
Candida glabrata Antifungal Resistance and Virulence Factors, a Perfect Pathogenic Combination

 , , ,
, , ,  ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
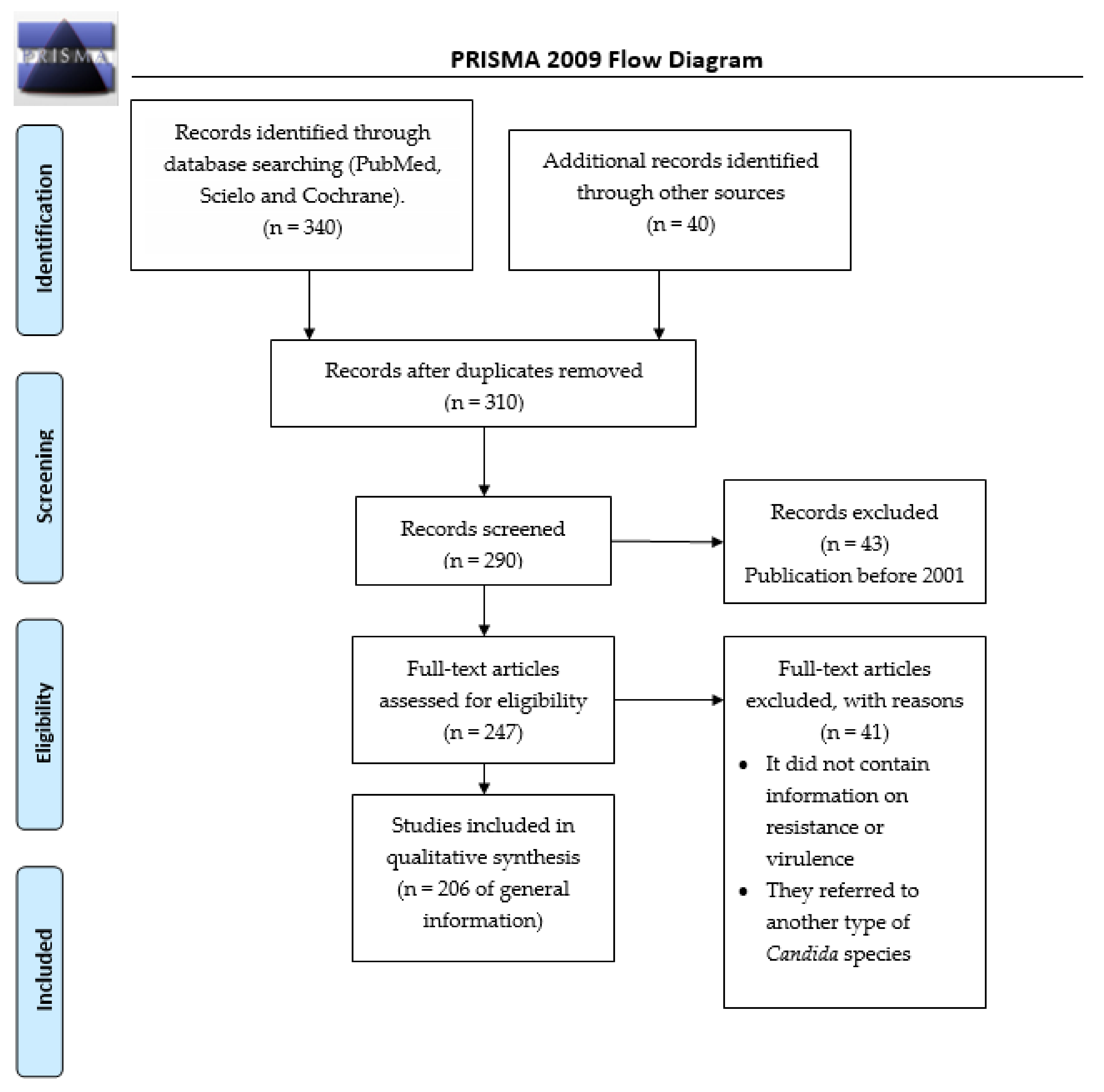
2. Materials and Methods
3. Immunological Regulation of the Host
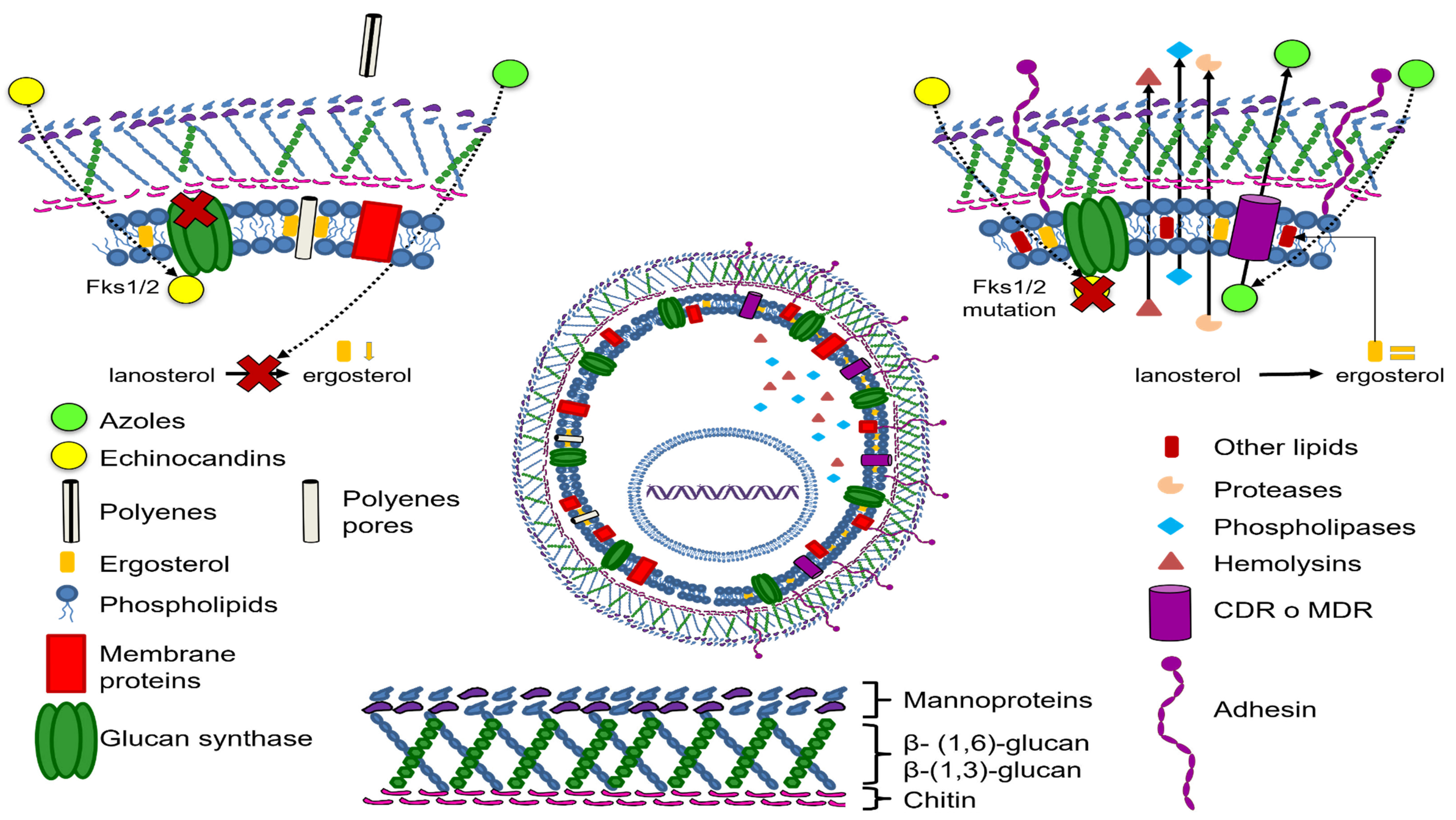
4. Antifungal Resistance of the C. glabrata Complex
4.1. Resistance to Azoles
4.2. Susceptibility Variations per Continent
4.3. Drug Resistance Fluctuations Caused by the Type of Candida and Genetic Variations
4.4. Drug Resistance Variations in C. Glabrata Complex Species
5. Virulence Factors
Adherence Mechanisms
6. Enzymatic Activity
6.1. Proteases/Proteinases/Aspartyl Proteases
6.2. Phospholipase
6.3. Esterase
6.4. Hemolysin
7. Biofilm Formation
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bitar, D.; Lortholary, O.; Le Strat, Y.; Nicolau, J.; Coignard, B.; Tattevin, P.; Che, D.; Dromer, F. Population-Based Analysis of Invasive Fungal Infections, France, 2001–2010. Emerg. Infect. Dis. 2014, 20, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; He, C.; Zhao, C.; Chen, X.; Hua, H.; Yan, Z. Characterization of Oral Candidiasis and the Candida Species Profile in Patients with Oral Mucosal Diseases. Microb. Pathog. 2019, 134, 103575. [Google Scholar] [CrossRef]
- Boan, P.; Gardam, D. Epidemiology and Antifungal Susceptibility Patterns of Candidemia from a Tertiary Centre in Western Australia. J. Chemother. 2019, 31, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, M.A.; Melo, A.P.; Bento, A.D.; Souza, L.B.; Neto, F.D.; Garcia, J.B.; Zuza-Alves, D.L.; Francisco, E.C.; Melo, A.S.; Chaves, G.M. Epidemiology and Prognostic Factors of Nosocomial Candidemia in Northeast Brazil: A Six-Year Retrospective Study. PLoS ONE 2019, 14, e0221033. [Google Scholar] [CrossRef]
- Ryan, P.; Motherway, C.; Powell, J.; Elsaka, A.; Sheikh, A.A.; Jahangir, A.; O’Connell, N.H.; Dunne, C.P. Candidaemia in an Irish Intensive Care Unit Setting between 2004 and 2018 Reflects Increased Incidence of Candida glabrata. J. Hosp. Infect. 2019, 102, 347–350. [Google Scholar] [CrossRef]
- Güzel, A.B.; Küçükgöz-Güleç, Ü.; Aydin, M.; Gümral, R.; Kalkanci, A.; Ilkit, M. Candida Vaginitis during Contraceptive Use: The Influence of Methods, Antifungal Susceptibility and Virulence Patterns. J. Obstet. Gynaecol. 2013, 33, 850–856. [Google Scholar] [CrossRef]
- Silva, M.G.C.; Rodrigues, G.S.; Gonçalves, I.L.; Grazziotin, N.A. Candida Species Distribution and Fluconazole Susceptibility of Blood Isolates at a Regional Hospital in Passo Fundo, RS, Brazil. Bras. Patol. Med. Lab. 2015, 51, 158–161. [Google Scholar] [CrossRef]
