
Improving Membrane Activity and Cargo Delivery Efficacy of a Cell-Penetrating Peptide by Loading with Carboranes



Abstract
:1. Introduction
2. Materials and Methods
2.1. Carborane Synthesis
2.2. Peptide Synthesis
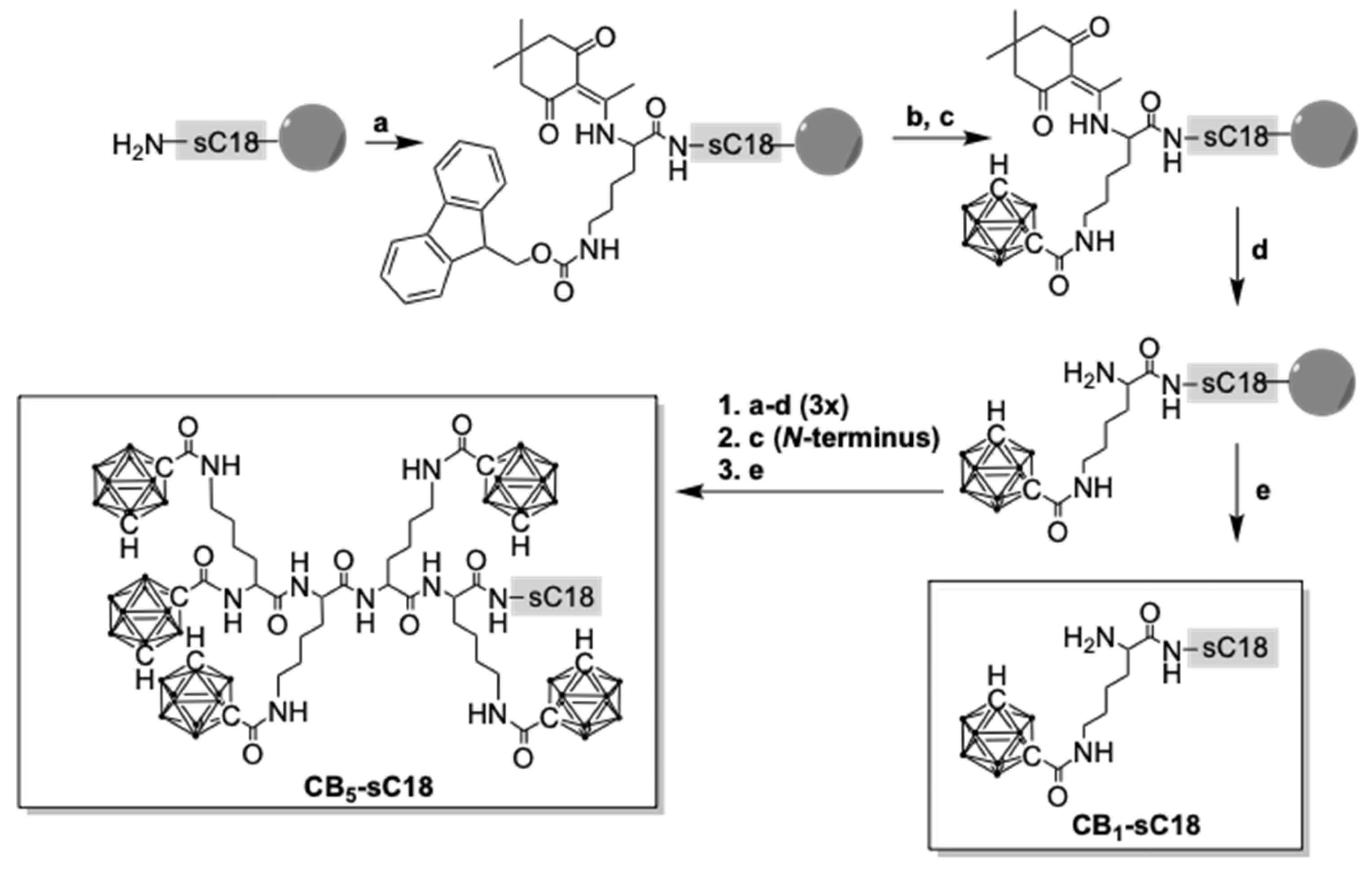
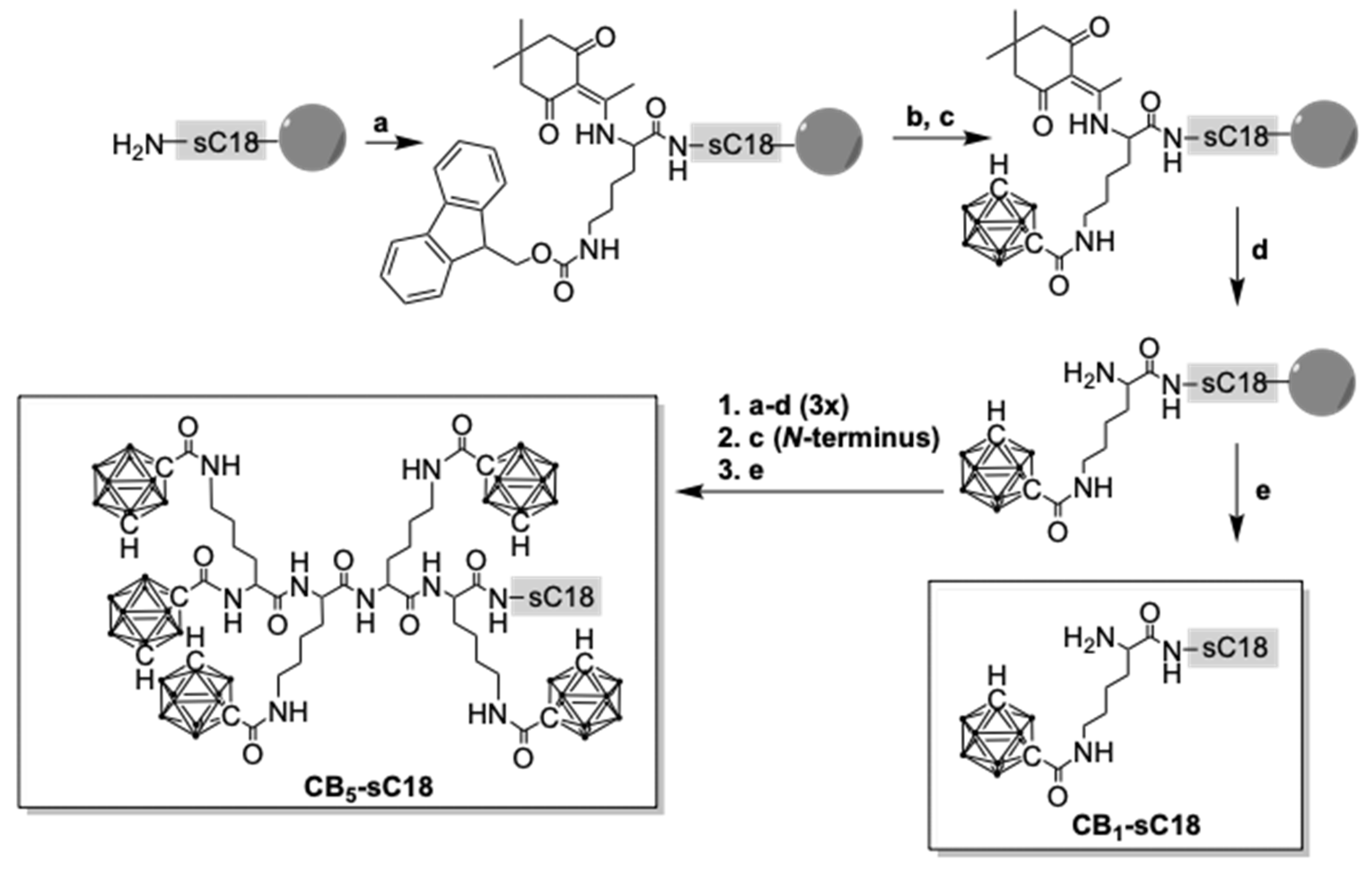
2.3. Carboranyl-Peptide Synthesis
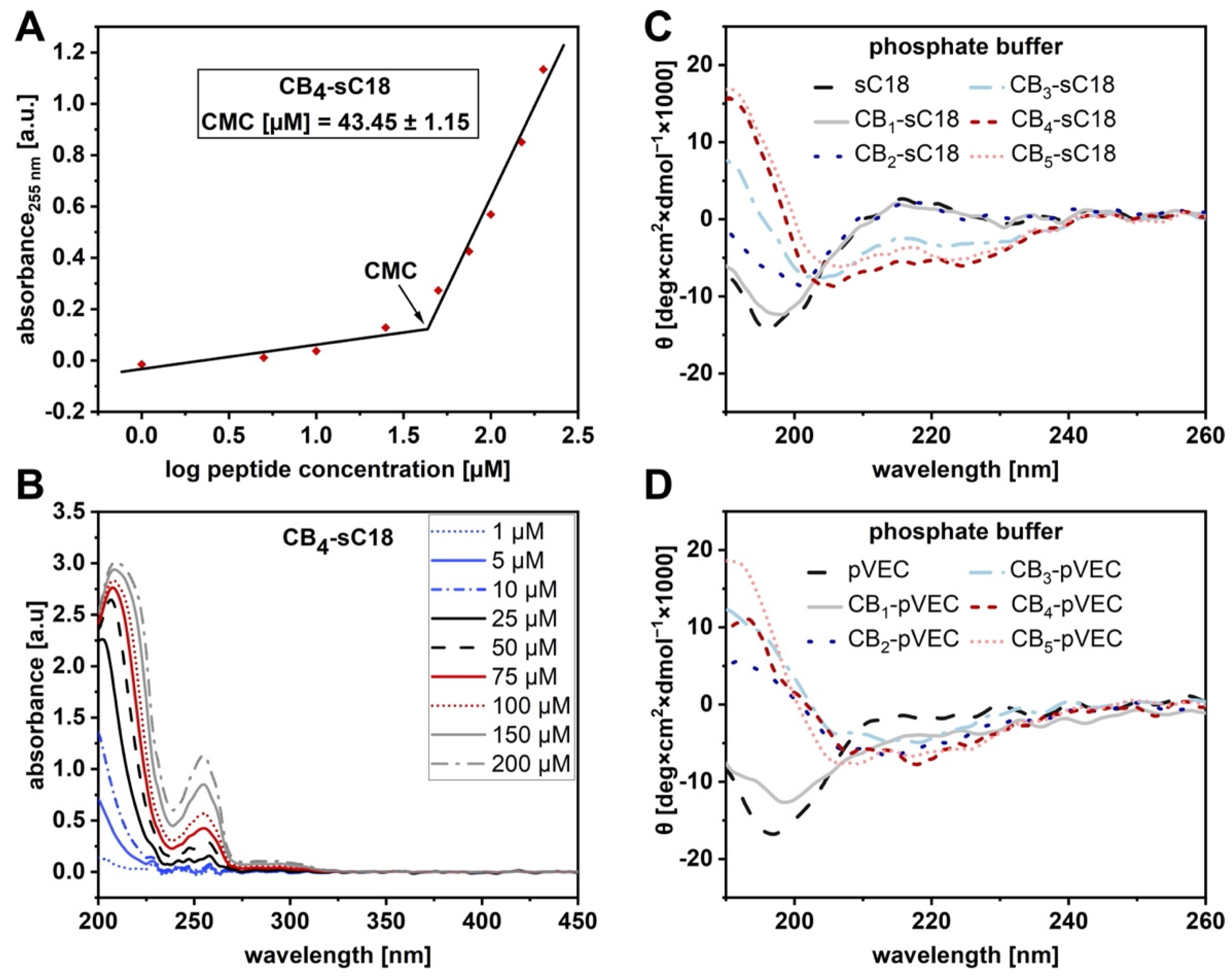
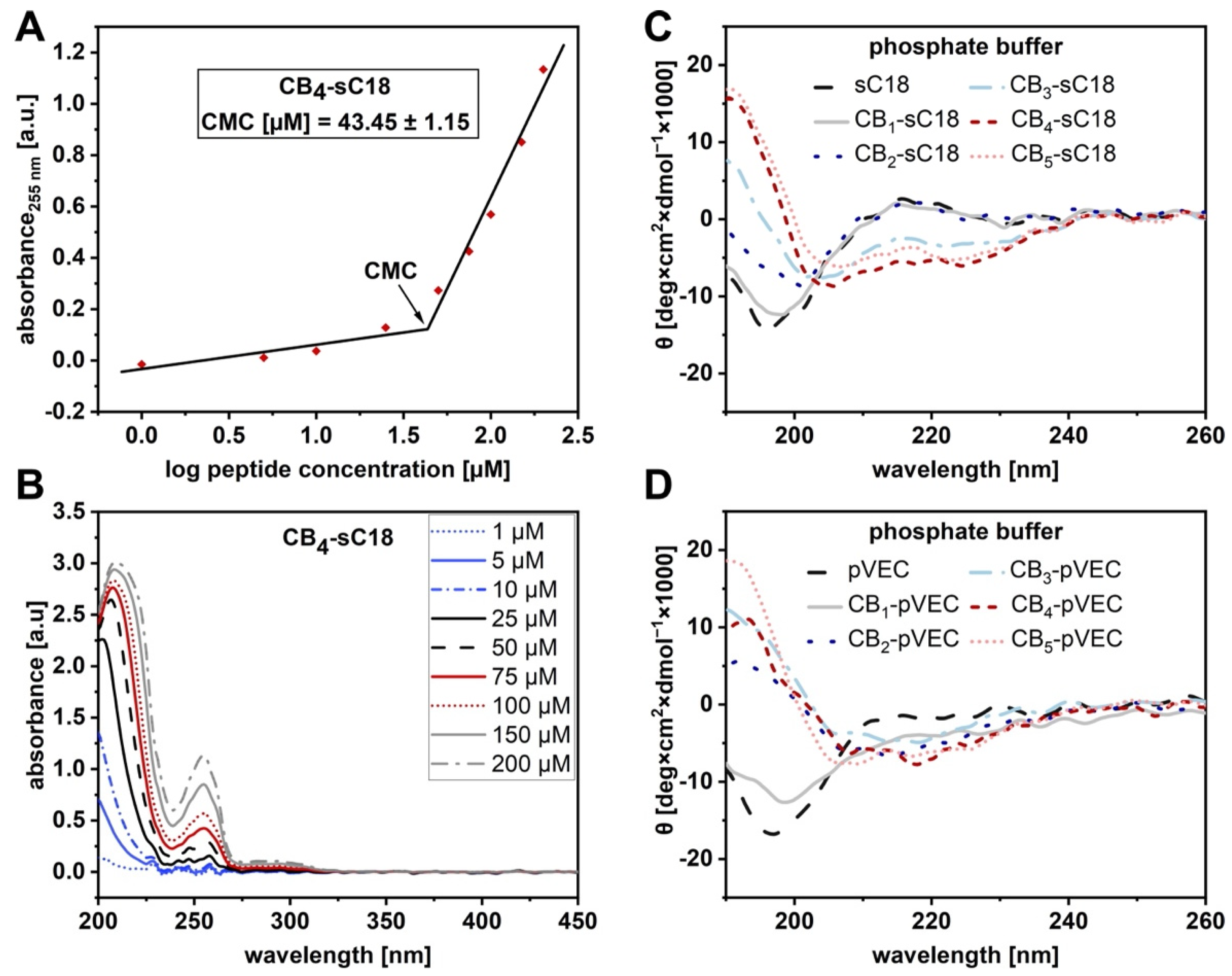
2.4. Circular Dichroism Spectroscopy
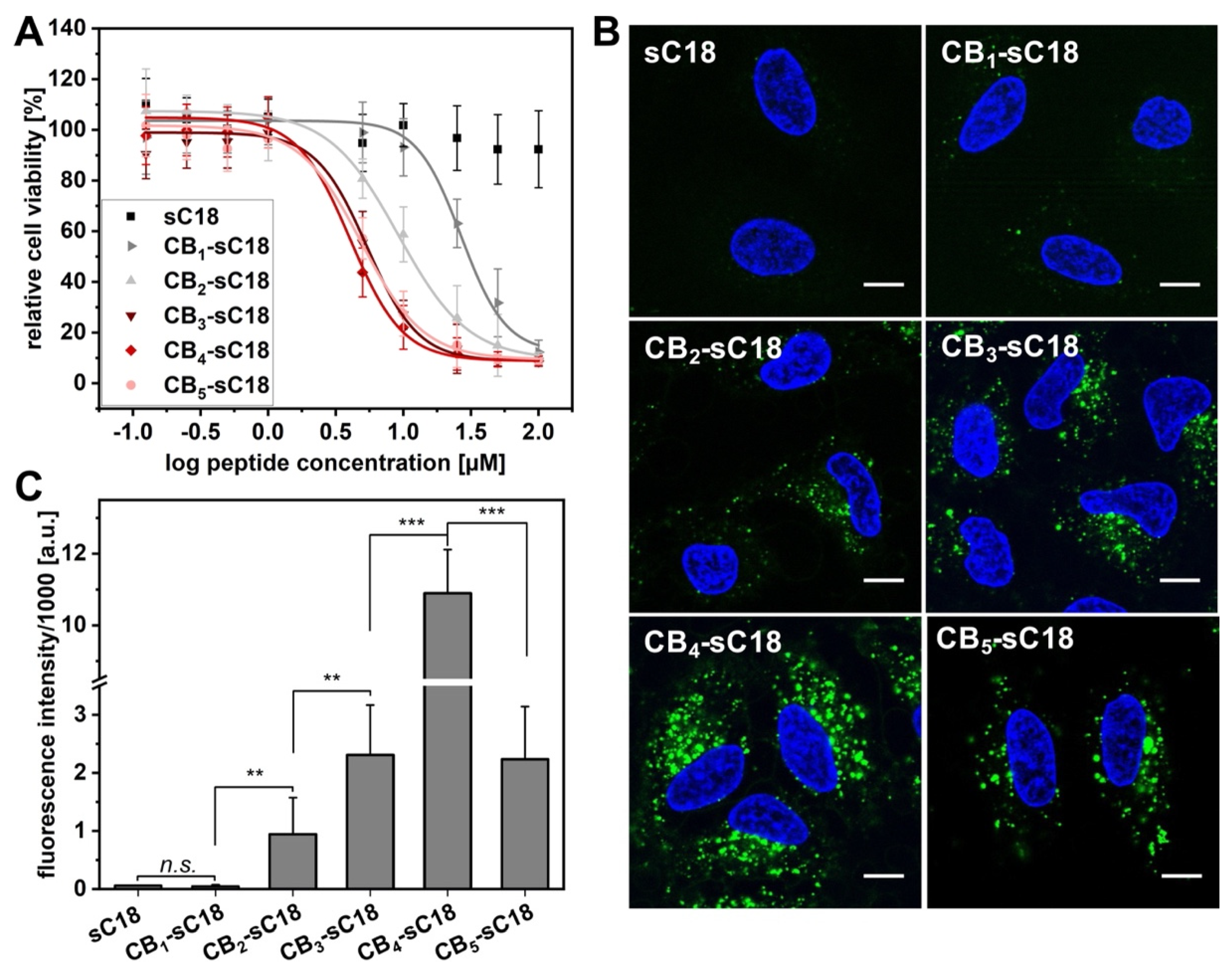
2.5. Cell Viability Assay
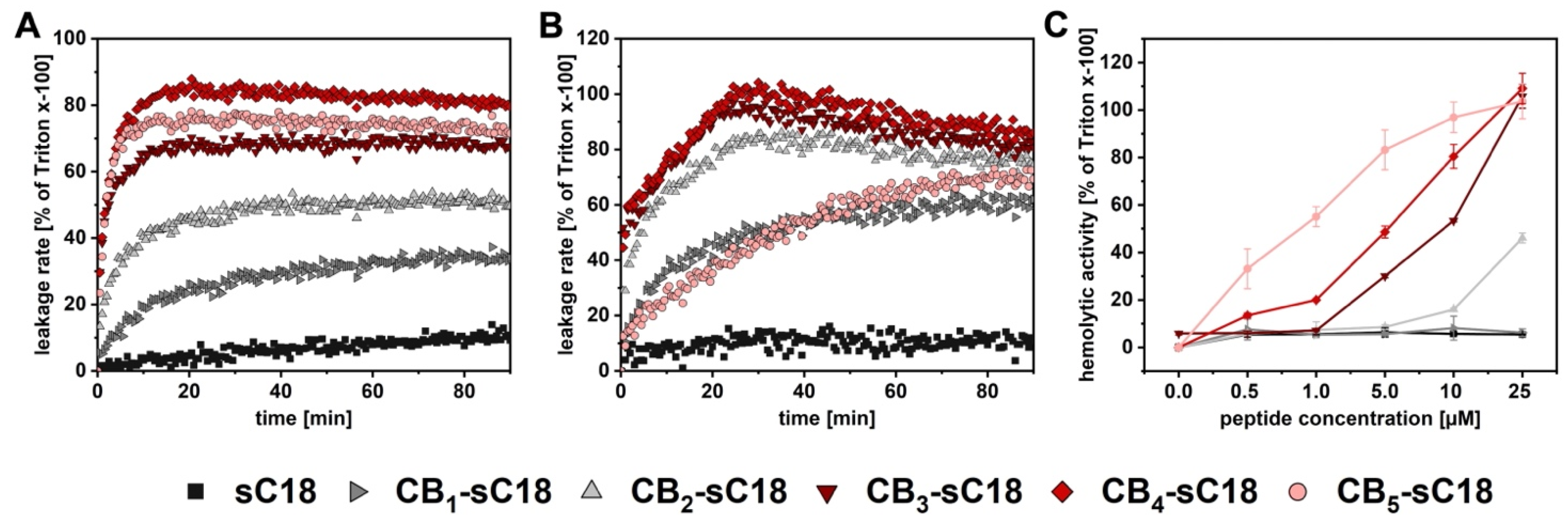
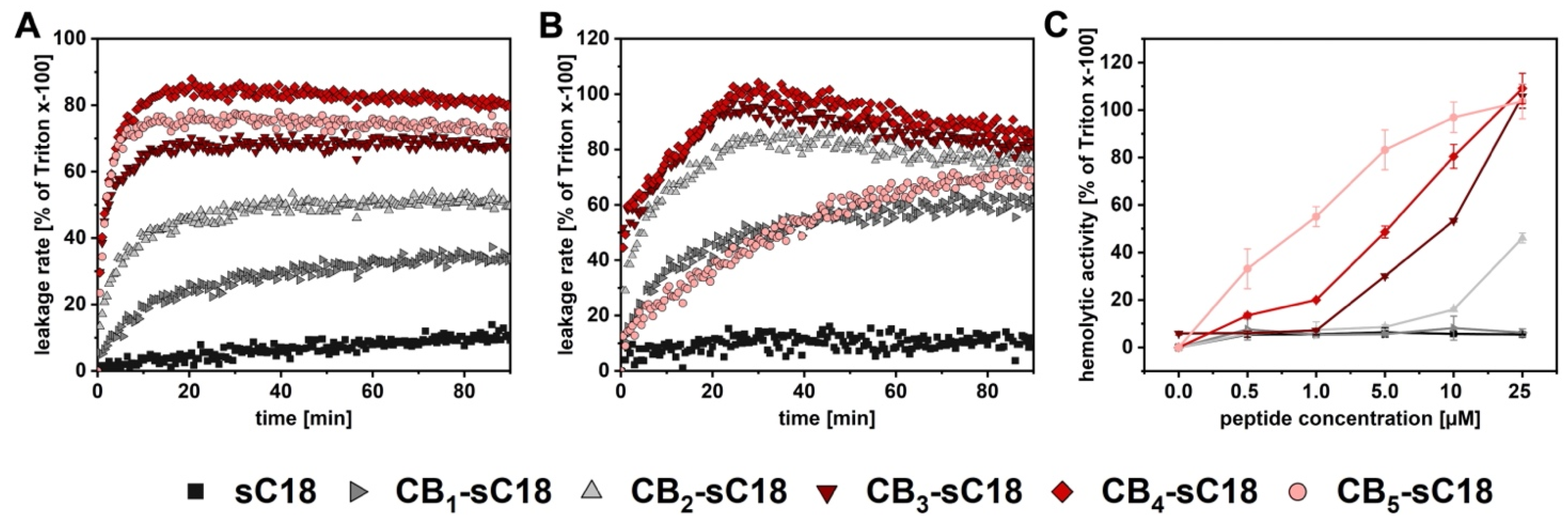
2.6. CF-Leakage Assay
2.7. Hemolysis Assay
2.8. Flow Cytometry
2.9. Confocal Laser Scanning Microscopy (CLSM)
2.10. Critical Micelle Concentration (CMC) Determination
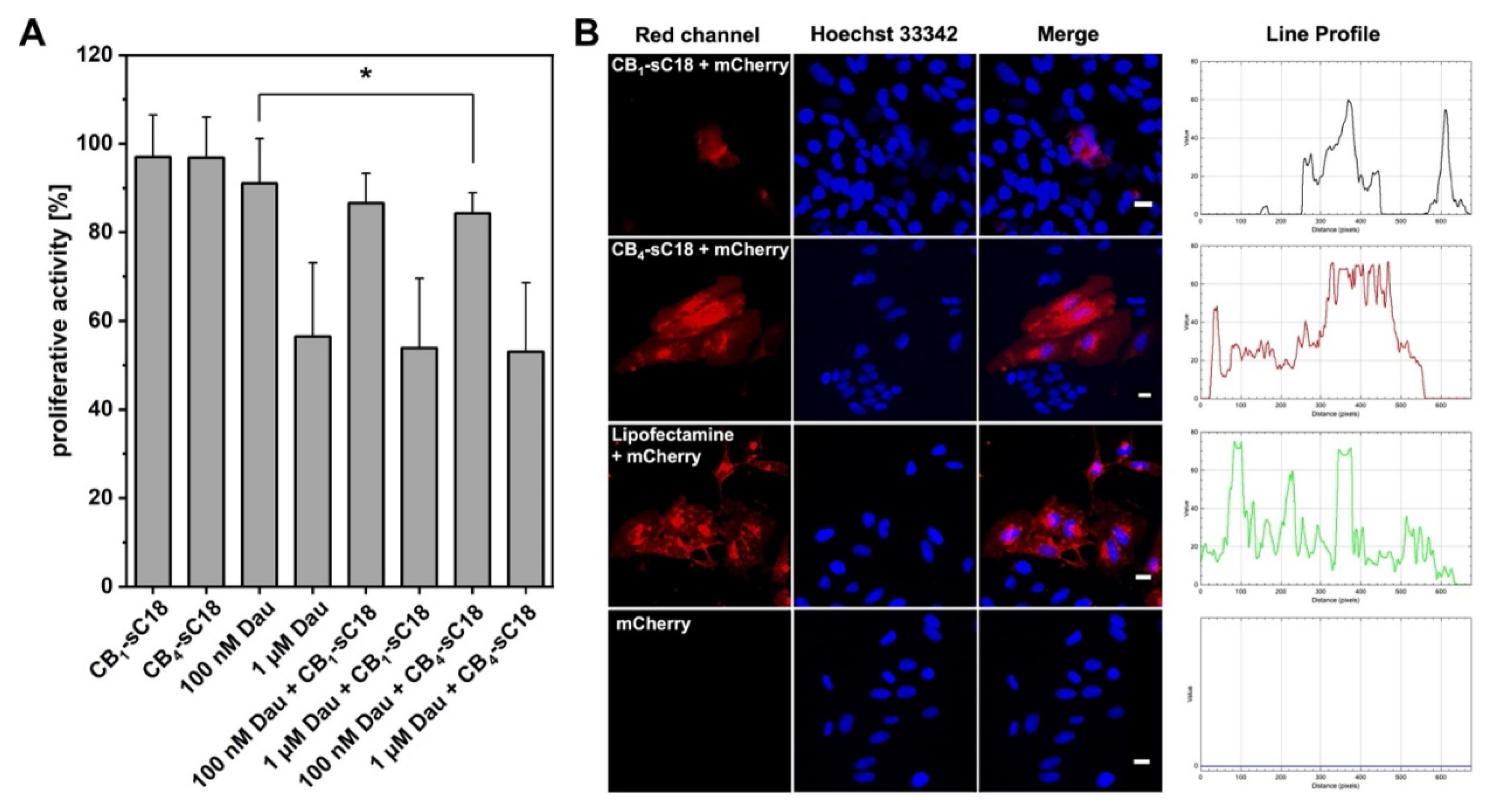
2.11. Proliferation Assay
2.12. Electrophoretic Mobility Shift Assay (EMSA)
2.13. Transfection with mCherry Plasmid
3. Results
3.1. Synthesis of Carborane-Peptide Conjugates
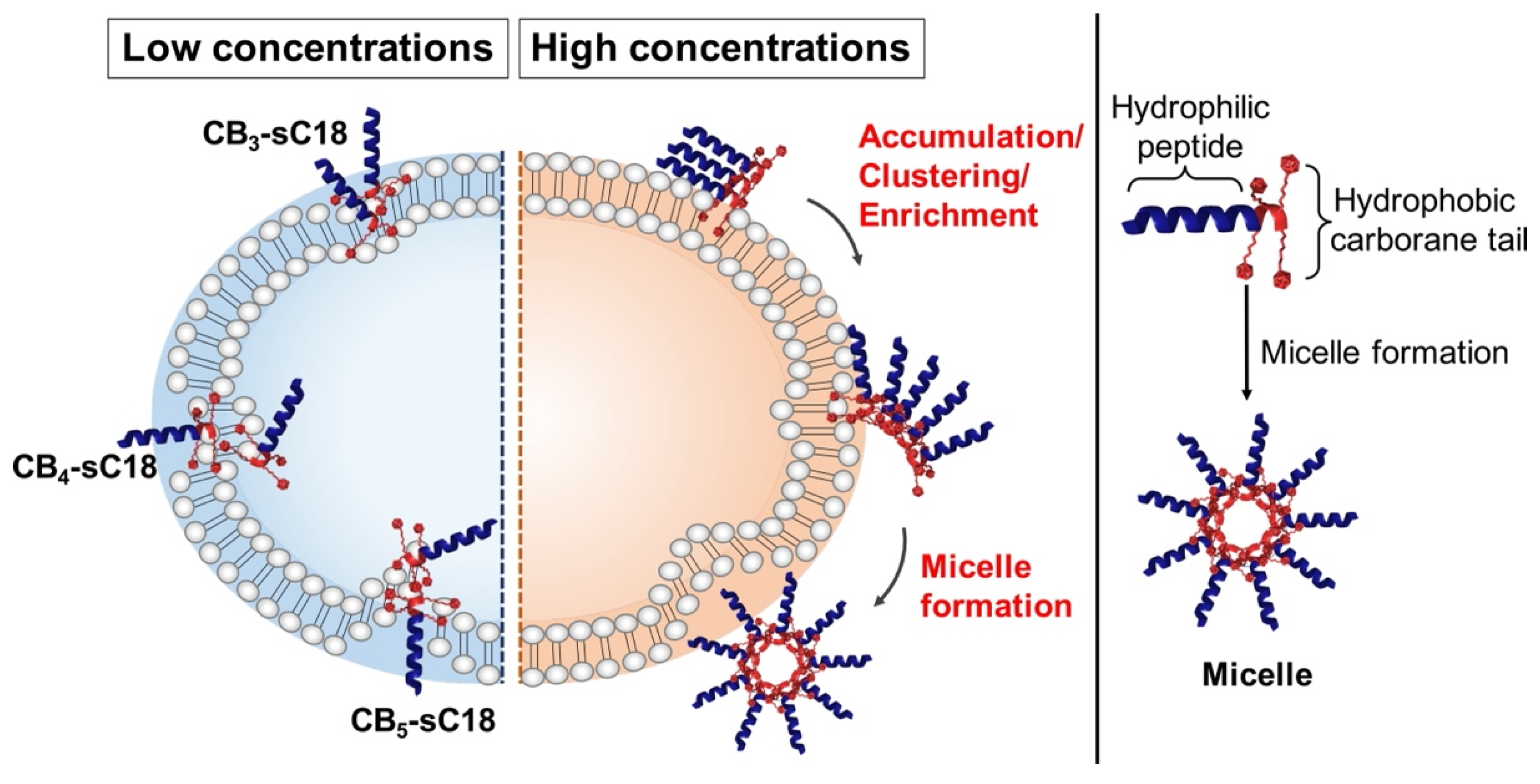
3.2. Physicochemical Properties of CB-CPP Conjugates
3.3. Interaction with Artificial and Biological Membranes
3.4. Cytotoxicity and Cellular Uptake Studies
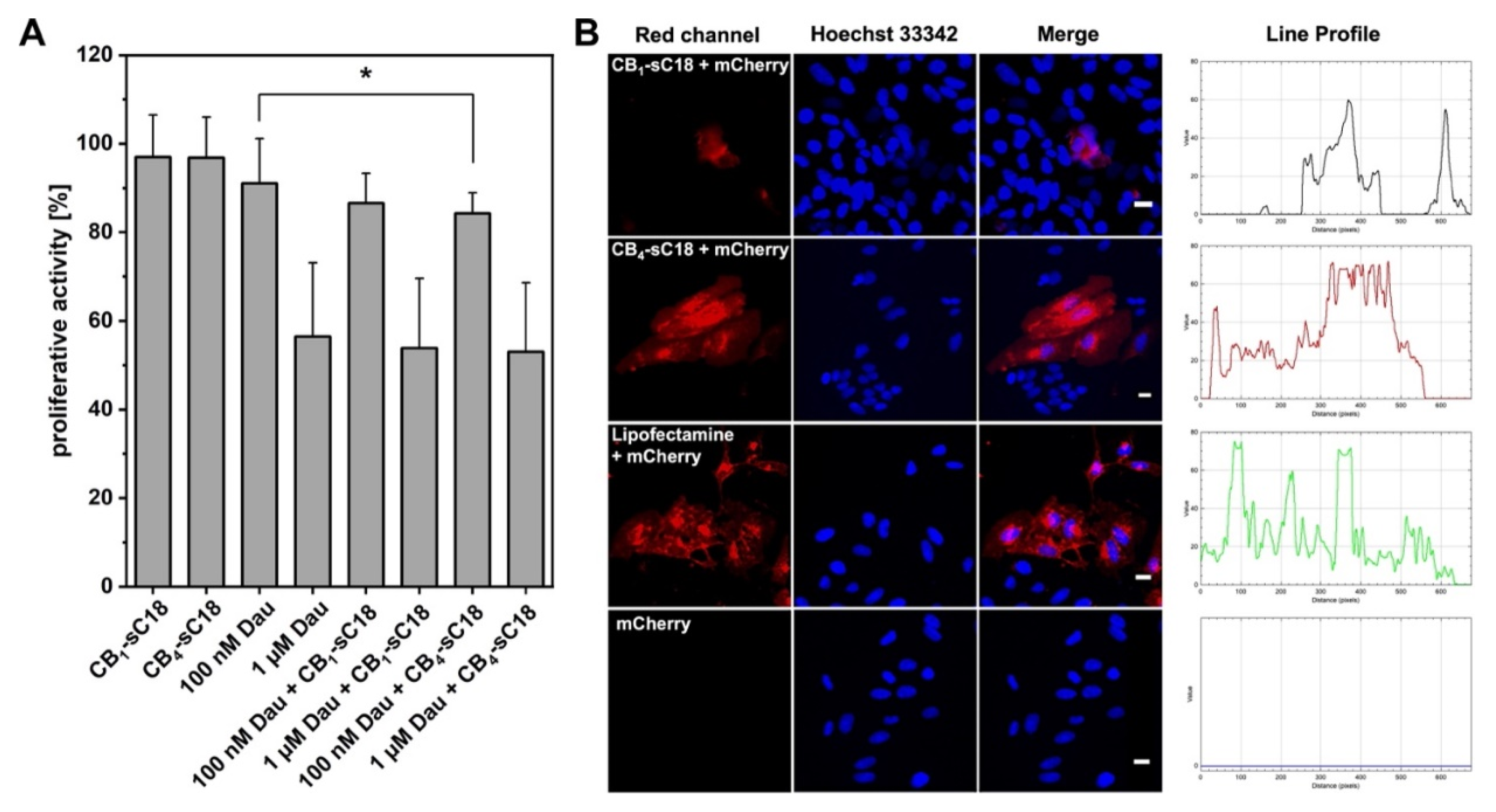
3.5. Cargo Delivery Studies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wagstaff, K.; Jans, D. Protein Transduction: Cell Penetrating Peptides and Their Therapeutic Applications. Curr. Med. Chem. 2006, 13, 1371–1387. [Google Scholar] [CrossRef] [PubMed]
- Di Pisa, M.; Chassaing, G.; Swiecicki, J.-M. Translocation Mechanism(s) of Cell-Penetrating Peptides: Biophysical Studies Using Artificial Membrane Bilayers. Biochemistry 2015, 54, 194–207. [Google Scholar] [CrossRef]
