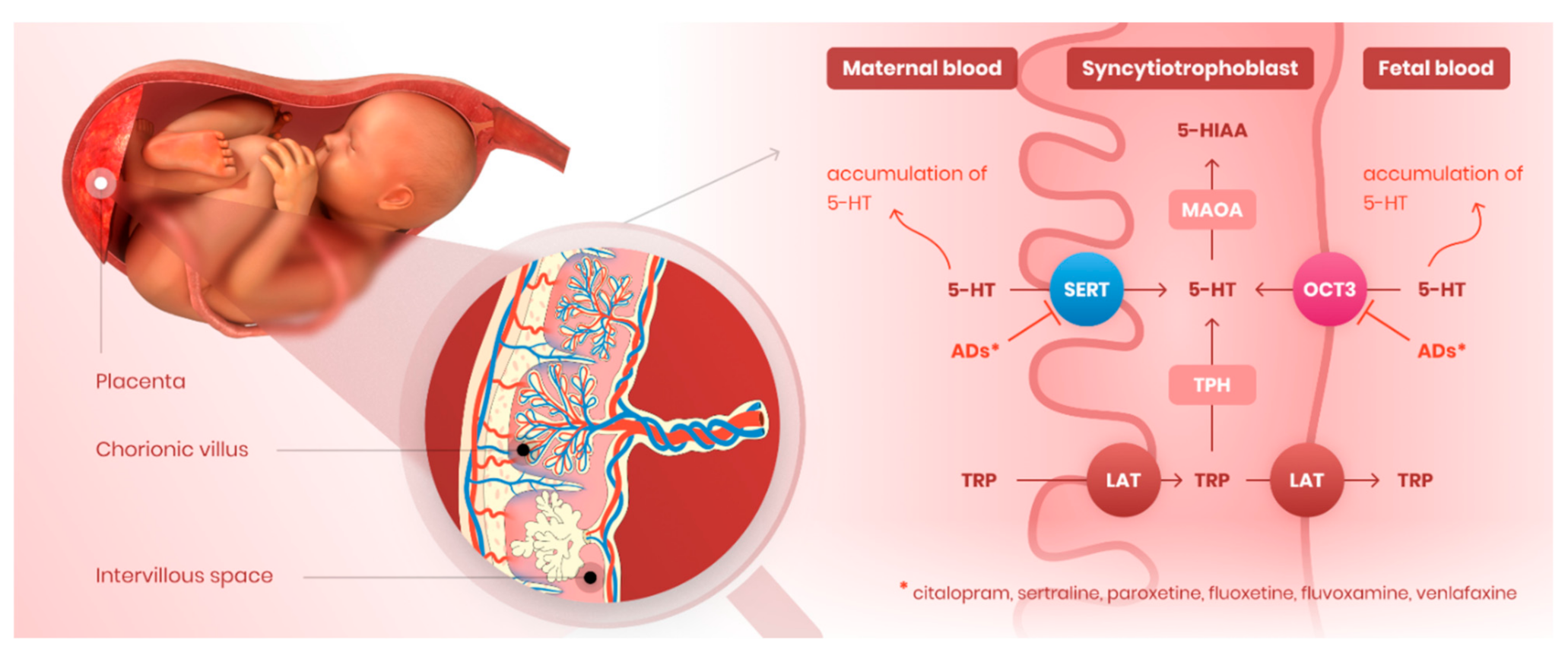
1. Introduction
2. Materials and Methods
2.1. Materials
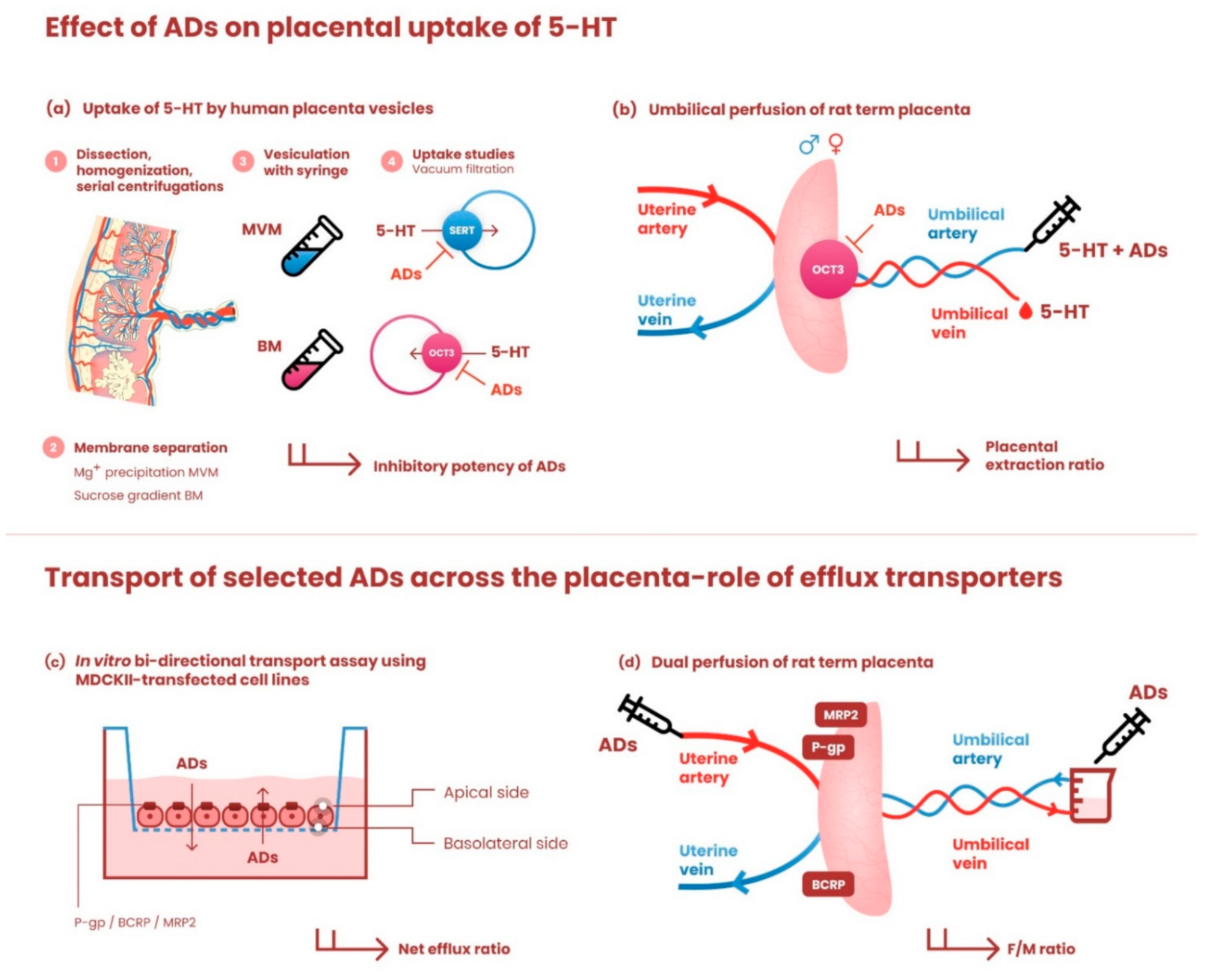
2.2. Preparation of Microvillous and Basal Membrane Vesicles from Human Term Placenta
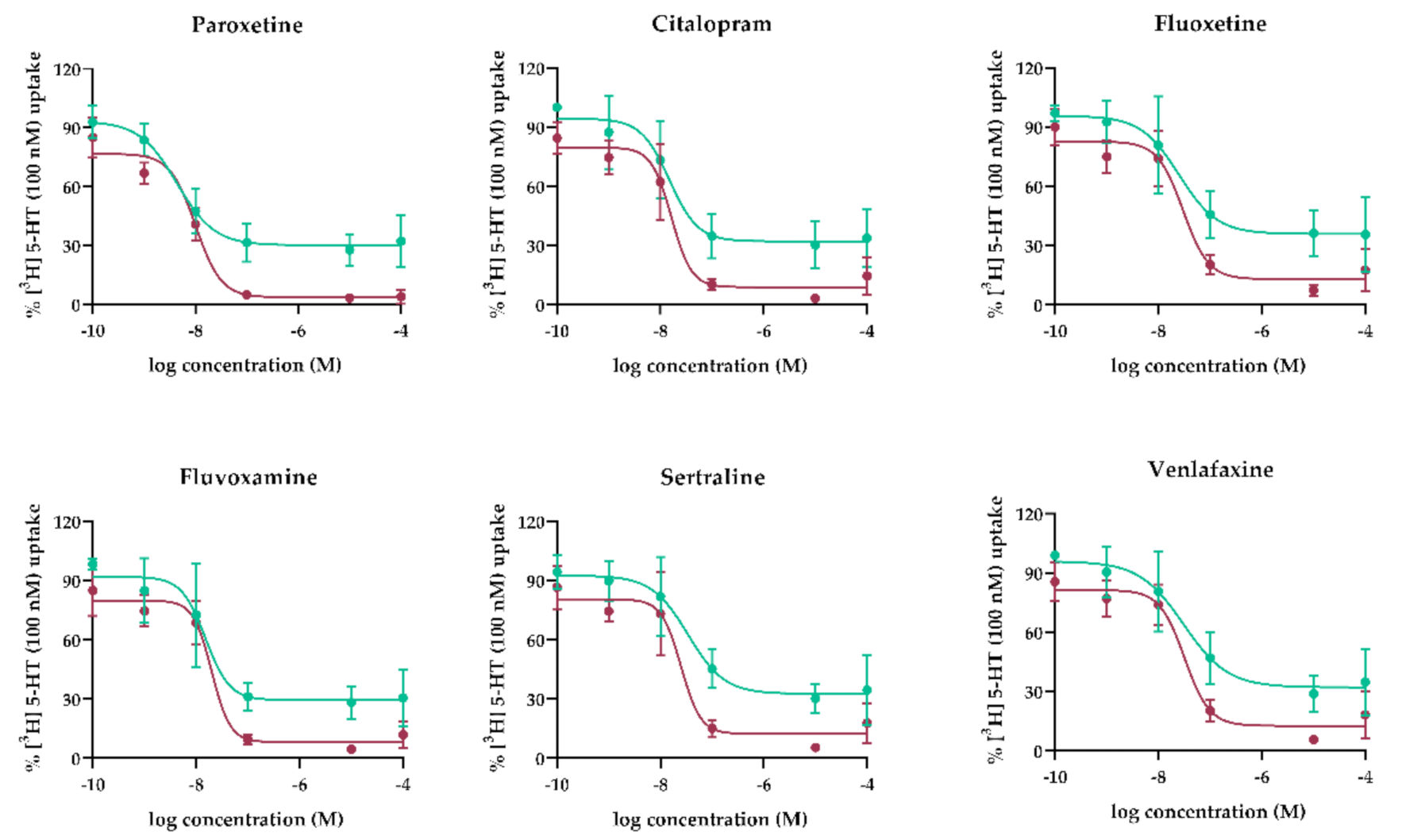
Concentration-Dependent Inhibition by Antidepressants of 5-HT Uptake by MVM and BM Vesicles
2.3. In Situ Perfusion of Rat Term Placenta
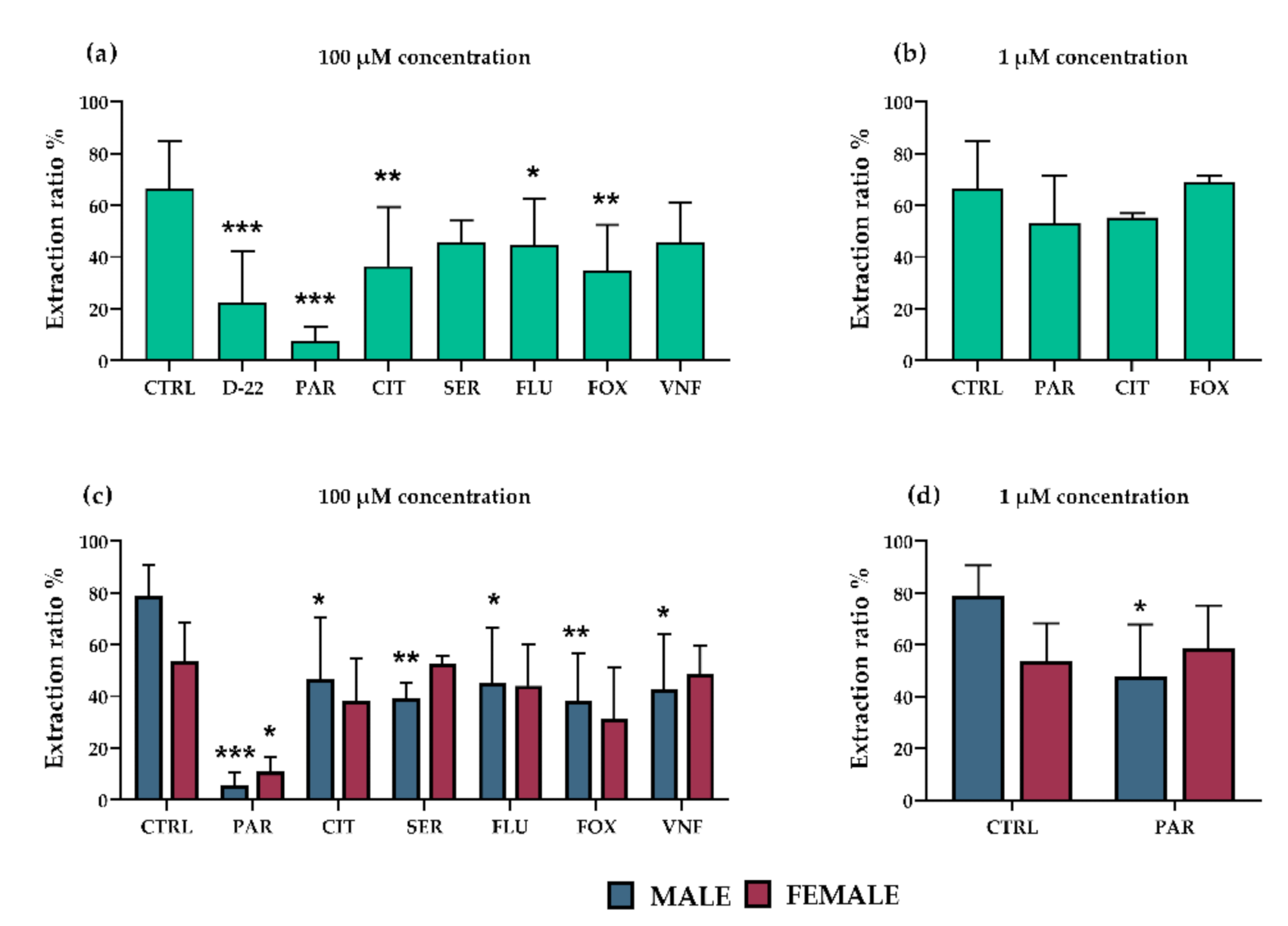
2.3.1. Effect of Antidepressants on Placental Extraction of 5-HT from Fetal Circulation; Influence of Fetal Sex
2.3.2. Effect of Efflux Transporters on Placental Transport of Antidepressants
2.4. RNA Isolation from Rat Placental Tissue
2.5. Reverse Transcription and Droplet Digital PCR Assay
2.6. Western Blot Analysis
2.7. Evaluation of MAO-A Activity and Lipid Peroxidation in Rat Placenta Homogenates
2.8. Cells
In Vitro Bidirectional Transport of Selected Antidepressants
2.9. Radioisotope Analysis
2.10. Statistical Analysis
3. Results
3.1. Concentration-Dependent Effect of Antidepressants on SERT- and OCT3-Mediated 5-HT Uptake by Human MVM and BM Vesicles
3.2. Effect of Antidepressants on OCT3-Mediated 5-HT Extraction from Fetal Circulation in Perfused Rat Term Placenta
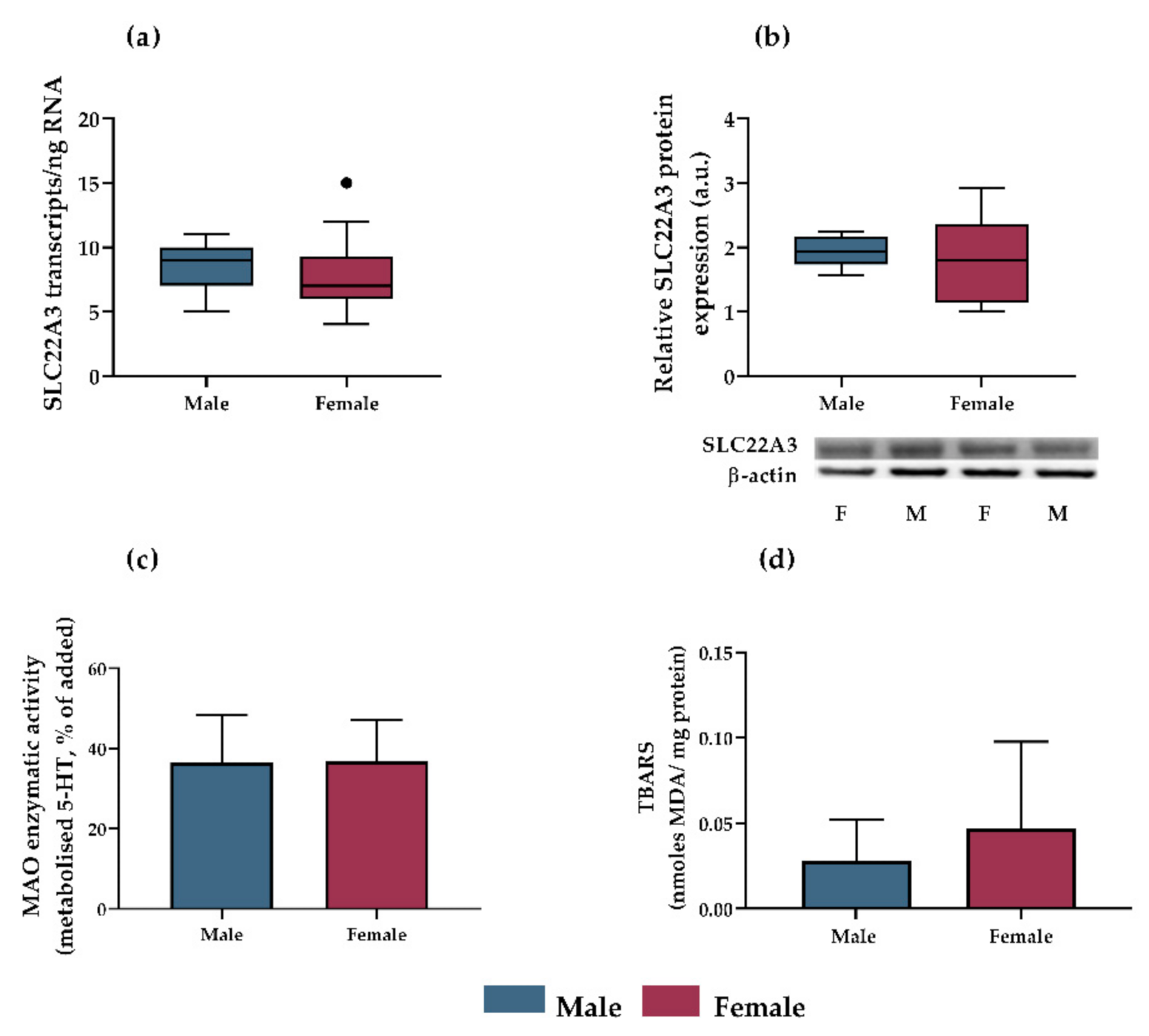
3.3. OCT3 Expression, MAO-A Activity and Lipid Peroxidation as Potential Mechanisms Accounting for Sex-Dependent Inhibition of Antidepressants on Serotonin Uptake in Rat Placenta
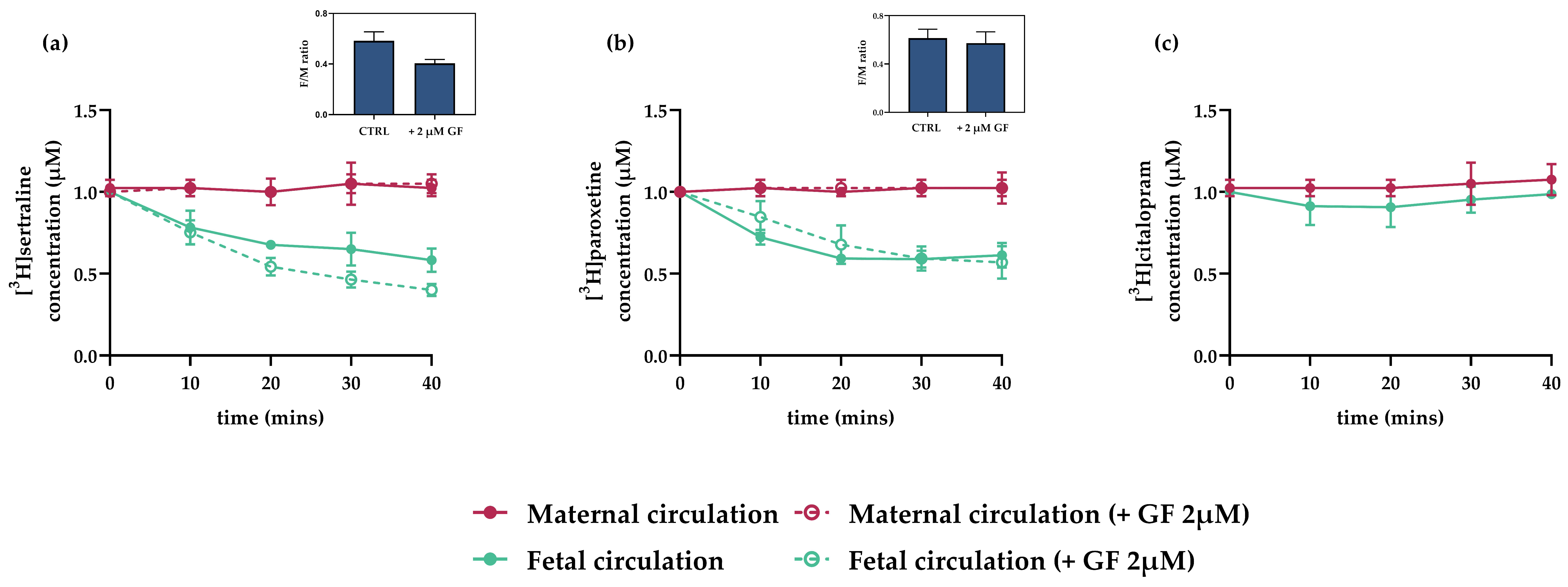
3.4. Interactions of Antidepressants with Efflux Transporters In Vitro
3.5. Interactions of Antidepressants with Efflux Transporters in Rat Term Placenta
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pawluski, J.L.; Lonstein, J.S.; Fleming, A.S. The Neurobiology of Postpartum Anxiety and Depression. Trends Neurosci. 2017, 40, 106–120. [Google Scholar] [CrossRef]
- Hagberg, K.W.; Robijn, A.L.; Jick, S. Maternal depression and antidepressant use during pregnancy and the risk of autism spectrum disorder in offspring. Clin. Epidemiol. 2018, 10, 1599–1612. [Google Scholar] [CrossRef] [PubMed]
- Cooper, W.O.; Willy, M.E.; Pont, S.J.; Ray, W.A. Increasing use of antidepressants in pregnancy. Am. J. Obstet. Gynecol. 2007, 196, 544.e1-5. [Google Scholar] [CrossRef] [PubMed]
- Charlton, R.A.; Jordan, S.; Pierini, A.; Garne, E.; Neville, A.J.; Hansen, A.V.; Gini, R.; Thayer, D.; Tingay, K.; Puccini, A.; et al. Selective serotonin reuptake inhibitor prescribing before, during and after pregnancy: A population-based study in six European regions. Bjog 2015, 122, 1010–1020. [Google Scholar] [CrossRef]
- Oberlander, T.F.; Gingrich, J.A.; Ansorge, M.S. Sustained neurobehavioral effects of exposure to SSRI antidepressants during development: Molecular to clinical evidence. Clin. Pharmacol. Ther. 2009, 86, 672–677. [Google Scholar] [CrossRef]