- Neves-Junior, A.; Cartágenes-Pinto, A.C.; Rocha, D.A.; Sá, L.F.; Junqueira, M.D.; Ferreira-Pereira, A. Prevalence and Fluconazole Susceptibility Profile of Candida spp. Clinical Isolates in a Brazilian Tertiary Hospital in Minas Gerais, Brazil. An. Acad. Bras. Ciênc. 2015, 87, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Ferreira, C.; Alves, C.T.; Henriques, M.; Azeredo, J.; Silva, S. Vulvovaginal Candidiasis: Epidemiology, Microbiology and Risk Factors. Crit. Rev. Microbiol. 2016, 42, 905–927. [Google Scholar] [CrossRef]
- Braga, P.R.; Cruz, I.L.; Ortiz, I.; Barreiros, G.; Nouér, S.A.; Nucci, M. Secular Trends of Candidemia at a Brazilian Tertiary Care Teaching Hospital. Braz. J. Infect. Dis. 2018, 22, 273–277. [Google Scholar] [CrossRef]
- Israel, S.; Amit, S.; Israel, A.; Livneh, A.; Nir-Paz, R.; Korem, M. The Epidemiology and Susceptibility of Candidemia in Jerusalem, Israel. Front. Cell. Infect. Microbiol. 2019, 9, 352. [Google Scholar] [CrossRef] [PubMed]
- Maraki, S.; Mavromanolaki, V.E.; Stafylaki, D.; Nioti, E.; Hamilos, G.; Kasimati, A. Epidemiology and Antifungal Susceptibility Patterns of Candida Isolates from Greek Women with Vulvovaginal Candidiasis. Mycoses 2019, 62, 692–697. [Google Scholar] [CrossRef]
- Tarapan, S.; Matangkasombut, O.; Trachootham, D.; Sattabanasuk, V.; Talungchit, S.; Paemuang, W.; Phonyiam, T.; Chokchaitam, O.; Mungkung, O.; Lam-ubol, A. Oral Candida Colonization in Xerostomic Postradiotherapy Head and Neck Cancer Patients. Oral Dis. 2019, 25, 1798–1808. [Google Scholar] [CrossRef] [PubMed]
- Tsega, A.; Mekonnen, F. Prevalence, Risk Factors and Antifungal Susceptibility Pattern of Candida Species among Pregnant Women at Debre Markos Referral Hospital, Northwest Ethiopia. BMC Pregnancy Childbirth 2019, 19, 527. [Google Scholar] [CrossRef]
- Matic Petrovic, S.; Radunovic, M.; Barac, M.; Kuzmanovic Pficer, J.; Pavlica, D.; Arsic Arsenijevic, V.; Pucar, A. Subgingival Areas as Potential Reservoirs of Different Candida spp. in Type 2 Diabetes Patients and Healthy Subjects. PLoS ONE 2019, 14, e0210527. [Google Scholar] [CrossRef]
- Santolaya, M.E.; Thompson, L.; Benadof, D.; Tapia, C.; Legarraga, P.; Cortés, C.; Rabello, M.; Valenzuela, R.; Rojas, P.; Rabagliati, R.; et al. A Prospective, Multi-Center Study of Candida Bloodstream Infections in Chile. PLoS ONE 2019, 14, e0212924. [Google Scholar] [CrossRef]
- Alrayyes, S.F.; Alruwaili, H.M.; Taher, I.A.; Elrahawy, K.M.; Almaeen, A.H.; Ashekhi, A.O.; Alam, M.K. Oral Candida Carriage and Associated Risk Indicators among Adults in Sakaka, Saudi Arabia. BMC Oral Health 2019, 19, 86. [Google Scholar] [CrossRef]
