
Glycan Profile Analysis of Engineered Trastuzumab with Rationally Added Glycosylation Sequons Presents Significantly Increased Glycan Complexity

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning and Mutation of HEK Trastuzumab and Glycosylation Mutants
2.2. Expression and Purification of HEK Trastuzumab and Glycosylation Mutants
2.3. LC–MS/MS Analysis of Tmab Variants
2.3.1. Glycopeptide Analysis
2.3.2. Free Glycan Analysis
2.4. Mass Spectrometry
2.5. Data Analysis
2.6. Binding Affinity to HER2 and Fc Receptors
2.7. Melting Temperature (Tm) and Onset Aggregation Temperature (Tagg) of Proteins
3. Results
3.1. Engineered Glycosylation Sites
3.2. Glycan Occupancy Prediction
3.3. Glycan Attachment Confirmation
3.4. Glycan Profile Analysis
3.5. Binding Affinity to HER2 and Fc Receptors
3.6. Tm and Tagg
4. Discussion
4.1. Preliminary Prediction and Confirmation of Glycan Attachment
4.2. Glycan Profile Analysis
4.3. Alterations in HER2 and Fc Receptor Affinity
4.4. Structural Stability and Aggregation Propensity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elgundi, Z.; Reslan, M.; Cruz, E.; Sifniotis, V.; Kayser, V. The state-of-play and future of antibody therapeutics. Adv. Drug Deliv. Rev. 2017, 122, 2–19. [Google Scholar] [CrossRef]
- Kaplon, H.; Reichert, J.M. Antibodies to watch in 2019. mAbs 2018, 11, 219–238. [Google Scholar] [CrossRef]
- Urquhart, L. Top companies and drugs by sales in 2019. Nat. Rev. Drug Discov. 2020, 19, 228. [Google Scholar] [CrossRef] [PubMed]
- Sifniotis, V.; Cruz, E.; Eroglu, B.; Kayser, V. Current Advancements in Addressing Key Challenges of Therapeutic Antibody Design, Manufacture, and Formulation. Antibodies 2019, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Cruz, E.; Kayser, V. Monoclonal antibody therapy of solid tumors: Clinical limitations and novel strategies to enhance treatment efficacy. Biologics 2019, 13, 33–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.K.H. The History of Monoclonal Antibody Development—Progress, Remaining Challenges and Future Innovations. Ann. Med. Surg. 2014, 3, 113–116. [Google Scholar] [CrossRef]
- Kayser, V.; Chennamsetty, N.; Voynov, V.; Helk, B.; Forrer, K.; Trout, B.L. Evaluation of a Non-Arrhenius Model for Therapeutic Monoclonal Antibody Aggregation. J. Pharm. Sci. 2011, 100, 2526–2542. [Google Scholar] [CrossRef] [PubMed]
- Kayser, V.; Chennamsetty, N.; Voynov, V.; Helk, B.; Trout, B.L. Conformational stability and aggregation of therapeutic monoclonal antibodies studied with ANS and Thioflavin T binding. mAbs 2011, 3, 408–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez-Rey, M.; Lang, D.A. Aggregates in monoclonal antibody manufacturing processes. Biotechnol. Bioeng. 2011, 108, 1494–1508. [Google Scholar] [CrossRef] [PubMed]
- Van der Kant, R.; Karow-Zwick, A.R.; Van Durme, J.; Blech, M.; Gallardo, R.; Seeliger, D.; Aßfalg, K.; Baatsen, P.; Compernolle, G.; Gils, A.; et al. Prediction and Reduction of the Aggregation of Monoclonal Antibodies. J. Mol. Biol. 2017, 429, 1244–1261. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Jiménez, J.; Martínez-Ortega, A.; Salmerón-García, A.; Cabeza, J.; Prados, J.C.; Ortíz, R.; Navas, N. Study of aggregation in therapeutic monoclonal antibodies subjected to stress and long-term stability tests by analyzing size exclusion liquid chromatographic profiles. Int. J. Biol. Macromol. 2018, 118, 511–524. [Google Scholar] [CrossRef]