- Kardani, K.; Milani, A.; Shabani, S.H.; Bolhassani, A. Cell Penetrating Peptides: The Potent Multi-Cargo Intracellular Carriers. Expert Opin. Drug Deliv. 2019, 16, 1227–1258. [Google Scholar] [CrossRef]
- Yamashita, H.; Demizu, Y.; Shoda, T.; Sato, Y.; Oba, M.; Tanaka, M.; Kurihara, M. Amphipathic Short Helix-Stabilized Peptides with Cell-Membrane Penetrating Ability. Bioorg. Med. Chem. 2014, 22, 2403–2408. [Google Scholar] [CrossRef]
- Copolovici, D.M.; Langel, K.; Eriste, E.; Langel, Ü. Cell-Penetrating Peptides: Design, Synthesis, and Applications. ACS Nano 2014, 8, 1972–1994. [Google Scholar] [CrossRef]
- Aroui, S.; Brahim, S.; De Waard, M.; Bréard, J.; Kenani, A. Efficient Induction of Apoptosis by Doxorubicin Coupled to Cell-Penetrating Peptides Compared to Unconjugated Doxorubicin in the Human Breast Cancer Cell Line MDA-MB 231. Cancer Lett. 2009, 285, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Gessner, I.; Neundorf, I. Nanoparticles Modified with Cell-Penetrating Peptides: Conjugation Mechanisms, Physicochemical Properties, and Application in Cancer Diagnosis and Therapy. Int. J. Mol. Sci. 2020, 21, 2536. [Google Scholar] [CrossRef] [Green Version]
- de Figueiredo, I.R.; Freire, J.M.; Flores, L.; Veiga, A.S.; Castanho, M.A.R.B. Cell-Penetrating Peptides: A Tool for Effective Delivery in Gene-Targeted Therapies. IUBMB Life 2014, 66, 182–194. [Google Scholar] [CrossRef]
- Künnapuu, K.; Veiman, K.-L.; Porosk, L.; Rammul, E.; Kiisholts, K.; Langel, Ü.; Kurrikoff, K. Tumor Gene Therapy by Systemic Delivery of Plasmid DNA with Cell-Penetrating Peptides. FASEB Bioadvances 2018, 1, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurrikoff, K.; Langel, Ü. Recent CPP-Based Applications in Medicine. Expert Opin. Drug Deliv. 