- Djohan, V.; Angora, K.E.; Vanga-Bosson, A.H.; Konaté, A.; Kassi, K.F.; Kiki-Barro, P.C.M.; Bedia-Tanoh, A.V.; Miezan, S.; Menan, E.I.H.; Yavo, W. Recurrent Vulvo-Vaginal Candidiasis in Abidjan (Côte d’Ivoire): Aetiology and Associated Factors. J. Mycol. Med. 2019, 29, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, S.I.; Choi, J.Y.; Yoon, S.K.; Na, G.H.; You, Y.K.; Kim, D.G.; Kim, M.S.; Lee, J.G.; Joo, D.J.; et al. Invasive Fungal Infection in Liver Transplant Recipients in a Prophylactic Era: A Multicenter Retrospective Cohort Study in Korea. Medicine 2019, 98, e16179. [Google Scholar] [CrossRef]
- Zeng, Z.; Tian, G.; Ding, Y.; Yang, K.; Liu, J.; Deng, J. Surveillance Study of the Prevalence, Species Distribution, Antifungal Susceptibility, Risk Factors and Mortality of Invasive Candidiasis in a Tertiary Teaching Hospital in Southwest China. BMC Infect. Dis. 2019, 19, 939. [Google Scholar] [CrossRef]
- Jain, A.K.; Misra, V.; Ranjan, N.; Jain, S.B.; Gandhi, S. Speciation, Biofilm Formation and Antifungal Susceptibility of Candida Isolates from Clinically Diagnosed Patient of UTI in a Tertiary Care Hospital. J. Assoc. Physicians India 2019, 67, 42–45. [Google Scholar] [PubMed]
- Ghaddar, N.; Anastasiadis, E.; Halimeh, R.; Ghaddar, A.; Dhar, R.; AlFouzan, W.; Yusef, H.; El Chaar, M. Prevalence and Antifungal Susceptibility of Candida albicans Causing Vaginal Discharge among Pregnant Women in Lebanon. BMC Infect. Dis. 2020, 20, 32. [Google Scholar] [CrossRef]
- Kumar, A.; Nair, R.; Kumar, M.; Banerjee, A.; Chakrabarti, A.; Rudramurthy, S.M.; Bagga, R.; Gaur, N.A.; Mondal, A.K.; Prasad, R. Assessment of Antifungal Resistance and Associated Molecular Mechanism in Candida albicans Isolates from Different Cohorts of Patients in North Indian State of Haryana. Folia Microbiol. 2020, 65, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Siopi, M.; Tarpatzi, A.; Kalogeropoulou, E.; Damianidou, S.; Vasilakopoulou, A.; Vourli, S.; Pournaras, S.; Meletiadis, J. Epidemiological Trends of Fungemia in Greece with a Focus on Candidemia during the Recent Financial Crisis: A 10-Year Survey in a Tertiary Care Academic Hospital and Review of Literature. Antimicrob. Agents Chemother. 2020, 64, e01516-19. [Google Scholar] [CrossRef] [PubMed]