- Voynov, V.; Chennamsetty, N.; Kayser, V.; Helk, B.; Trout, B.L. Predictive tools for stabilization of therapeutic proteins. mAbs 2009, 1, 580–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambrano, R.; Jamroz, M.; Szczasiuk, A.; Pujols, J.; Kmiecik, S.; Ventura, S. AGGRESCAN3D (A3D): Server for prediction of aggregation properties of protein structures. Nucleic Acids Res. 2015, 43, W306–W313. [Google Scholar] [CrossRef]
- Van Durme, J.; De Baets, G.; van der Kant, R.; Ramakers, M.; Ganesan, A.; Wilkinson, H.; Gallardo, R.; Rousseau, F.; Schymkowitz, J. Solubis: A webserver to reduce protein aggregation through mutation. Protein Eng. Des. Sel. 2016, 29, 285–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuyucak, S.; Kayser, V. Biobetters from an Integrated Computational/Experimental Approach. Comput. Struct. Biotechnol. J. 2017, 15, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Reslan, M.; Kayser, V. The effect of deuterium oxide on the conformational stability and aggregation of bovine serum albumin. Pharm. Dev. Technol. 2018, 23, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Razinkov, V.I.; Treuheit, M.J.; Becker, G.W. Accelerated Formulation Development of Monoclonal Antibodies (Mabs) and Mab-Based Modalities: Review of Methods and Tools. J. Biomol. Screen 2015, 20, 468–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reslan, M.; Ranganathan, V.; Macfarlane, D.R.; Kayser, V. Choline ionic liquid enhances the stability of Herceptin® (trastuzumab). Chem. Commun. 2018, 54, 10622–10625. [Google Scholar] [CrossRef]
- Reslan, M.; Kayser, V. Ionic liquids as biocompatible stabilizers of proteins. Biophys. Rev. 2018, 10, 781–793. [Google Scholar] [CrossRef]
- Mathaes, R.; Koulov, A.; Joerg, S.; Mahler, H.-C. Subcutaneous Injection Volume of Biopharmaceuticals—Pushing the Boundaries. J. Pharm. Sci. 2016, 105, 2255–2259. [Google Scholar] [CrossRef]
- Chennamsetty, N.; Helk, B.; Voynov, V.; Kayser, V.; Trout, B.L. Aggregation-Prone Motifs in Human Immunoglobulin G. J. Mol. Biol. 2009, 391, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Chennamsetty, N.; Voynov, V.; Kayser, V.; Helk, B.; Trout, B.L. Design of therapeutic proteins with enhanced stability. Proc. Natl. Acad. Sci. USA 2009, 106, 11937–11942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepinsky, R.B.; Silvian, L.; Berkowitz, S.A.; Farrington, G.; Lugovskoy, A.; Walus, L.; Eldredge, J.; Capili, A.; Mi, S.; Graff, C.; et al. Improving the solubility of anti-LINGO-1 monoclonal antibody Li33 by isotype switching and targeted mutagenesis. Protein Sci. 2010, 19, 954–966. [Google Scholar] [CrossRef] [Green Version]
- Kayser, V.; Chennamsetty, N.; Voynov, V.; Forrer, K.; Helk, B.; Trout, B.L. Glycosylation influences on the aggregation propensity of therapeutic monoclonal antibodies. Biotechnol. J. 2011, 6, 38–44. [Google Scholar] [CrossRef]
- Courtois, F.; Agrawal, N.J.; Lauer, T.M.; Trout, B.L. Rational design of therapeutic mAbs against aggregation through protein engineering and incorporation of glycosylation motifs applied to bevacizumab. mAbs 2016, 8, 99–112. [Google Scholar] [CrossRef] [Green Version]