2019, 16, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Sebbage, V. Cell-Penetrating Peptides and Their Therapeutic Applications. Biosci. Horiz. Int. J. Stud. Res. 2009, 2, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Klimpel, A.; Lützenburg, T.; Neundorf, I. Recent Advances of Anti-Cancer Therapies Including the Use of Cell-Penetrating Peptides. Curr. Opin. Pharmacol. 2019, 47, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Yin, L.; Kim, K.H.; Cheng, J. Helical Poly(Arginine) Mimics with Superior Cell-Penetrating and Molecular Transporting Properties. Chem. Sci. 2013, 4, 3839–3844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karle, I.L. Controls Exerted by the Aib Residue: Helix Formation and Helix Reversal. Pept. Sci. 2001, 60, 351–365. [Google Scholar] [CrossRef]
- Jobin, M.-L.; Blanchet, M.; Henry, S.; Chaignepain, S.; Manigand, C.; Castano, S.; Lecomte, S.; Burlina, F.; Sagan, S.; Alves, I.D. The Role of Tryptophans on the Cellular Uptake and Membrane Interaction of Arginine-Rich Cell Penetrating Peptides. Biochim. Biophys. Acta (BBA)-Biomembr. 2015, 1848, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Carneado, J.; Kogan, M.J.; Van Mau, N.; Pujals, S.; López-Iglesias, C.; Heitz, F.; Giralt, E. Fatty Acyl Moieties: Improving Pro-Rich Peptide Uptake inside HeLa Cells. J. Pept. Res. 2005, 65, 580–590. [Google Scholar] [CrossRef]
- Som, A.; Reuter, A.; Tew, G.N. Protein Transduction Domain Mimics: The Role of Aromatic Functionality. Angew. Chem. Int. Ed. 2012, 51, 980–983. [Google Scholar] [CrossRef] [PubMed]