- Kord, M.; Salehi, M.; Khodavaisy, S.; Hashemi, S.J.; Daie Ghazvini, R.; Rezaei, S.; Maleki, A.; Elmimoghaddam, A.; Alijani, N.; Abdollahi, A.; et al. Epidemiology of Yeast Species Causing Bloodstream Infection in Tehran, Iran (2015–2017); Superiority of 21-Plex PCR over the Vitek 2 System for Yeast Identification. J. Med. Microbiol. 2020, 69, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.C.; Laghi, L.; Parolin, C.; Foschi, C.; Marangoni, A.; Liberatore, A.; Dias, A.L.T.; Cricca, M.; Vitali, B. Metabolic Profiling of Candida Clinical Isolates of Different Species and Infection Sources. Sci. Rep. 2020, 10, 16716. [Google Scholar] [CrossRef] [PubMed]
- Gabaldón, T.; Carreté, L. The Birth of a Deadly Yeast: Tracing the Evolutionary Emergence of Virulence Traits in Candida glabrata. FEMS Yeast Res. 2016, 16, fov110. [Google Scholar] [CrossRef]
- Aldardeer, N.F.; Albar, H.; Al-Attas, M.; Eldali, A.; Qutub, M.; Hassanien, A.; Alraddadi, B. Antifungal Resistance in Patients with Candidaemia: A Retrospective Cohort Study. BMC Infect. Dis. 2020, 20, 55. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.; Sherry, L.; Nile, C.J.; Sherriff, A.; Johnson, E.M.; Hanson, M.F.; Williams, C.; Munro, C.A.; Jones, B.J.; Ramage, G. Biofilm Formation Is a Risk Factor for Mortality in Patients with Candida albicans Bloodstream Infection—Scotland, 2012–2013. Clin. Microbiol. Infect. 2016, 22, 87–93. [Google Scholar] [CrossRef]
- Aguilar, G.; Araujo, P.; Godoy, E.; Falcón, M.; Centurión, M.G.; Ortiz, R.; Britez, M.; Martínez, M. Identification and Characteristics of Candida spp. in Vaginal Secretion of Pregnant and Non-Pregnant Patients. Mem. Inst. Investig. Cienc. Salud 2017, 15, 6–12. [Google Scholar] [CrossRef][Green Version]
- Gülmez, D.; Sığ, A.K.; Akar, N.; Duyan, S.; Arıkan Akdağlı, S. Changing Trends in Isolation Frequencies and Species of Clinical Fungal Strains: What Do the 12-years (2008–2019) Mycology Laboratory Data Tell About? Mikrobiyol. Bul. 2021, 55, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Sganga, G.; Wang, M.; Capparella, M.R.; Tawadrous, M.; Yan, J.L.; Aram, J.A.; Montravers, P. Evaluation of Anidulafungin in the Treatment of Intra-Abdominal Candidiasis: A Pooled Analysis of Patient-Level Data from 5 Prospective Studies. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1849–1856. [Google Scholar] [CrossRef]
- Logan, C.; Martin-Loeches, I.; Bicanic, T. Invasive Candidiasis in Critical Care: Challenges and Future Directions. Intensive Care Med. 2020, 46, 2001–2014. [Google Scholar] [CrossRef]
- Ambe, N.F.; Longdoh, N.A.; Tebid, P.; Bobga, T.P.; Nkfusai, C.N.; Ngwa, S.B.; Nsai, F.S.; Cumber, S.N. The Prevalence, Risk Factors and Antifungal Sensitivity Pattern of Oral Candidiasis in HIV/AIDS Patients in Kumba District Hospital, South West Region, Cameroon. Pan Afr. Med. J. 2020, 36, 23. [Google Scholar] [CrossRef]
