
Broad Spectrum Antiviral Properties of Cardiotonic Steroids Used as Potential Therapeutics for Emerging Coronavirus Infections

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Compounds
2.2. Cells and Viruses
2.3. Immunofluorescence Antiviral Assays
2.4. Viral Cytopathic Effect Assays
2.5. RNA Isolation and QuantSeq 3′ mRNA-Seq Analysis
2.6. Five-Day Repeated Dose Toxicity Study
2.7. Liver Microsomal Metabolic Stability Assays
2.8. hERG K+ Channel Binding Assays
2.9. Plasma Protein Binding Assays
2.10. Cytochrome P-450 (CYP450) Enzyme Inhibition Assays
2.11. Pharmacokinetic Studies
2.12. Statistical Analysis
3. Results
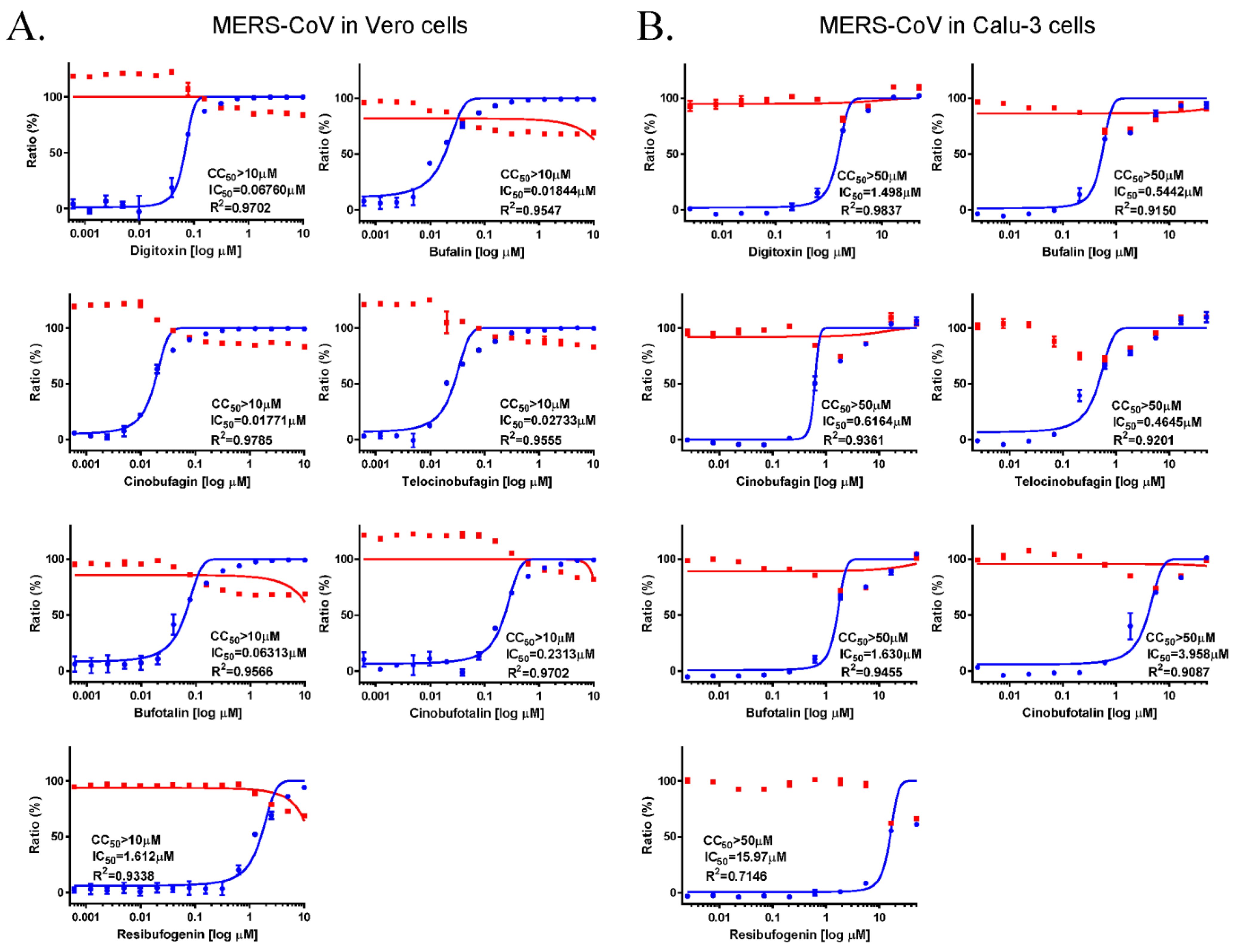
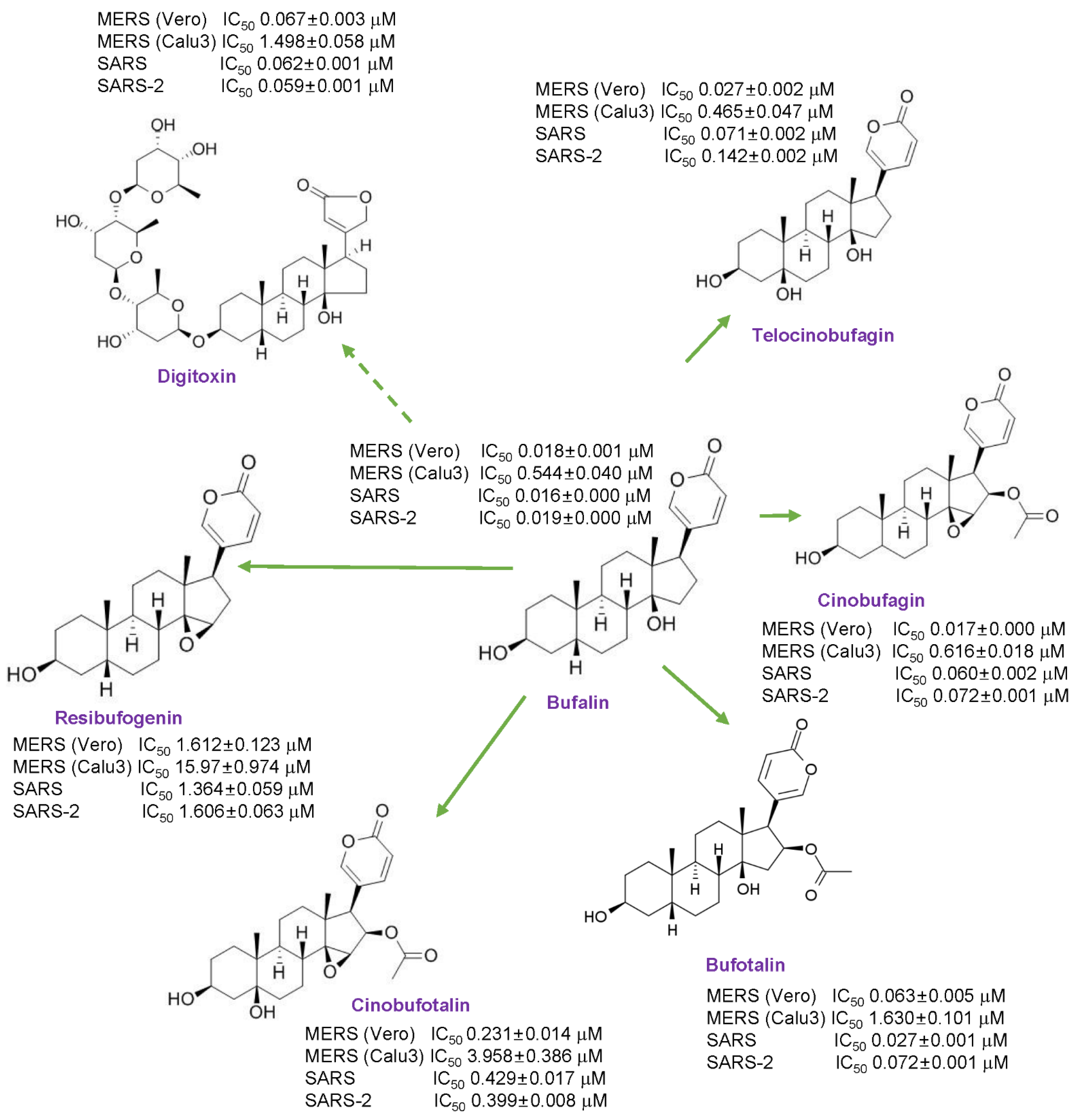
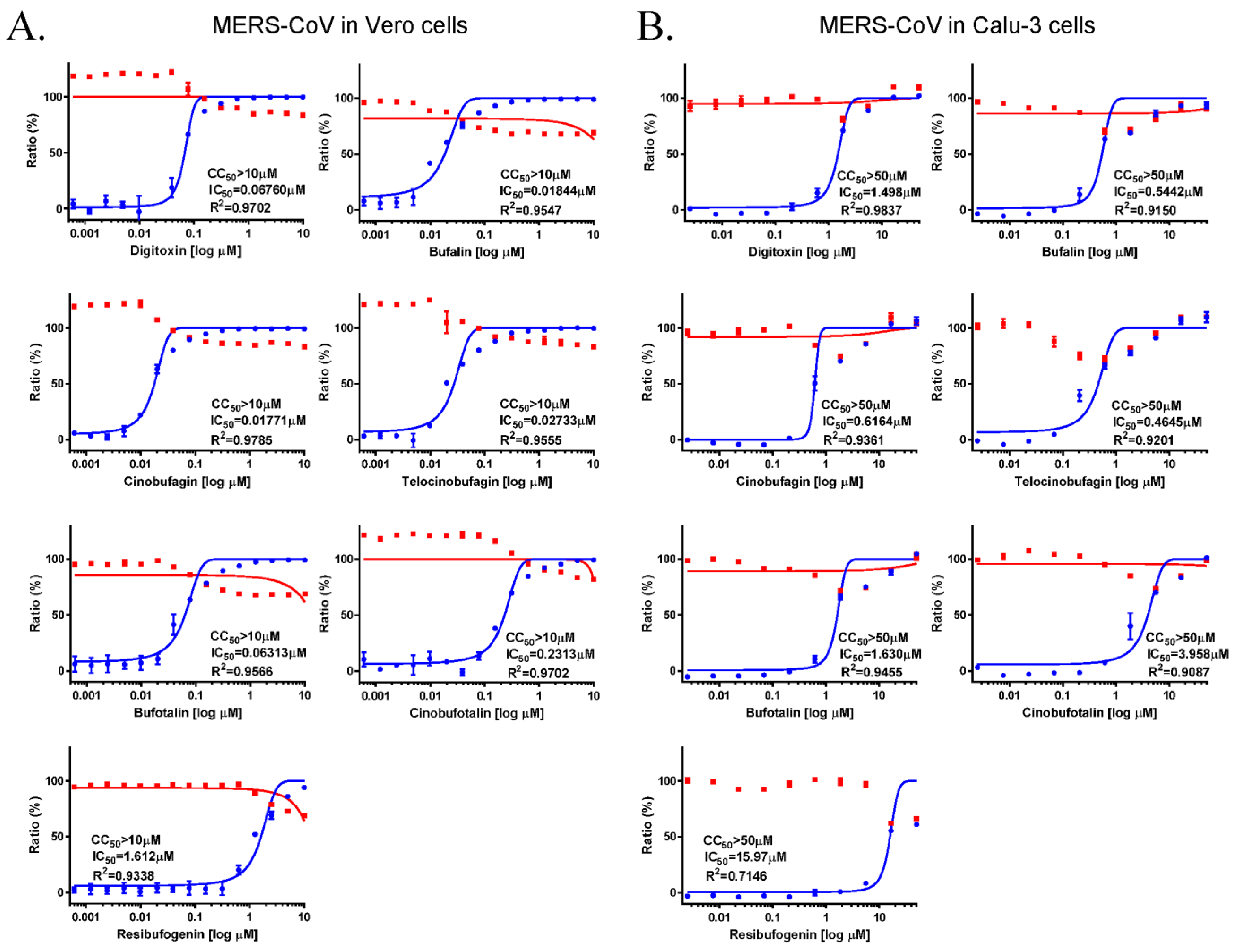
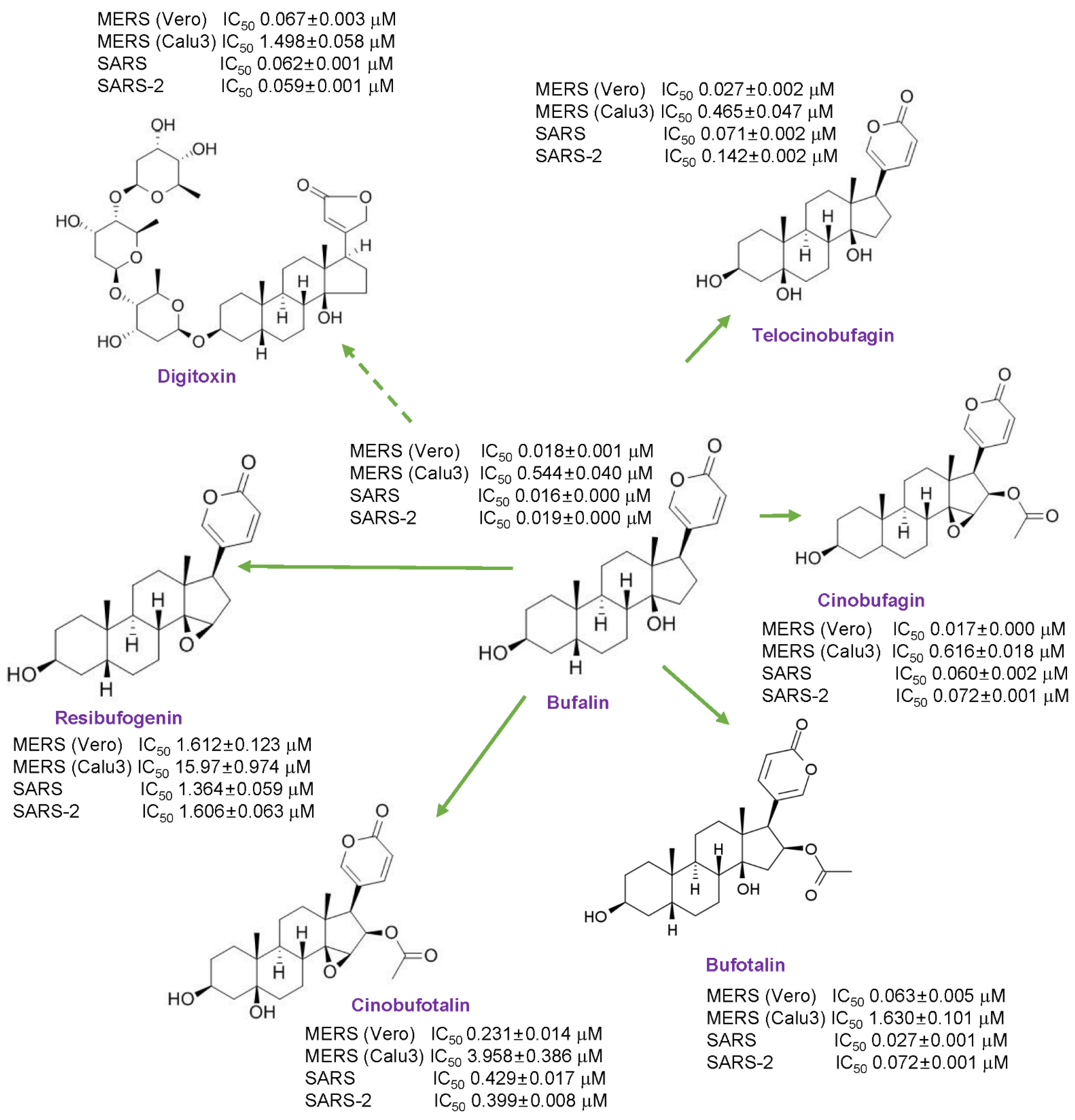
3.1. Anti-MERS-CoV Activity of Cardiotonic Steroids
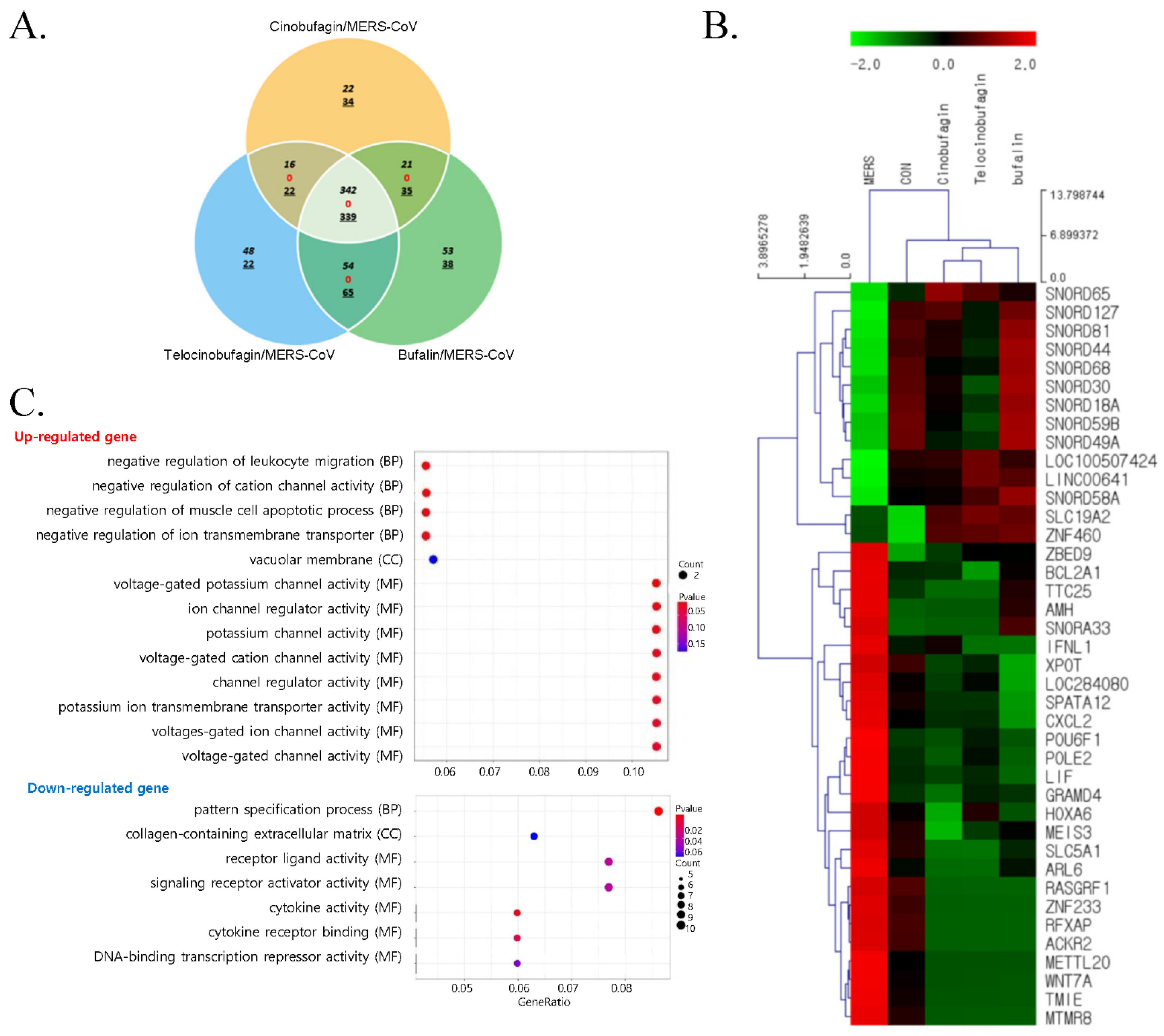
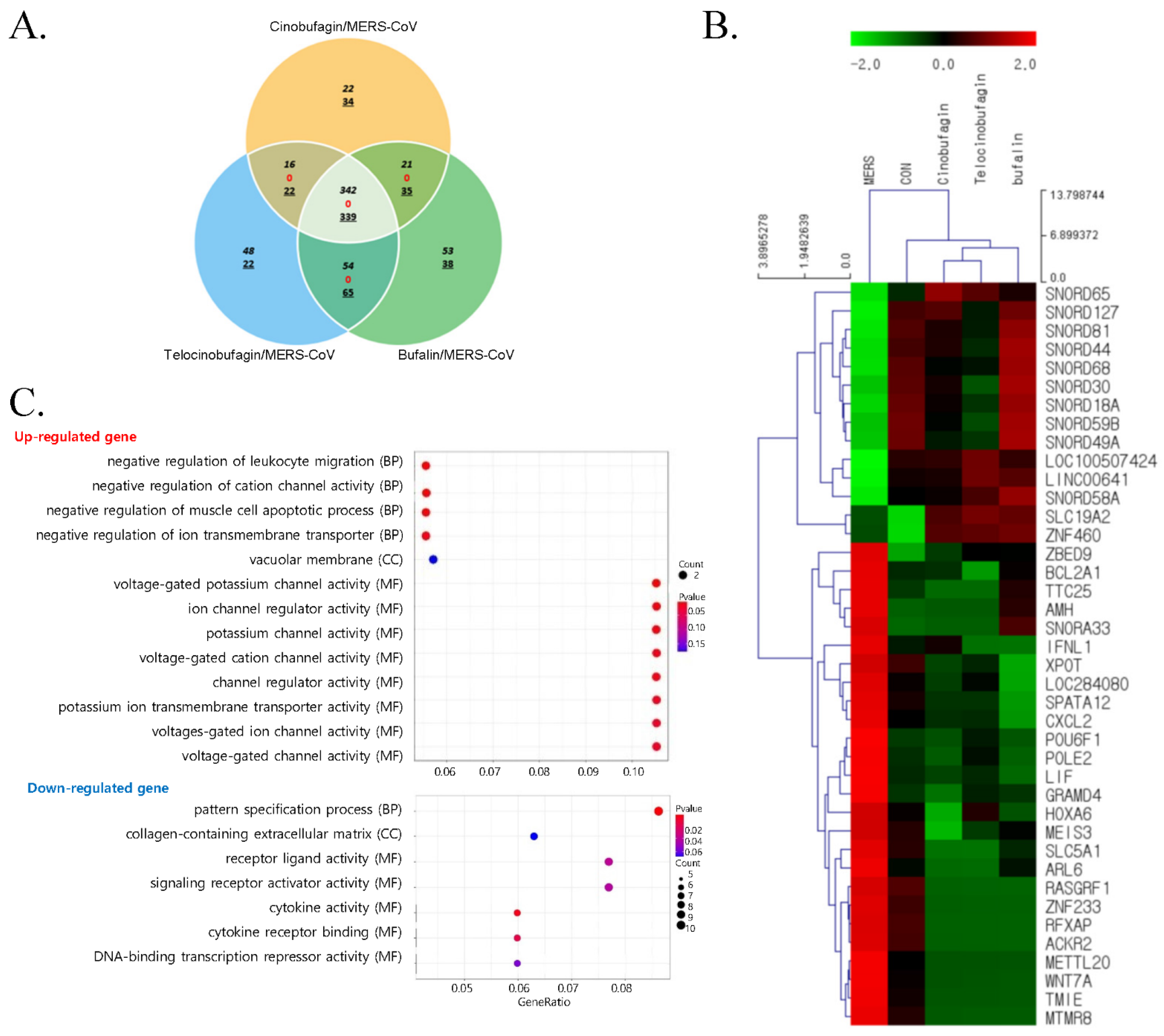
3.2. Comparative Gene Expression Analysis of Compound-Treated Calu-3 Cells during MERS-CoV Infection
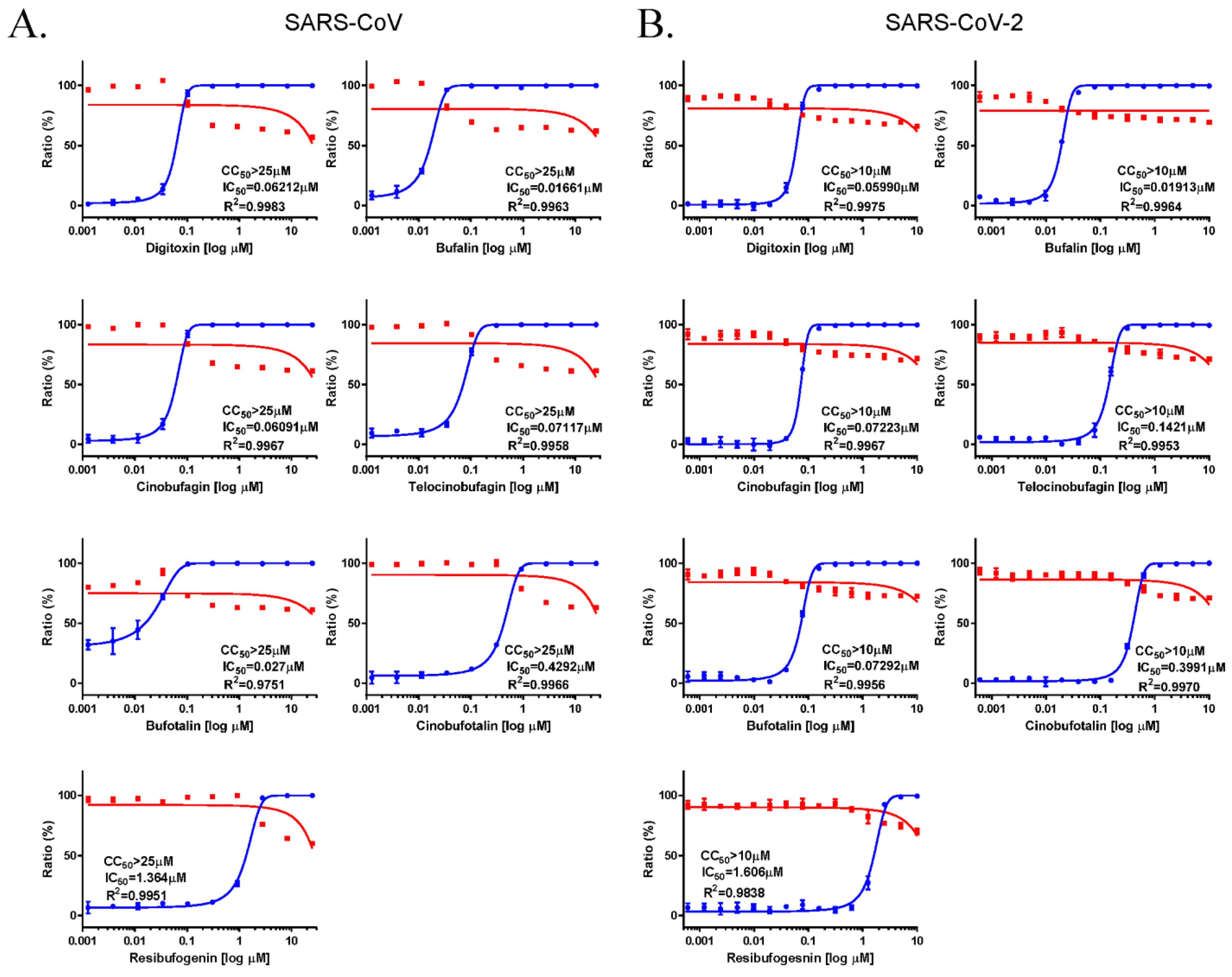
3.3. Anti-SARS-CoV and SARS-CoV-2 Activity of Cardiotonic Steroids
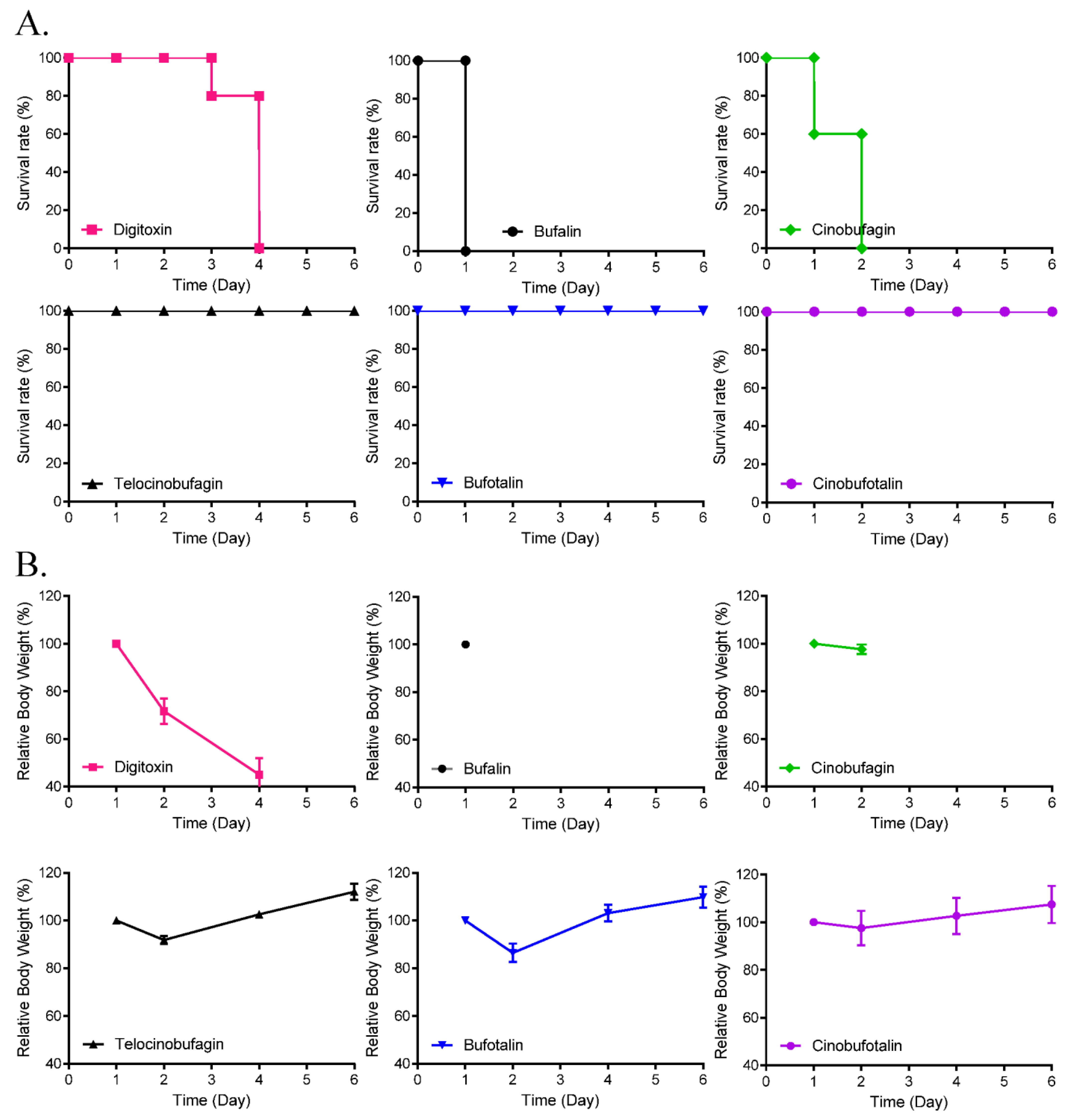