- Carrigan, C.N.; Imperiali, B. The Engineering of Membrane-Permeable Peptides. Anal. Biochem. 2005, 341, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Scholz, M.S.; Wingen, L.M.; Brunst, S.; Wittmann, S.K.; Cardoso, I.L.A.; Weizel, L.; Proschak, E. Soluble Epoxide Hydrolase Inhibitors with Carboranes as Non-Natural 3-D Pharmacophores. Eur. J. Med. Chem. 2020, 185, 111766. [Google Scholar] [CrossRef] [PubMed]
- Worm, D.J.; Hoppenz, P.; Els-Heindl, S.; Kellert, M.; Kuhnert, R.; Saretz, S.; Köbberling, J.; Riedl, B.; Hey-Hawkins, E.; Beck-Sickinger, A.G. Selective Neuropeptide Y Conjugates with Maximized Carborane Loading as Promising Boron Delivery Agents for Boron Neutron Capture Therapy. J. Med. Chem. 2019, 63, 2358–2371. [Google Scholar] [CrossRef] [PubMed]
- Scholz, M.; Hey-Hawkins, E. Carbaboranes as Pharmacophores: Properties, Synthesis, and Application Strategies. Chem. Rev. 2011, 111, 7035–7062. [Google Scholar] [CrossRef]
- Hoppenz, P.; Els-Heindl, S.; Kellert, M.; Kuhnert, R.; Saretz, S.; Lerchen, H.-G.; Köbberling, J.; Riedl, B.; Hey-Hawkins, E.; Beck-Sickinger, A.G. A Selective Carborane-Functionalized Gastrin-Releasing Peptide Receptor Agonist as Boron Delivery Agent for Boron Neutron Capture Therapy. J. Org. Chem. 2020, 85, 1446–1457. [Google Scholar] [CrossRef]
- Michiue, H.; Sakurai, Y.; Kondo, N.; Kitamatsu, M.; Bin, F.; Nakajima, K.; Hirota, Y.; Kawabata, S.; Nishiki, T.I.; Ohmori, I.; et al. The Acceleration of Boron Neutron Capture Therapy Using Multi-Linked Mercaptoundecahydrododecaborate (BSH) Fused Cell-Penetrating Peptide. Biomaterials 2014, 35, 3396–3405. [Google Scholar] [CrossRef]
- Lützenburg, T.; Neundorf, I.; Scholz, M. Direct Carborane-Peptide Conjugates: Synthesis and Evaluation as Non-Natural Lipopeptide Mimetics. Chem. Phys. Lipids 2018, 213, 62–67. [Google Scholar] [CrossRef]