- Eghtedar Nejad, E.; Ghasemi Nejad Almani, P.; Mohammadi, M.A.; Salari, S. Molecular Identification of Candida Isolates by Real-time PCR-high-resolution Melting Analysis and Investigation of the Genetic Diversity of Candida Species. J. Clin. Lab. Anal. 2020, 34, e23444. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.R.; Horcajada, J.P.; Locke, J.B.; Viani, R.; Pappas, P.; Ghannoum, M.; Sandison, T.; Soriano, A. 1284. Outcomes by Baseline Pathogens and Susceptibility in the STRIVE Phase 2 Trial of Once-Weekly Rezafungin for Treatment of Candidemia and Invasive Candidiasis Compared with Caspofungin. Open Forum Infect. Dis. 2020, 7, S657–S658. [Google Scholar] [CrossRef]
- Fuller, J.; Dingle, T.C.; Bull, A.; Shokoples, S.; Laverdière, M.; Baxter, M.R.; Adam, H.J.; Karlowsky, J.A.; Zhanel, G.G.; Canadian Antimicrobial Resistance Alliance (CARA) and CANWARD; et al. Species Distribution and Antifungal Susceptibility of Invasive Candida Isolates from Canadian Hospitals: Results of the CANWARD 2011–16 Study. J. Antimicrob. Chemother. 2019, 74, iv48–iv54. [Google Scholar] [CrossRef] [PubMed]
- Ulrych, J.; Adámková, V.; Matek, J.; Komarc, M.; Frýba, V.; Schmidt, D.; Koželský, P.; Studená, A.; Bříza, J.; Krška, Z. Intra-Abdominal Candidiasis in Surgical Intensive Care Unit—Epidemiology Characteristics and Trends. Epidemiol. Mikrobiol. Imunol. 2020, 69, 57–63. [Google Scholar]
- Timmermans, B.; De Las Peñas, A.; Castaño, I.; Van Dijck, P. Adhesins in Candida glabrata. J. Fungi 2018, 4, 60. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, T.; Bai, C.; Zhang, Q.; Li, Z.; Li, X. A Predictive Nomogram for Mortality of Cancer Patients with Invasive Candidiasis: A 10-Year Study in a Cancer Center of North China. BMC Infect. Dis. 2021, 21, 76. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.S.; Desa, M.N.M.; Sandai, D.; Chong, P.P.; Than, L.T.L. Growth, Biofilm Formation, Antifungal Susceptibility and Oxidative Stress Resistance of Candida glabrata Are Affected by Different Glucose Concentrations. Infect. Genet. Evol. 2016, 40, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Chaves, G.M.; Diniz, M.G.; da Silva-Rocha, W.P.; de Souza, L.B.F.C.; Gondim, L.A.M.; Ferreira, M.A.F.; Svidzinski, T.I.E.; Milan, E.P. Species Distribution and Virulence Factors of Candida spp. Isolated from the Oral Cavity of Kidney Transplant Recipients in Brazil. Mycopathologia 2013, 175, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Polke, M.; Hube, B.; Jacobsen, I.D. Candida Survival Strategies. Adv. Appl. Microbiol. 2015, 91, 139–235. [Google Scholar] [CrossRef]