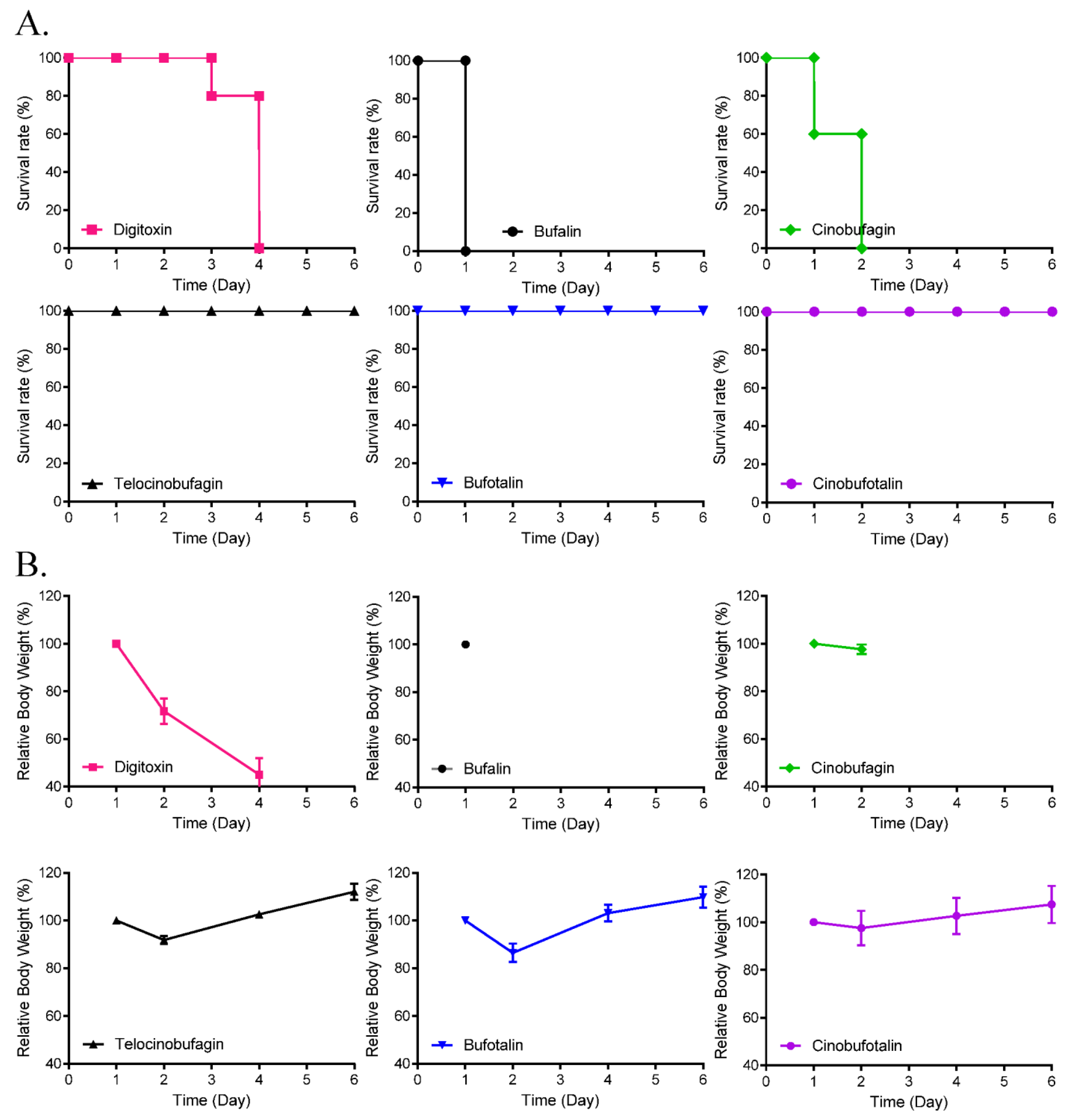
3.4. Toxicity and Pharmacokinetics of Cinobufagin and Telocinobufagin
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Coronavirus Disease (COVID-19) Pandemic. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 22 September 2021).
- Amarelle, L.; Lecuona, E. The Antiviral Effects of Na, K-ATPase Inhibition: A Minireview. Int. J. Mol. Sci. 2018, 19, 2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, D.; Kumavath, R.; Barh, D.; Azevedo, V.; Ghosh, P. Anticancer and Antiviral Properties of Cardiac Glycosides: A Review to Explore the Mechanism of Actions. Molecules 2020, 25, 3596. [Google Scholar] [CrossRef]