- Neundorf, I.; Rennert, R.; Hoyer, J.; Schramm, F.; Löbner, K.; Kitanovic, I.; Wölfl, S. Fusion of a Short HA2-Derived Peptide Sequence to Cell-Penetrating Peptides Improves Cytosolic Uptake, but Enhances Cytotoxic Activity. Pharmaceuticals 2009, 2, 49–65. [Google Scholar] [CrossRef] [Green Version]
- Negrete-Hurtado, A.; Overhoff, M.; Bera, S.; De Bruyckere, E.; Schätzmüller, K.; Kye, M.J.; Qin, C.; Lammers, M.; Kondylis, V.; Neundorf, I.; et al. Autophagy Lipidation Machinery Regulates Axonal Microtubule Dynamics but Is Dispensable for Survival of Mammalian Neurons. Nat. Commun. 2020, 11, 1535. [Google Scholar] [CrossRef] [Green Version]
- Feni, L.; Parente, S.; Robert, C.; Gazzola, S.; Arosio, D.; Piarulli, U.; Neundorf, I. Kiss and Run: Promoting Effective and Targeted Cellular Uptake of a Drug Delivery Vehicle Composed of an Integrin-Targeting Diketopiperazine Peptidomimetic and a Cell-Penetrating Peptide. Bioconjugate Chem. 2019, 30, 2011–2022. [Google Scholar] [CrossRef] [Green Version]
- Klimpel, A.; Stillger, K.; Wiederstein, J.L.; Krüger, M.; Neundorf, I. Cell-Permeable CaaX-Peptides Affect K-Ras Downstream Signaling and Promote Cell Death in Cancer Cells. FEBS J. 2021, 288, 2911–2929. [Google Scholar] [CrossRef]
- Feni, L.; Jütten, L.; Parente, S.; Piarulli, U.; Neundorf, I.; Diaz, D. Cell-Penetrating Peptides Containing 2,5-Diketopiperazine (DKP) Scaffolds as Shuttles for Anti-Cancer Drugs: Conformational Studies and Biological Activity. Chem. Commun. 2020, 56, 5685–5688. [Google Scholar] [CrossRef]
- Ihsan, A.B.; Nargis, M.; Koyama, Y. Effects of the Hydrophilic-Lipophilic Balance of Alternating Peptides on Self-Assembly and Thermo-Responsive Behaviors. Int. J. Mol. Sci. 2019, 20, 4604. [Google Scholar] [CrossRef] [Green Version]