- Tiraboschi, I.N.; Pozzi, N.C.; Farías, L.; García, S.; Fernández, N.B. Epidemiología, Especies, Resistencia Antifúngica y Evolución de Las Candidemias En Un Hospital Universitario de Buenos Aires, Argentina, Durante 16 Años. Rev. Chil. Infectol. 2017, 34, 431–440. [Google Scholar] [CrossRef]
- Fernández-Ruiz, M.; Cardozo, C.; Salavert, M.; Aguilar-Guisado, M.; Escolà-Vergé, L.; Muñoz, P.; Gioia, F.; Montejo, M.; Merino, P.; Cuervo, G.; et al. Candidemia in Solid Organ Transplant Recipients in Spain: Epidemiological Trends and Determinants of Outcome. Transpl. Infect. Dis. 2019, 21, e13195. [Google Scholar] [CrossRef] [PubMed]
- Anderson, H.W. Yeast-Like Fungi of the Human Intestinal Tract. J. Infect. Dis. 1917, 21, 341–386. [Google Scholar] [CrossRef]
- Plaut, A. Human Infection with Cryptococcus glabratus: Report of Case Involving Uterus and Fallopian Tube. Am. J. Clin. Pathol. 1950, 20, 377–380. [Google Scholar] [CrossRef]
- Hazen, K.C. New and Emerging Yeast Pathogens. Clin. Microbiol. Rev. 1995, 8, 462–478. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Silva, S.; Henriques, M. Candida glabrata: A Review of Its Features and Resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 673–688. [Google Scholar] [CrossRef]
- Gabaldón, T.; Fairhead, C. Genomes Shed Light on the Secret Life of Candida glabrata: Not so Asexual, Not so Commensal. Curr. Genet. 2019, 65, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Savastano, C.; de Oliveira Silva, E.; Gonçalves, L.L.; Nery, J.M.; Silva, N.C.; Dias, A.L.T. Candida glabrata among Candida spp. from Environmental Health Practitioners of a Brazilian Hospital. Braz. J. Microbiol. 2016, 47, 367–372. [Google Scholar] [CrossRef]
- Sakita, K.M.; Faria, D.R.; da Silva, E.M.; Tobaldini-Valério, F.K.; Kioshima, E.S.; Svidzinski, T.I.; de Souza Bonfim-Mendonça, P. Healthcare Workers’ Hands as a Vehicle for the Transmission of Virulent Strains of Candida spp.: A Virulence Factor Approach. Microb. Pathog. 2017, 113, 225–232. [Google Scholar] [CrossRef] [PubMed]
- de Paula Menezes, R.; Silva, F.F.; Melo, S.G.; Alves, P.G.; Brito, M.O.; de Souza Bessa, M.A.; Amante Penatti, M.P.; Pedroso, R.S.; Abdallah, V.O.; Röder, D.V. Characterization of Candida Species Isolated from the Hands of the Healthcare Workers in the Neonatal Intensive Care Unit. Med. Mycol. 2019, 57, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Opulente, D.A.; Langdon, Q.K.; Buh, K.V.; Haase, M.A.B.; Sylvester, K.; Moriarty, R.V.; Jarzyna, M.; Considine, S.L.; Schneider, R.M.; Hittinger, C.T. Pathogenic Budding Yeasts Isolated Outside of Clinical Settings. FEMS Yeast Res. 2019, 19, foz032. [Google Scholar] [CrossRef]