- Videman, M.; Tokariev, A.; Saikkonen, H.; Stjerna, S.; Heiskala, H.; Mantere, O.; Vanhatalo, S. Newborn Brain Function Is Affected by Fetal Exposure to Maternal Serotonin Reuptake Inhibitors. Cereb. Cortex 2017, 27, 3208–3216. [Google Scholar] [CrossRef]
- Bonnin, A.; Levitt, P. Placental Source for 5-HT that Tunes Fetal Brain Development. Neuropsychopharmacology 2012, 37, 299–300. [Google Scholar] [CrossRef]
- Maroteaux, L.; Kilic, F. Frontiers of Serotonin Beyond the Brain. Pharmacol. Res. 2019, 140, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Staud, F.; Karahoda, R. Trophoblast: The central unit of fetal growth, protection and programming. Int. J. Biochem. Cell Biol. 2018, 105, 35–40. [Google Scholar] [CrossRef]
- Malm, H.; Brown, A.S.; Gissler, M.; Gyllenberg, D.; Hinkka-Yli-Salomäki, S.; McKeague, I.W.; Weissman, M.; Wickramaratne, P.; Artama, M.; Gingrich, J.A.; et al. Gestational Exposure to Selective Serotonin Reuptake Inhibitors and Offspring Psychiatric Disorders: A National Register-Based Study. J. Am. Acad. Child Adolesc. Psychiatry 2016, 55, 359–366. [Google Scholar] [CrossRef]
- Liu, X.; Agerbo, E.; Ingstrup, K.G.; Musliner, K.; Meltzer-Brody, S.; Bergink, V.; Munk-Olsen, T. Antidepressant use during pregnancy and psychiatric disorders in offspring: Danish nationwide register based cohort study. BMJ 2017, 358, j3668. [Google Scholar] [CrossRef]
- Viktorin, A.; Uher, R.; Reichenberg, A.; Levine, S.Z.; Sandin, S. Autism risk following antidepressant medication during pregnancy. Psychol. Med. 2017, 47, 2787–2796. [Google Scholar] [CrossRef]
- Morales, D.R.; Slattery, J.; Evans, S.; Kurz, X. Antidepressant use during pregnancy and risk of autism spectrum disorder and attention deficit hyperactivity disorder: Systematic review of observational studies and methodological considerations. BMC Med. 2018, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Boukhris, T.; Sheehy, O.; Mottron, L.; Berard, A. Antidepressant Use During Pregnancy and the Risk of Autism Spectrum Disorder in Children. JAMA Pediatr. 2016, 170, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Gidaya, N.B.; Lee, B.K.; Burstyn, I.; Yudell, M.; Mortensen, E.L.; Newschaffer, C.J. In utero exposure to selective serotonin reuptake inhibitors and risk for autism spectrum disorder. J. Autism Dev. Disord. 2014, 44, 2558–2567. [Google Scholar] [CrossRef] [PubMed]
- Sato, K. Placenta-derived hypo-serotonin situations in the developing forebrain cause autism. Med. Hypotheses 2013, 80, 368–372. [Google Scholar] [CrossRef]
- Yang, C.J.; Tan, H.P.; Du, Y.J. The developmental disruptions of serotonin signaling may involved in autism during early brain development. Neuroscience 2014, 267, 1–10. [Google Scholar] [CrossRef]