- Sparks, D.L.; Lund-Katz, S.; Phillips, M.C. The Charge and Structural Stability of Apolipoprotein A-I in Discoidal and Spherical Recombinant High Density Lipoprotein Particles. J. Biol. Chem. 1992, 267, 25839–25847. [Google Scholar] [CrossRef]
- Lakshminarayanan, R.; Fan, D.; Du, C.; Moradian-Oldak, J. The Role of Secondary Structure in the Entropically Driven Amelogenin Self-Assembly. Biophys. J. 2007, 93, 3664–3674. [Google Scholar] [CrossRef] [Green Version]
- Elmquist, A.; Lindgren, M.; Bartfai, T.; Langel, U. VE-Cadherin-Derived Cell-Penetrating Peptide, PVEC, with Carrier Functions. Exp. Cell Res. 2001, 269, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Gessner, I.; Klimpel, A.; Klußmann, M.; Neundorf, I.; Mathur, S. Interdependence of Charge and Secondary Structure on Cellular Uptake of Cell Penetrating Peptide Functionalized Silica Nanoparticles. Nanoscale Adv. 2020, 2, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Gronewold, A.; Horn, M.; Ranđelović, I.; Tóvári, J.; Muñoz Vázquez, S.; Schomäcker, K.; Neundorf, I. Characterization of a Cell-Penetrating Peptide with Potential Anticancer Activity. ChemMedChem 2017, 12, 42–49. [Google Scholar] [CrossRef]
- Hedegaard, S.F.; Derbas, M.S.; Lind, T.K.; Kasimova, M.R.; Christensen, M.V.; Michaelsen, M.H.; Campbell, R.A.; Jorgensen, L.; Franzyk, H.; Cárdenas, M.; et al. Fluorophore Labeling of a Cell-Penetrating Peptide Significantly Alters the Mode and Degree of Biomembrane Interaction. Sci. Rep. 2018, 8, 6327. [Google Scholar] [CrossRef] [PubMed]