- Turner, S.A.; Butler, G. The Candida Pathogenic Species Complex. Cold Spring Harb. Perspect. Med. 2014, 4, a019778. [Google Scholar] [CrossRef]
- Swoboda-Kopeć, E.; Sikora, M.; Golas, M.; Piskorska, K.; Gozdowski, D.; Netsvyetayeva, I. Candida nivariensis in Comparison to Different Phenotypes of Candida glabrata. Mycoses 2014, 57, 747–753. [Google Scholar] [CrossRef]
- Sikora, M.; Kuthan, R.; Piskorska-Malolepsza, K.; Golas-Pradzynska, M.; Domański, D.; Augustynowicz-Kopeć, E.; Swoboda-Kopec, E. Prevalence and Antifungal Susceptibility of the Emerging Fungal Species, Candida nivariensis, Isolated in a Teaching Hospital in Poland. Pol. J. Microbiol. 2019, 68, 303–308. [Google Scholar] [CrossRef]
- Małek, M.; Mrowiec, P.; Klesiewicz, K.; Skiba-Kurek, I.; Szczepański, A.; Białecka, J.; Żak, I.; Bogusz, B.; Kędzierska, J.; Budak, A.; et al. Prevalence of Human Pathogens of the Clade Nakaseomyces in a Culture Collection—The First Report on Candida bracarensis in Poland. Folia Microbiol. 2019, 64, 307–312. [Google Scholar] [CrossRef]
- Correia, A. Candida bracarensis sp. Nov., a Novel Anamorphic Yeast Species Phenotypically Similar to Candida glabrata. Int. J. Syst. Evol. Microbiol. 2006, 56, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Alcoba-Flórez, J.; Méndez-Álvarez, S.; Cano, J.; Guarro, J.; Pérez-Roth, E.; del Pilar Arévalo, M. Phenotypic and Molecular Characterization of Candida nivariensis sp. Nov., a Possible New Opportunistic Fungus. J. Clin. Microbiol. 2005, 43, 4107–4111. [Google Scholar] [CrossRef]
- Borman, A.M.; Petch, R.; Linton, C.J.; Palmer, M.D.; Bridge, P.D.; Johnson, E.M. Candida nivariensis, an Emerging Pathogenic Fungus with Multidrug Resistance to Antifungal Agents. J. Clin. Microbiol. 2008, 46, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Hernando-Ortiz, A.; Mateo, E.; Ortega-Riveros, M.; De-la-Pinta, I.; Quindós, G.; Eraso, E. Caenorhabditis elegans as a Model System To Assess Candida glabrata, Candida nivariensis, and Candida bracarensis Virulence and Antifungal Efficacy. Antimicrob. Agents Chemother. 2020, 64, e00824-20. [Google Scholar] [CrossRef] [PubMed]
- Treviño-Rangel, R.D.; Espinosa-Pérez, J.F.; Villanueva-Lozano, H.; Montoya, A.M.; Andrade, A.; Bonifaz, A.; González, G.M. First Report of Candida bracarensis in Mexico: Hydrolytic Enzymes and Antifungal Susceptibility Pattern. Folia Microbiol. 2018, 63, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhu, Y.; Fan, S.; Vitagliano, A.; Liu, X.; Liao, Y.; Liang, Y.; Vitale, S.G. Clinical Characteristics and Antifungal Susceptibility of Candida nivariensis from Vulvovaginal Candidiasis. Gynecol. Obstet. Investig. 2020, 85, 88–93. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Adherence and Biofilm Formation of Non-Candida albicans Candida Species. Trends Microbiol. 2011, 19, 241–247. [Google Scholar] [CrossRef]