- Pawluski, J.L.; Rayen, I.; Niessen, N.A.; Kristensen, S.; van Donkelaar, E.L.; Balthazart, J.; Steinbusch, H.W.; Charlier, T.D. Developmental fluoxetine exposure differentially alters central and peripheral measures of the HPA system in adolescent male and female offspring. Neuroscience 2012, 220, 131–141. [Google Scholar] [CrossRef]
- Palmsten, K.; Huybrechts, K.F.; Michels, K.B.; Williams, P.L.; Mogun, H.; Setoguchi, S.; Hernández-Díaz, S. Antidepressant use and risk for preeclampsia. Epidemiology 2013, 24, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Bernard, N.; Forest, J.-C.; Tarabulsy, G.M.; Bujold, E.; Bouvier, D.; Giguère, Y. Use of antidepressants and anxiolytics in early pregnancy and the risk of preeclampsia and gestational hypertension: A prospective study. BMC Pregnancy Childbirth 2019, 19, 146. [Google Scholar] [CrossRef] [PubMed]
- Marchocki, Z.; Russell, N.E.; Donoghue, K.O. Selective serotonin reuptake inhibitors and pregnancy: A review of maternal, fetal and neonatal risks and benefits. Obstet. Med. 2013, 6, 155–158. [Google Scholar] [CrossRef]
- Fornaro, E.; Li, D.; Pan, J.; Belik, J. Prenatal exposure to fluoxetine induces fetal pulmonary hypertension in the rat. Am. J. Respir. Crit. Care Med. 2007, 176, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Harrington, R.A.; Lee, L.C.; Crum, R.M.; Zimmerman, A.W.; Hertz-Picciotto, I. Prenatal SSRI use and offspring with autism spectrum disorder or developmental delay. Pediatrics 2014, 133, e1241–e1248. [Google Scholar] [CrossRef]
- Bérard, A.; Zhao, J.P.; Sheehy, O. Antidepressant use during pregnancy and the risk of major congenital malformations in a cohort of depressed pregnant women: An updated analysis of the Quebec Pregnancy Cohort. BMJ Open 2017, 7, e013372. [Google Scholar] [CrossRef]
- Diav-Citrin, O.; Shechtman, S.; Weinbaum, D.; Wajnberg, R.; Avgil, M.; Di Gianantonio, E.; Clementi, M.; Weber-Schoendorfer, C.; Schaefer, C.; Ornoy, A. Paroxetine and fluoxetine in pregnancy: A prospective, multicentre, controlled, observational study. Br. J. Clin. Pharm. 2008, 66, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.-Y.; Wu, Q.-J.; Sun, C.; Zhang, T.-N.; Shen, Z.-Q.; Liu, C.-X.; Gong, T.-T.; Xu, X.; Ji, C.; Huang, D.-H.; et al. Selective serotonin reuptake inhibitor use during early pregnancy and congenital malformations: A systematic review and meta-analysis of cohort studies of more than 9 million births. BMC Med. 2018, 16, 205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.N.; Gao, S.Y.; Shen, Z.Q.; Li, D.; Liu, C.X.; Lv, H.C.; Zhang, Y.; Gong, T.T.; Xu, X.; Ji, C.; et al. Use of selective serotonin-reuptake inhibitors in the first trimester and risk of cardiovascular-related malformations: A meta-analysis of cohort studies. Sci. Rep. 2017, 7, 43085. [Google Scholar] [CrossRef] [PubMed]