- Reslan, M.; Sifniotis, V.; Cruz, E.; Sumer-Bayraktar, Z.; Cordwell, S.; Kayser, V. Enhancing the stability of adalimumab by engineering additional glycosylation motifs. Int. J. Biol. Macromol. 2020, 158, 189–196. [Google Scholar] [CrossRef]
- Jensen, P.H.; Karlsson, N.G.; Kolarich, D.; Packer, N.H. Structural analysis of N- and O-glycans released from glycoproteins. Nat. Protoc. 2012, 7, 1299–1310. [Google Scholar] [CrossRef]
- Nakamura, H.; Oda-Ueda, N.; Ueda, T.; Ohkuri, T. Introduction of a glycosylation site in the constant region decreases the aggregation of adalimumab Fab. Biochem. Biophys. Res. Commun. 2018, 503, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.; Bagchi, P.; Dutta, D.; Mukherjee, A.; Kobayashi, N.; Chawlasarkar, M. Computational identification of the post-translational modification sites and the functional family prediction reveals possible moonlighting role of rotaviral proteins. Bioinformation 2010, 4, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Eshaghi, A.; Duvvuri, V.; Lai, R.; Nadarajah, J.T.; Li, A.; Patel, S.N.; Low, D.E.; Gubbay, J.B. Genetic Variability of Human Respiratory Syncytial Virus A Strains Circulating in Ontario: A Novel Genotype with a 72 Nucleotide G Gene Duplication. PLoS ONE 2012, 7, e32807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, E.; Cain, J.; Crossett, B.; Kayser, V. Site-specific glycosylation profile of influenza A (H1N1) hemagglutinin through tandem mass spectrometry. Hum. Vaccines Immunother. 2018, 14, 508–517. [Google Scholar] [CrossRef]
- Chuang, G.-Y.; Boyington, J.C.; Joyce, M.G.; Zhu, J.; Nabel, G.J.; Kwong, P.D.; Georgiev, I. Computational prediction of N-linked glycosylation incorporating structural properties and patterns. Bioinformatics 2012, 28, 2249–2255. [Google Scholar] [CrossRef]
- Xie, H.; Chakraborty, A.; Ahn, J.; Yu, Y.Q.; Dakshinamoorthy, D.P.; Gilar, M.; Chen, W.; Skilton, S.J.; Mazzeo, J.R. Rapid comparison of a candidate biosimilar to an innovator monoclonal antibody with advanced liquid chromatography and mass spectrometry technologies. mAbs 2010, 2, 379–394. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.H.; Lee, J.; Bae, J.S.; Kim, Y.J.; Kang, H.A.; Kim, S.H.; Lee, S.J.; Lim, K.J.; Lee, J.W.; Jung, S.K.; et al. Analytical similarity assessment of rituximab biosimilar CT-P10 to reference medicinal product. mAbs 2018, 10, 380–396. [Google Scholar] [CrossRef]
- Croset, A.; Delafosse, L.; Gaudry, J.-P.; Arod, C.; Glez, L.; Losberger, C.; Begue, D.; Krstanovic, A.; Robert, F.; Vilbois, F.; et al. Differences in the glycosylation of recombinant proteins expressed in HEK and CHO cells. J. Biotechnol. 2012, 161, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Girardi, E.; Holdom, M.D.; Davies, A.M.; Sutton, B.J.; Beavil, A.J. The Crystal Structure of Rabbit Igg-Fc. Biochem. J. 2009, 417, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Van de Bovenkamp, F.S.; Derkesn, N.I.L.; Ooijevaar-de Heer, P.; van Schie, K.A.; Kruithof, S.; Berkowska, M.A.; van der Schoot, C.E.; IJspeert, H.; van der Burg, M.; Gils, A.; et al. Adaptive Antibody Diversification through ≪Em≫N≪/Em≫-Linked Glycosylation of The Immunoglobulin Variable Region. Proc. Natl. Acad. Sci. USA 2018, 115, 1901–1906. [Google Scholar] [PubMed] [Green Version]