- Burkard, C.; Verheije, M.H.; Haagmans, B.L.; van Kuppeveld, F.J.; Rottier, P.J.; Bosch, B.J.; de Haan, C.A. ATP1A1-mediated Src signaling inhibits coronavirus entry into host cells. J. Virol. 2015, 89, 4434–4448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.W.; Lee, Y.Z.; Hsu, H.Y.; Jan, J.T.; Lin, Y.L.; Chang, S.Y.; Peng, T.T.; Yang, R.B.; Liang, J.J.; Liao, C.C.; et al. Inhibition of SARS-CoV-2 by Highly Potent Broad-Spectrum Anti-Coronaviral Tylophorine-Based Derivatives. Front. Pharmacol. 2020, 11, 606097. [Google Scholar] [CrossRef]
- Prassas, I.; Diamandis, E.P. Novel therapeutic applications of cardiac glycosides. Nat. Rev. Drug Discov. 2008, 7, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Botelho, A.F.M.; Pierezan, F.; Soto-Blanco, B.; Melo, M.M. A review of cardiac glycosides: Structure, toxicokinetics, clinical signs, diagnosis and antineoplastic potential. Toxicon 2019, 158, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.L.; An, Y.L.; Zhang, Y.Z.; Li, Z.W.; Zhou, Y.; Lei, M.; Zhang, J.Q.; Qu, H.; Da, J.; Wu, W.Y.; et al. Quantitative analysis