- Nordeng, H.; Lindemann, R.; Perminov, K.V.; Reikvam, A. Neonatal withdrawal syndrome after in utero exposure to selective serotonin reuptake inhibitors. Acta Paediatr. 2001, 90, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Jensen, H.M.; Grøn, R.; Lidegaard, Ø.; Pedersen, L.H.; Andersen, P.K.; Kessing, L.V. Maternal depression, antidepressant use in pregnancy and Apgar scores in infants. Br. J. Psychiatry 2013, 202, 347–351. [Google Scholar] [CrossRef]
- Simon, G.E.; Cunningham, M.L.; Davis, R.L. Outcomes of prenatal antidepressant exposure. Am. J. Psychiatry 2002, 159, 2055–2061. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, Q.; Cao, S.; Pang, J.; Zhang, H.; Feng, T.; Deng, Y.; Yao, J.; Li, H. A meta-analysis of selective serotonin reuptake inhibitors (SSRIs) use during prenatal depression and risk of low birth weight and small for gestational age. J. Affect Disord. 2018, 241, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Karahoda, R.; Horackova, H.; Kastner, P.; Matthios, A.; Cerveny, L.; Kucera, R.; Kacerovsky, M.; Duintjer Tebbens, J.; Bonnin, A.; Abad, C.; et al. Serotonin homeostasis in the materno-foetal interface at term: Role of transporters (SERT/SLC6A4 and OCT3/SLC22A3) and monoamine oxidase A (MAO-A) in uptake and degradation of serotonin by human and rat term placenta. Acta Physiol. 2020, 229, e13478. [Google Scholar] [CrossRef] [PubMed]
- Karahoda, R.; Abad, C.; Horackova, H.; Kastner, P.; Zaugg, J.; Cerveny, L.; Kucera, R.; Albrecht, C.; Staud, F. Dynamics of Tryptophan Metabolic Pathways in Human Placenta and Placental-Derived Cells: Effect of Gestation Age and Trophoblast Differentiation. Front. Cell Dev. Biol. 2020, 8, 574034. [Google Scholar] [CrossRef]
- Abad, C.; Karahoda, R.; Kastner, P.; Portillo, R.; Horackova, H.; Kucera, R.; Nachtigal, P.; Staud, F. Profiling of Tryptophan Metabolic Pathways in the Rat Fetoplacental Unit During Gestation. Int. J. Mol. Sci. 2020, 21, 7578. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, A.; Goeden, N.; Chen, K.; Wilson, M.L.; King, J.; Shih, J.C.; Blakely, R.D.; Deneris, E.S.; Levitt, P. A transient placental source of serotonin for the fetal forebrain. Nature 2011, 472, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Sano, M.; Ferchaud-Roucher, V.; Kaeffer, B.; Poupeau, G.; Castellano, B.; Darmaun, D. Maternal and fetal tryptophan metabolism in gestating rats: Effects of intrauterine growth restriction. Amino Acids 2016, 48, 281–290. [Google Scholar] [CrossRef]
- Arevalo, R.; Afonso, D.; Castro, R.; Rodriguez, M. Fetal brain serotonin synthesis and catabolism is under control by mother intake of tryptophan. Life Sci. 1991, 49, 53–66. [Google Scholar] [CrossRef]