- Jefferies, R. Glycosylation of Antibody Molecules. In Handbook of Therapeutic Antibodies; Dübel, S., Reichert, J.M., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2014; pp. 171–200. [Google Scholar]
- Anumula, K.R. Quantitative glycan profiling of normal human plasma derived immunoglobulin and its fragments Fab and Fc. J. Immunol. Methods 2012, 382, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Bondt, A.; Rombouts, Y.; Selman, M.H.J.; Hensbergen, P.J.; Reiding, K.R.; Hazes, J.M.W.; Dolhain, R.J.E.M.; Wuhrer, M. Immunoglobulin G (IgG) Fab Glycosylation Analysis Using a New Mass Spectrometric High-throughput Profiling Method Reveals Pregnancy-associated Changes. Mol. Cell. Proteom. 2014, 13, 3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, M.; Yagi, H.; Takahashi, N.; Kato, K.; Savage, C.O.S.; Goodall, D.M.; Jefferis, R. Differential glycosylation of polyclonal IgG, IgG-Fc and IgG-Fab isolated from the sera of patients with ANCA-associated systemic vasculitis. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2006, 1760, 669–677. [Google Scholar] [CrossRef]
- Boyd, P.; Lines, A.; Patel, A. The effect of the removal of sialic acid, galactose and total carbohydrate on the functional activity of Campath-1H. Mol. Immunol. 1995, 32, 1311–1318. [Google Scholar] [CrossRef]
- Dakshinamurthy, P.; Mukunda, P.; Kodaganti, B.P.; Shenoy, B.R.; Natarajan, B.; Maliwalave, A.; Halan, V.; Murugesan, S.; Maity, S. Charge variant analysis of proposed biosimilar to Trastuzumab. Biologicals 2017, 46, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Lakayan, D.; Haselberg, R.; Gahoual, R.; Somsen, G.W.; Kool, J. Affinity profiling of monoclonal antibody and antibody-drug-conjugate preparations by coupled liquid chromatography-surface plasmon resonance biosensing. Anal. Bioanal. Chem. 2018, 410, 7837–7848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lua, W.-H.; Gan, S.K.-E.; Lane, D.P.; Verma, C.S. A search for synergy in the binding kinetics of Trastuzumab and Pertuzumab whole and F(ab) to Her2. NPJ Breast Cancer 2015, 1, 15012. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| aa Substitution (Position) | Region | Proposed Function |
|---|---|---|
| L115N | VH | Mask APR by introducing glycosylation site [26] |
| A121N | CH1 | Improve solubility by introducing glycosylation site [25] |
| L177N | Mutate aa with high spatial-aggregation propensity and introduce glycosylation site [26] | |
| Q178N | Sterically hinder self-association by introducing glycosylation site and increase solubility [25,26] | |
| L182N | Mask APR by introducing glycosylation site [26] | |
| T198N | Improve solubility and sterically hinder self-association by introducing glycosylation site [25] | |
| Q160N | Ck | Mask APR by introducing glycosylation site [26] |
| aa Substitution (Position) | Region | Sequon | NetNGlyc | NGlycPred | SASA (Å2) |
|---|---|---|---|---|---|
| Q160N | CK | 160 NESV | 0.647 | 0.834 | 62.478 |
| L115N | VH | 115 NVTV | 0.679 | 0.462 | 43.709 |
| A121N | CH1 | 121 NSTK | 0.547 | 0.686 | 85.338 |
| L177N | CH1 | 177 NQSS | 0.537 | 1.000 | 96.039 |
| Q178N | 178 NSSG | 0 | 0.157 | 20.5 | |
| L182N | 182 NYSL | 0.568 | 0.999 | 30.123 | |
| T198N | 198 NQTY | 0.592 | 1.000 | 141.149 |
| Mass (Da) | Position | MC | Sequence |
|---|---|---|---|
| 4000.01 | 162–201 | 1 | NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTY |
| 3248.63 | 153–183 | 1 | FPEPVTVSWNSGALTSGVHTFPAVLQSSGLY |
| 3148.59 | 184–213 | 1 | SLSSVVTVPSSSLGTQTYIC NVNHKPSNTK |
| 2206.11 | 162–183 | 0 | NSGALTSGVHTFPAVLQSSG LY |
| 1812.92 | 184–201 | 0 | SLSSVVTVPSSSLGTQTY |
| # Structure | Type | Composition | Structure | [M-2H]2− | Glycosite |
|---|---|---|---|---|---|
| 1a | high mannose | HexNAc(2)Hex(5) |  | 617.22 | Q178N |
| 1b | high mannose | HexNAc(2)Hex(5) |  | 617.22 | L177N Q178N L182N |
| 2a (G0) | complex | HexNAc(4)Hex(3) |  | 658.24 | L177N Herceptin |
| 2b (G0) | complex | HexNAc(4)Hex(3) |  | 658.24 | L177N |
| 3 | high mannose | HexNAc(2)Hex(6) |  | 698.25 | Q178N L182N |
| 4 (G0F) | complex | HexNAc(4)Hex(3)Fuc(1) |  | 731.27 | All Tmab variants |
| 5 | complex | HexNAc(5)Hex(3) |  | 759.78 | L177N |
| 6a | hybrid | HexNAc(3)Hex(4)Man(2) Fuc(1) |  | 791.79 | L177N |
| 6b | hybrid | HexNAc(3)Hex(4)Man(2) Fuc(1) |  | 791.79 | L177N |
| 6c | hybrid | HexNAc(3)Hex(4)Man(2) Fuc(1) |  | 791.79 | L177N |
| 7 | hybrid | HexNAc(3)Hex(5)Man(1) |  | 799.79 | Q178N L182N |
| 8a (G1F) | complex | HexNAc(4)Hex(3)Man(1) Fuc(1) |  | 812.3 | L177N |
| 8b (G1F) | complex | HexNAc(4)Hex(3)Man(1) Fuc(1) |  | 812.3 | All Tmab variants |
| 9 | complex | HexNAc(5)Hex(3)Man(1) |  | 840.81 | L177N |
| 10 | high mannose | HexNAc(2)Hex(8) |  | 860.3 | L115N |
| 11 | hybrid | HexNAc(3)Hex(4)Man(1) NeuAc(1) |  | 864.31 | L182N |
| 12 | hybrid | HexNAc(3)Hex(5)Man(1) Fuc(1) |  | 872.81 | L177N |
| 13 (G2F) | complex | HexNAc(4)Hex(3)Man(2) Fuc(1) |  | 893.33 | All Tmab variants |
| 14a | complex | HexNAc(5)Hex(3)Man(1) Fuc(1) |  | 913.84 | A121N L177N Q178N L182N T198N |
| 14b | complex | HexNAc(5)Hex(3)Man(1) Fuc(1) |  | 913.84 | A121N L177N Q178N L182N T198N |
| 15 | high mannose | HexNAc(2)Hex(9) | 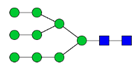 | 941.33 | L115N |
| 16 | hybrid | HexNAc(3)Hex(5)Man(1) NeuAc(1) |  | 944.84 | A121N L177N L182N T198N |
| 17a | complex | HexNAc(4)Hex(3)Man(2) NeuAc(1) |  | 965.84 | L177N L182N |
| 17b | complex | HexNAc(4)Hex(3)Man(2) NeuAc(1) |  | 965.84 | L177N |
| 18 | complex | HexNAc(5)Hex(3)Man(1) Fuc(2) |  | 986.87 | A121N L177N |
| 19a | complex | HexNAc(5)Hex(3)Man(2) Fuc(1) |  | 994.87 | L115N A121N L177N Q178N L182N T198N |
| 19b | complex | HexNAc(5)Hex(3)Man(2) Fuc(1) |  | 994.87 | L177N |
| 20a | complex | HexNAc(4)Hex(3)Man(2) Fuc(1)NeuAc(1) |  | 1038.87 | L115N A121N L177N Q178N L182N T198N |
| 20b | complex | HexNAc(4)Hex(3)Man(2) NeuAc(1) |  | 1038.87 | A121N L177N Q178N L182N T198N |
| 21a | complex | HexNAc(5)Hex(3)Man(1) NeuAc(1) |  | 1059.39 | A121N L177N T198N |
| 21b | complex | HexNAc(5)Hex(3)Man(1) NeuAc(1) |  | 1059.39 | L177N |
| 21c | complex | HexNAc(5)Hex(3)Man(1) NeuAc(1) |  | 1059.39 | L177N |
| Protein | HER2 | FcγR1A | ||||
|---|---|---|---|---|---|---|
| Ka (M−1 × s−1) | Kd (s−1) | KD (pM) | Ka (M−1 × s−1) | Kd (s−1) | KD (nM) | |
| Herceptin | 1.3 × 106 | 2.4 × 10−5 | 18 | 2.4 × 105 | 1.1 × 10−3 | 4.7 |
| HEK Trastuzumab | 7.9 × 105 | 2.7 × 10−5 | 34 | 1.1 × 105 | 1.0 × 10−3 | 9.9 |
| L115N | 2.4 × 105 | 2.7 × 10−4 | 1130 | 7.8 × 105 | 5.0 × 10−4 | 6.4 |
| L115N | 2.4 × 105 | 2.8 × 10−4 | 1160 | 1.1 × 105 | 4.0 × 10−4 | 3.7 |
| A121N | 3.8 × 105 | 3.0 × 10−4 | 790 | 1.7 × 105 | 8.3 × 10−4 | 4.8 |