of fourteen bufadienolides in Venenum Bufonis crude drug and its Chinese patent medicines by ultra-high performance liquid chromatography coupled with tandem mass spectrometry. J. Ethnopharmacol. 2020, 251, 112490. [Google Scholar] [CrossRef] [PubMed]
- Kumavath, R.; Paul, S.; Pavithran, H.; Paul, M.K.; Ghosh, P.; Barh, D.; Azevedo, V. Emergence of Cardiac Glycosides as Potential Drugs: Current and Future Scope for Cancer Therapeutics. Biomolecules 2021, 11, 1275. [Google Scholar] [CrossRef]
- Wei, W.L.; Hou, J.J.; Wang, X.; Yu, Y.; Li, H.J.; Li, Z.W.; Feng, Z.J.; Qu, H.; Wu, W.Y.; Guo, D.A. Venenum bufonis: An overview of its traditional use, natural product chemistry, pharmacology, pharmacokinetics and toxicology. J. Ethnopharmacol. 2019, 237, 215–235. [Google Scholar] [CrossRef]
- Souza, E.S.K.F.C.; Moraes, B.P.T.; Paixao, I.; Burth, P.; Silva, A.R.; Goncalves-de-Albuquerque, C.F. Na(+)/K(+)-ATPase as a Target of Cardiac Glycosides for the Treatment of SARS-CoV-2 Infection. Front. Pharmacol. 2021, 12, 624704. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R.C. Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org (accessed on 22 September 2021).