- Zhu, H.-J.; Appel, D.I.; Gründemann, D.; Richelson, E.; Markowitz, J.S. Evaluation of organic cation transporter 3 (SLC22A3) inhibition as a potential mechanism of antidepressant action. Pharmacol. Res. 2012, 65, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Staud, F.; Cerveny, L.; Ceckova, M. Pharmacotherapy in pregnancy; effect of ABC and SLC transporters on drug transport across the placenta and fetal drug exposure. J. Drug Target. 2012, 20, 736–763. [Google Scholar] [CrossRef] [PubMed]
- Illsley, N.P.; Wang, Z.Q.; Gray, A.; Sellers, M.C.; Jacobs, M.M. Simultaneous preparation of paired, syncytial, microvillous and basal membranes from human placenta. Biochim. Biophys. Acta 1990, 1029, 218–226. [Google Scholar] [CrossRef]
- Staud, F.; Vackova, Z.; Pospechova, K.; Pavek, P.; Ceckova, M.; Libra, A.; Cygalova, L.; Nachtigal, P.; Fendrich, Z. Expression and transport activity of breast cancer resistance protein (Bcrp/Abcg2) in dually perfused rat placenta and HRP-1 cell line. J. Pharmacol. Exp. Ther. 2006, 319, 53–62. [Google Scholar] [CrossRef]
- Karbanova, S.; Cerveny, L.; Jiraskova, L.; Karahoda, R.; Ceckova, M.; Ptackova, Z.; Staud, F. Transport of ribavirin across the rat and human placental barrier: Roles of nucleoside and ATP-binding cassette drug efflux transporters. Biochem. Pharmacol. 2019, 163, 60–70. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Feix, J.B.; Bachowski, G.J.; Girotti, A.W. Photodynamic action of merocyanine 540 on erythrocyte membranes: Structural perturbation of lipid and protein constituents. Biochim. Biophys. Acta 1991, 1075, 28–35. [Google Scholar] [CrossRef]
- Neumanova, Z.; Cerveny, L.; Ceckova, M.; Staud, F. Interactions of tenofovir and tenofovir disoproxil fumarate with drug efflux transporters ABCB1, ABCG2, and ABCC2; role in transport across the placenta. Aids 2014, 28, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Neumanova, Z.; Cerveny, L.; Greenwood, S.L.; Ceckova, M.; Staud, F. Effect of drug efflux transporters on placental transport of antiretroviral agent abacavir. Reprod. Toxicol. 2015, 57, 176–182. [Google Scholar] [CrossRef]
- Martinec, O.; Huliciak, M.; Staud, F.; Cecka, F.; Vokral, I.; Cerveny, L. Anti-HIV and Anti-Hepatitis C Virus Drugs Inhibit P-Glycoprotein Efflux Activity in Caco-2 Cells and Precision-Cut Rat and Human Intestinal Slices. Antimicrob. Agents Chemother. 2019, 63, e00910-00919. [Google Scholar] [CrossRef] [PubMed]