- Birch, D.; Christensen, M.V.; Staerk, D.; Franzyk, H.; Nielsen, H.M. Fluorophore Labeling of a Cell-Penetrating Peptide Induces Differential Effects on Its Cellular Distribution and Affects Cell Viability. Biochim. Biophys. Acta (BBA)-Biomembr. 2017, 1859, 2483–2494. [Google Scholar] [CrossRef]
- Nair, P.; Dey, D.; Borkotoky, S.; Shukla, A.; Banerjee, M. Hydrophobicity and Oligomerization Are Essential Parameters for Membrane Penetration Activity of the VP4 Peptide from Hepatitis A Virus (HAV). Arch. Biochem. Biophys. 2019, 678, 108188. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.-A.; Separovic, F. How Membrane-Active Peptides Get into Lipid Membranes. Acc. Chem. Res. 2016, 49, 1130–1138. [Google Scholar] [CrossRef]
- Takayama, K.; Nakase, I.; Michiue, H.; Takeuchi, T.; Tomizawa, K.; Matsui, H.; Futaki, S. Enhanced Intracellular Delivery Using Arginine-Rich Peptides by the Addition of Penetration Accelerating Sequences (Pas). J. Control. Release 2009, 138, 128–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiríksdóttir, E.; Konate, K.; Langel, Ü.; Divita, G.; Deshayes, S. Secondary Structure of Cell-Penetrating Peptides Controls Membrane Interaction and Insertion. Biochim. Biophys. Acta (BBA)-Biomembr. 2010, 1798, 1119–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | MWcalc. [Da] | MWexp. [Da] |
|---|---|---|---|
| sC18 | 1GLRKRLRKFRNKIKEK16 | 2069.55 | 2070.34 |
| CB1-sC18 | K-GLRKRLRKFRNKIKEK | 2367.95 | 2368.74 |
| CB2-sC18 | KK-GLRKRLRKFRNKIKEK | 2666.37 | 2666.97 |
| CB3-sC18 | KKK-GLRKRLRKFRNKIKEK | 2965.79 | 2965.99 |
| CB4-sC18 | KKKK-GLRKRLRKFRNKIKEK | 3264.21 | 3263.82 |
| CB5-sC18 | CB-KKKK-GLRKRLRKFRNKIKEK | 3433.44 | 3434.29 |
| Peptide | Phosphate Buffer 1 | 50% TFE 1 | Phosphate Buffer 2 | 50% TFE 2 | CMC [µM] |