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Gene Ontol. Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Maxwell, E.S.; Fournier, M.J. The small nucleolar RNAs. Annu. Rev. Biochem. 1995, 64, 897–934. [Google Scholar] [CrossRef]
- Bachellerie, J.P.; Cavaille, J.; Huttenhofer, A. The expanding snoRNA world. Biochimie 2002, 84, 775–790. [Google Scholar] [CrossRef]
- Stepanov, G.A.; Filippova, J.A.; Komissarov, A.B.; Kuligina, E.V.; Richter, V.A.; Semenov, D.V. Regulatory role of small nucleolar RNAs in human diseases. Biomed. Res. Int. 2015, 2015, 206849. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Gralinski, L.; Ferris, M.T.; Frieman, M.B.; Thomas, M.J.; Proll, S.; Korth, M.J.; Tisoncik, J.R.; Heise, M.; Luo, S.; et al. Integrative deep sequencing of the mouse lung transcriptome reveals differential expression of diverse classes of small RNAs in response to respiratory virus infection. mBio 2011, 2, e00198-11. [Google Scholar] [CrossRef] [Green Version]
- Lafaille, F.G.; Harschnitz, O.; Lee, Y.S.; Zhang, P.; Hasek, M.L.; Kerner, G.; Itan, Y.; Ewaleifoh, O.; Rapaport, F.; Carlile, T.M.; et al. Human SNORA31 variations impair cortical neuron-intrinsic immunity to HSV-1 and underlie herpes simplex encephalitis. Nat. Med. 2019, 25, 1873–1884. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.L.; Sheng, J.; Rubin, D.H. A role for H/ACA and C/D small nucleolar RNAs in viral replication. Mol. Biotechnol. 2014, 56, 429–437. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound (1 μM) | MS (% of Remaining) | hERG Inhibition (%) | PPB (%) | CYP450 Inhibition (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mice | Rat | Human | Mice | Rat | 1A2 | 2C9 | 2C19 | 2D6 | 3A4 | ||
| CIN | 0.18 | 4.40 | 2.99 | 24.6 | 90.8 | 78.2 | 4.09 | 33.0 | 46.7 | 11.0 | 6.14 |
| ±0.07 | ±0.51 | ±0.39 | ±6.82 | ±1.74 | ±8.84 | ±1.0 | ±1.2 | ±9.3 | ±3.9 | ±5.3 | |
| TEL | 21.7 | 31.9 | 15.1 | 22.6 | 97.8 | 96.8 | 6.22 | 21.1 | 19.5 | 1.40 | 9.42 |
| ±0.90 | ±1.31 | ±1.27 | ±6.02 | ±0.35 | ±0.78 | ±1.3 | ±2.8 | ±11.1 | ±3.2 | ±5.3 | |
| Parameters (n = 3) | I.V., 1 mg/kg | P.O., 2 mg/kg |
|---|---|---|
| Tmax (h) | NA | 0.333 ± 0.144 |
| Cmax (μg/h) | NA | 0.537 ± 0.227 |
| T1/2 (h) | 0.225 ± 0.02 | 0.51 ± 0.407 |
| AUC (μg∙h/mL) | 0.604 ± 0.159 | 0.488 ± 0.204 |
| CL (L/h/kg) | 1.73 ± 0.432 | not applicable (NA) |
| Vss (L/kg) | 0.275 ± 0.169 | NA |
| Ft (%) | NA | 33.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Y.-H.; Jeon, S.; Lee, J.; Kim, S.; Jang, M.S.; Park, C.M.; Song, J.H.; Kim, H.R.; Kwon, S. Broad Spectrum Antiviral Properties of Cardiotonic Steroids Used as Potential Therapeutics for Emerging Coronavirus Infections. Pharmaceutics 2021, 13, 1839. https://doi.org/10.3390/pharmaceutics13111839
Jin Y-H, Jeon S, Lee J, Kim S, Jang MS, Park CM, Song JH, Kim HR, Kwon S. Broad Spectrum Antiviral Properties of Cardiotonic Steroids Used as Potential Therapeutics for Emerging Coronavirus Infections. Pharmaceutics. 2021; 13(11):1839. https://doi.org/10.3390/pharmaceutics13111839
Chicago/Turabian StyleJin, Young-Hee, Sangeun Jeon, Jihye Lee, Seungtaek Kim, Min Seong Jang, Chul Min Park, Jong Hwan Song, Hyoung Rae Kim, and Sunoh Kwon. 2021. "Broad Spectrum Antiviral Properties of Cardiotonic Steroids Used as Potential Therapeutics for Emerging Coronavirus Infections" Pharmaceutics 13, no. 11: 1839. https://doi.org/10.3390/pharmaceutics13111839
APA StyleJin, Y.-H., Jeon, S., Lee, J., Kim, S., Jang, M. S., Park, C. M., Song, J. H., Kim, H. R., & Kwon, S. (2021). Broad Spectrum Antiviral Properties of Cardiotonic Steroids Used as Potential Therapeutics for Emerging Coronavirus Infections. Pharmaceutics, 13(11), 1839. https://doi.org/10.3390/pharmaceutics13111839