- Eap, C.B.; Bouchoux, G.; Amey, M.; Cochard, N.; Savary, L.; Baumann, P. Simultaneous determination of human plasma levels of citalopram, paroxetine, sertraline, and their metabolites by gas chromatography-mass spectrometry. J. Chromatogr. Sci. 1998, 36, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Reis, M.; Aamo, T.; Spigset, O.; Ahlner, J. Serum concentrations of antidepressant drugs in a naturalistic setting: Compilation based on a large therapeutic drug monitoring database. Ther. Drug Monit. 2009, 31, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Baumann, P. Pharmacokinetic-pharmacodynamic relationship of the selective serotonin reuptake inhibitors. Clin. Pharm. 1996, 31, 444–469. [Google Scholar] [CrossRef] [PubMed]
- Ewing, G.; Tatarchuk, Y.; Appleby, D.; Schwartz, N.; Kim, D. Placental transfer of antidepressant medications: Implications for postnatal adaptation syndrome. Clin. Pharmacokinet. 2015, 54, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Chiarello, D.I.; Abad, C.; Rojas, D.; Toledo, F.; Vázquez, C.M.; Mate, A.; Sobrevia, L.; Marín, R. Oxidative stress: Normal pregnancy versus preeclampsia. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165354. [Google Scholar] [CrossRef] [PubMed]
- Dubovicky, M.; Belovicova, K.; Csatlosova, K.; Bogi, E. Risks of using SSRI / SNRI antidepressants during pregnancy and lactation. Interdiscip. Toxicol. 2017, 10, 30–34. [Google Scholar] [CrossRef]
- Corti, S.; Pileri, P.; Mazzocco, M.I.; Mandò, C.; Moscatiello, A.F.; Cattaneo, D.; Cheli, S.; Baldelli, S.; Pogliani, L.; Clementi, E.; et al. Neonatal Outcomes in Maternal Depression in Relation to Intrauterine Drug Exposure. Front. Pediatrics. 2019, 7, 309. [Google Scholar] [CrossRef] [PubMed]
- Ng, Q.X.; Venkatanarayanan, N.; Ho, C.Y.X.; Sim, W.S.; Lim, D.Y.; Yeo, W.S. Selective Serotonin Reuptake Inhibitors and Persistent Pulmonary Hypertension of the Newborn: An Update Meta-Analysis. J. Womens Health 2019, 28, 331–338. [Google Scholar] [CrossRef]
- Man, K.K.C.; Tong, H.H.Y.; Wong, L.Y.L.; Chan, E.W.; Simonoff, E.; Wong, I.C.K. Exposure to selective serotonin reuptake inhibitors during pregnancy and risk of autism spectrum disorder in children: A systematic review and meta-analysis of observational studies. Neurosci. Biobehav. Rev. 2015, 49, 82–89. [Google Scholar] [CrossRef]
- Kobayashi, T.; Matsuyama, T.; Takeuchi, M.; Ito, S. Autism spectrum disorder and prenatal exposure to selective serotonin reuptake inhibitors: A systematic review and meta-analysis. Reprod. Toxicol. 2016, 65, 170–178. [Google Scholar] [CrossRef]
- Hranilovic, D.; Blazevic, S. Hyperserotonemia in Autism: 5HT-Regulating Proteins. In Comprehensive Guide to Autism; Patel, V.B., Preedy, V.R., Martin, C.R., Eds.; Springer: New York, NY, USA, 2014; pp. 717–739. [Google Scholar] [CrossRef]
- Ramamoorthy, S.; Bauman, A.L.; Moore, K.R.; Han, H.; Yang-Feng, T.; Chang, A.S.; Ganapathy, V.; Blakely, R.D. Antidepressant- and cocaine-sensitive human serotonin transporter: Molecular cloning, expression, and chromosomal localization. Proc. Natl. Acad. Sci. USA 1993, 90, 2542–2546. [Google Scholar] [CrossRef]