| A121N | 4.1 × 105 | 2.7 × 10−4 | 640 | 2.0 × 105 | 9.8 × 10−4 | 4.8 |
| Q160N | 2.2 × 105 | 4.3 × 10−5 | 198 | 1.2 × 105 | 7.6 × 10−4 | 6.5 |
| Q160N | Responses to low for accurate fit | 1.1 × 105 | 7.6 × 10−4 | 6.7 | ||
| L177N | 4.0 × 105 | 1.9 × 10−4 | 480 | 2.5 × 105 | 1.4 × 10−3 | 5.6 |
| L177N | 4.5 × 105 | 2.4 × 10−4 | 530 | 2.6 × 105 | 1.5 × 10−3 | 5.6 |
| Q178N | 3.9 × 105 | 1.8 × 10−4 | 480 | 3.7 × 105 | 9.6 × 10−4 | 2.6 |
| Q178N | 3.8 × 105 | 8.7 × 10−5 | 230 | 3.8 × 105 | 9.6 × 10−4 | 2.5 |
| L182N | 3.6 × 105 | 3.9 × 10−5 | 109 | 1.3 × 105 | 1.1 × 10-3 | 8.2 |
| L182N | 5.6 × 105 | 6.4 × 10−5 | 110 | 1.8 × 105 | 1.5 × 10−3 | 8.1 |
| T198N | 2.5 × 105 | 2.3 × 10−4 | 920 | 2.4 × 105 | 8.8 × 10−4 | 3.8 |
| T198N | 4.6 × 105 | 2.5 × 10−4 | 550 | 2.4 × 105 | 9.0 × 10−4 | 3.8 |
| Protein | FcγR2B | FcγR3A |
|---|---|---|
| KD (nM) | KD (nM) | |
| Herceptin | 666 ± 63 | 267 ± 34 |
| HEK Trastuzumab | 686 ± 127 | 433 ± 65 |
| L115N | 338 ±100 | 394 ± 37 |
| L115N | 269 ± 160 | 301 ± 40 |
| A121N | 3142 ± 340 | 295 ± 17 |
| A121N | 3743 ± 720 | 268 ± 14 |
| Q160N | 143 ± 43 | 596 ± 100 |
| Q160N | poor data set | 535 ± 67 |
| L177N | 1275 ± 120 | 535 ± 79 |
| L177N | 992 ± 230 | 551 ± 71 |
| Q178N | Responses too low for accurate fit | 239 ± 25 |
| Q178N | 485 ± 240 | 217 ± 30 |
| L182N | No observable binding | ND |
| L182N | ||
| T198N | Responses too low for accurate fit | ND |
| T198N | 666 ± 63 |
| Protein | Tm1 (°C) | Tm2 (°C) | Tagg @ 266 nm |
|---|---|---|---|
| Herceptin | 68.4 ± 0.2 | 82.5 ± 0.1 | 81.2 ± 0.1 |
| Tmab | 68.2 ± 0.1 | 83.1 ± 0.3 | 85.2 ± 0.1 |
| L115N | 73.1 ± 0.7 | 84.3 ± 0.4 | 91.2 ± 0.3 |
| A121N | 68.8 ± 0.2 | 80.7 ± 0.5 | 85.2 ± 0.1 |
| L177N | 71.2 ± 0.3 | 79.1 ± 0.3 | 85.1 ± 0.1 |
| Q160N | ND | ND | ND |
| Q178N | 69.1 ± 0.2 | 81.1 ± 0.4 | 85.5 ± 0.1 |
| L182N | 73.1 ± 1.9 | 81.0 ± 0.3 | 86.1 ± 0.1 |
| T198N | 67.9 ± 0.4 | 83.3 ± 0.4 | 87.8 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz, E.; Sifniotis, V.; Sumer-Bayraktar, Z.; Reslan, M.; Wilkinson-White, L.; Cordwell, S.; Kayser, V. Glycan Profile Analysis of Engineered Trastuzumab with Rationally Added Glycosylation Sequons Presents Significantly Increased Glycan Complexity. Pharmaceutics 2021, 13, 1747. https://doi.org/10.3390/pharmaceutics13111747
Cruz E, Sifniotis V, Sumer-Bayraktar Z, Reslan M, Wilkinson-White L, Cordwell S, Kayser V. Glycan Profile Analysis of Engineered Trastuzumab with Rationally Added Glycosylation Sequons Presents Significantly Increased Glycan Complexity. Pharmaceutics. 2021; 13(11):1747. https://doi.org/10.3390/pharmaceutics13111747
Chicago/Turabian StyleCruz, Esteban, Vicki Sifniotis, Zeynep Sumer-Bayraktar, Mouhamad Reslan, Lorna Wilkinson-White, Stuart Cordwell, and Veysel Kayser. 2021. "Glycan Profile Analysis of Engineered Trastuzumab with Rationally Added Glycosylation Sequons Presents Significantly Increased Glycan Complexity" Pharmaceutics 13, no. 11: 1747. https://doi.org/10.3390/pharmaceutics13111747
APA StyleCruz, E., Sifniotis, V., Sumer-Bayraktar, Z., Reslan, M., Wilkinson-White, L., Cordwell, S., & Kayser, V. (2021). Glycan Profile Analysis of Engineered Trastuzumab with Rationally Added Glycosylation Sequons Presents Significantly Increased Glycan Complexity. Pharmaceutics, 13(11), 1747. https://doi.org/10.3390/pharmaceutics13111747