|---|---|---|---|---|---|
| sC18 | 3.45 | 71.22 | / | 0.89 | - |
| CB1-sC18 | 5.02 | 50.68 | / | 0.85 | - |
| CB2-sC18 | 5.45 | 60.49 | / | 0.89 | - |
| CB3-sC18 | 16.68 | 56.10 | 0.56 | 0.88 | 44.46 ± 1.14 |
| CB4-sC18 | 21.68 | 56.30 | 0.73 | 0.98 | 43.45 ± 1.15 |
| CB5-sC18 | 21.09 | 50.15 | 0.87 | 0.92 | 44.37 ± 1.16 |
| pVEC | 12.18 | 48.60 | 0.37 | 0.73 | - |
| CB1-pVEC | 17.72 | 44.28 | 0.57 | 0.82 | - |
| CB2-pVEC | 20.41 | 51.36 | 0.84 | 0.82 | - |
| CB3-pVEC | 16.98 | 65.76 | 0.99 | 0.82 | - |
| CB4-pVEC | 22.59 | 59.84 | 0.92 | 0.84 | - |
| CB5-pVEC | 22.96 | 45.48 | 0.79 | 0.79 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lützenburg, T.; Burdina, N.; Scholz, M.S.; Neundorf, I. Improving Membrane Activity and Cargo Delivery Efficacy of a Cell-Penetrating Peptide by Loading with Carboranes. Pharmaceutics 2021, 13, 2075. https://doi.org/10.3390/pharmaceutics13122075
Lützenburg T, Burdina N, Scholz MS, Neundorf I. Improving Membrane Activity and Cargo Delivery Efficacy of a Cell-Penetrating Peptide by Loading with Carboranes. Pharmaceutics. 2021; 13(12):2075. https://doi.org/10.3390/pharmaceutics13122075
Chicago/Turabian StyleLützenburg, Tamara, Nele Burdina, Matthias S. Scholz, and Ines Neundorf. 2021. "Improving Membrane Activity and Cargo Delivery Efficacy of a Cell-Penetrating Peptide by Loading with Carboranes" Pharmaceutics 13, no. 12: 2075. https://doi.org/10.3390/pharmaceutics13122075
APA StyleLützenburg, T., Burdina, N., Scholz, M. S., & Neundorf, I. (2021). Improving Membrane Activity and Cargo Delivery Efficacy of a Cell-Penetrating Peptide by Loading with Carboranes. Pharmaceutics, 13(12), 2075. https://doi.org/10.3390/pharmaceutics13122075