- Rosenfeld, C.S. The placenta-brain-axis. J. Neurosci. Res. 2021, 99, 271–283. [Google Scholar] [CrossRef]
- Balkovetz, D.F.; Tiruppathi, C.; Leibach, F.H.; Mahesh, V.B.; Ganapathy, V. Evidence for an imipramine-sensitive serotonin transporter in human placental brush-border membranes. J. Biol. Chem. 1989, 264, 2195–2198. [Google Scholar] [CrossRef]
- Fraser-Spears, R.; Krause-Heuer, A.M.; Basiouny, M.; Mayer, F.P.; Manishimwe, R.; Wyatt, N.A.; Dobrowolski, J.C.; Roberts, M.P.; Greguric, I.; Kumar, N.; et al. Comparative analysis of novel decynium-22 analogs to inhibit transport by the low-affinity, high-capacity monoamine transporters, organic cation transporters 2 and 3, and plasma membrane monoamine transporter. Eur. J. Pharmacol. 2019, 842, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Jansson, T.; Illsley, N.P. Osmotic water permeabilities of human placental microvillous and basal membranes. J. Membr. Biol. 1993, 132, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, H.; Moss, T.J.; Gatford, K.L.; Moritz, K.M.; Akison, L.; Fullston, T.; Hryciw, D.H.; Maloney, C.A.; Morris, M.J.; Wooldridge, A.L.; et al. A review of fundamental principles for animal models of DOHaD research: An Australian perspective. J. Dev. Orig. Health Dis. 2016, 7, 449–472. [Google Scholar] [CrossRef]
- Gobinath, A.R.; Workman, J.L.; Chow, C.; Lieblich, S.E.; Galea, L.A.M. Sex-dependent effects of maternal corticosterone and SSRI treatment on hippocampal neurogenesis across development. Biol. Sex Differ. 2017, 8, 20. [Google Scholar] [CrossRef]
- Gobinath, A.R.; Workman, J.L.; Chow, C.; Lieblich, S.E.; Galea, L.A. Maternal postpartum corticosterone and fluoxetine differentially affect adult male and female offspring on anxiety-like behavior, stress reactivity, and hippocampal neurogenesis. Neuropharmacology 2016, 101, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Audus, K.L. Controlling drug delivery across the placenta. Eur. J. Pharm. Sci. 1999, 8, 161–165. [Google Scholar] [CrossRef][Green Version]
- Uhr, M.; Tontsch, A.; Namendorf, C.; Ripke, S.; Lucae, S.; Ising, M.; Dose, T.; Ebinger, M.; Rosenhagen, M.; Kohli, M.; et al. Polymorphisms in the Drug Transporter Gene ABCB1 Predict Antidepressant Treatment Response in Depression. Neuron 2008, 57, 203–209. [Google Scholar] [CrossRef]
- Feng, B.; Mills, J.B.; Davidson, R.E.; Mireles, R.J.; Janiszewski, J.S.; Troutman, M.D.; de Morais, S.M. In vitro P-glycoprotein assays to predict the in vivo interactions of P-glycoprotein with drugs in the central nervous system. Drug Metab. Dispos. 2008, 36, 268–275. [Google Scholar] [CrossRef]
- Wichman, C.L.; Stern, T.A. Diagnosing and Treating Depression During Pregnancy. Prim. Care Companion CNS Disord. 2015, 17, 10.4088/PCC.4015f01776. [Google Scholar] [CrossRef]
- Thormahlen, G.M. Paroxetine use during pregnancy: Is it safe? Ann. Pharm. 2006, 40, 1834–1837. [Google Scholar] [CrossRef]
- Horton, R.E. Decynium-22 enhances SSRI-induced antidepressant-like effects in mice: Uncovering novel targets to treat depression. J. Neurosci. 2013, 33, 10534–10543. [Google Scholar] [CrossRef]
- Margraf, A. Maturation of Platelet Function During Murine Fetal Development In Vivo. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1076–1086. [Google Scholar] [CrossRef] [PubMed]
- Mercado, C.P.; Kilic, F. Molecular mechanisms of SERT in platelets: Regulation of plasma serotonin levels. Mol. Interv. 2010, 10, 231–241. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antidepressant | nM | F/M Ratio | |||||
|---|---|---|---|---|---|---|---|
| IC50 | IC50 | Ca | Cmax | Ca;fetal | |||
| MVM | BM | ||||||
| Stronger inhibitor | Paroxetine | 10.09 | 4.39 | 455 | 1000 | 209 | 0.46 |
| Citalopram | 16.52 | 15.07 | 216 | 1200 | 147 | 0.68 | |
| Fluvoxamine | 19.95 | 15.21 | 446 | 1300 | 348 | 0.78 | |
| Weaker inhibitor | Fluoxetine | 29.99 | 25.18 | 557 | 1900 | 368 | 0.66 |
| Sertraline | 25.18 | 34.43 | 104 | 1300 | 53 | 0.51 | |
| Venlafaxine | 32.07 | 30.76 | 225 | 1400 | 162 | 0.72 | |
| Efflux Transporter | Citalopram | Paroxetine | Sertraline |
|---|---|---|---|
| MDCKII-P-gp | 2.75 ± 0.79 | 1.39 ± 0.30 | 1.17 ± 0.20 |
| MDCKII-BCRP | 1.18 ± 0.32 | 0.92 ± 0.02 | 1.11 ± 0.61 |
| MDCKII-MRP2 | 0.61 ± 0.11 | - | 1.69 ± 0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horackova, H.; Karahoda, R.; Cerveny, L.; Vachalova, V.; Ebner, R.; Abad, C.; Staud, F. Effect of Selected Antidepressants on Placental Homeostasis of Serotonin: Maternal and Fetal Perspectives. Pharmaceutics 2021, 13, 1306. https://doi.org/10.3390/pharmaceutics13081306
Horackova H, Karahoda R, Cerveny L, Vachalova V, Ebner R, Abad C, Staud F. Effect of Selected Antidepressants on Placental Homeostasis of Serotonin: Maternal and Fetal Perspectives. Pharmaceutics. 2021; 13(8):1306. https://doi.org/10.3390/pharmaceutics13081306
Chicago/Turabian StyleHorackova, Hana, Rona Karahoda, Lukas Cerveny, Veronika Vachalova, Ronja Ebner, Cilia Abad, and Frantisek Staud. 2021. "Effect of Selected Antidepressants on Placental Homeostasis of Serotonin: Maternal and Fetal Perspectives" Pharmaceutics 13, no. 8: 1306. https://doi.org/10.3390/pharmaceutics13081306
APA StyleHorackova, H., Karahoda, R., Cerveny, L., Vachalova, V., Ebner, R., Abad, C., & Staud, F. (2021). Effect of Selected Antidepressants on Placental Homeostasis of Serotonin: Maternal and Fetal Perspectives. Pharmaceutics, 13(8), 1306. https://doi.org/10.3390/pharmaceutics13081306