- Rodrigues, C.F.; Silva, S.; Azeredo, J.; Henriques, M. Candida glabrata’s Recurrent Infections: Biofilm Formation during Amphotericin B Treatment. Lett. Appl. Microbiol. 2016, 63, 77–81. [Google Scholar] [CrossRef]
- Nakamura-Vasconcelos, S.S.; Fiorini, A.; Zanni, P.D.; de Souza Bonfim-Mendonça, P.; Godoy, J.R.; Almeida-Apolonio, A.A.; Consolaro, M.E.; Svidzinski, T.I. Emergence of Candida glabrata in Vulvovaginal Candidiasis Should Be Attributed to Selective Pressure or Virulence Ability? Arch. Gynecol. Obstet. 2017, 296, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-H.; Jung, D.S.; Lee, J.Y.; Kim, H.A.; Ryu, S.Y.; Jung, S.-I.; Joo, E.-J.; Cheon, S.; Kim, Y.-S.; Kim, S.-W.; et al. Changing Epidemiology of Non-albicans Candidemia in Korea. J. Infect. Chemother. 2019, 25, 388–391. [Google Scholar] [CrossRef]
- Wang, H.; Xu, Y.-C.; Hsueh, P.-R. Epidemiology of Candidemia and Antifungal Susceptibility in Invasive Candida Species in the Asia-Pacific Region. Future Microbiol. 2016, 11, 1461–1477. [Google Scholar] [CrossRef]
- Pote, S.T.; Sonawane, M.S.; Rahi, P.; Shah, S.R.; Shouche, Y.S.; Patole, M.S.; Thakar, M.R.; Sharma, R. Distribution of Pathogenic Yeasts in Different Clinical Samples: Their Identification, Antifungal Susceptibility Pattern, and Cell Invasion Assays. Infect. Drug Resist. 2020, 13, 1133–1145. [Google Scholar] [CrossRef]
- Kaan, Ö.; Koç, A.N.; Atalay, M.A.; Mutlu Sarigüzel, F. Molecular Epidemiology, Antifungal Susceptibility and Virulence Factors of Candida glabrata Complex Strains in Kayseri/Turkey. Microb. Pathog. 2021, 154, 104870. [Google Scholar] [CrossRef]
- Alves, R.; Barata-Antunes, C.; Casal, M.; Brown, A.J.P.; Van Dijck, P.; Paiva, S. Adapting to Survive: How Candida Overcomes Host-Imposed Constraints during Human Colonization. PLoS Pathog. 2020, 16, e1008478. [Google Scholar] [CrossRef] [PubMed]
- Seider, K.; Gerwien, F.; Kasper, L.; Allert, S.; Brunke, S.; Jablonowski, N.; Schwarzmüller, T.; Barz, D.; Rupp, S.; Kuchler, K.; et al. Immune Evasion, Stress Resistance, and Efficient Nutrient Acquisition Are Crucial for Intracellular Survival of Candida glabrata within Macrophages. Eukaryot. Cell 2014, 13, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Pais, P.; Galocha, M.; Teixeira, M.C. Genome-Wide Response to Drugs and Stress in the Pathogenic Yeast Candida glabrata. Prog. Mol. Subcell. Biol. 2019, 58, 155–193. [Google Scholar] [CrossRef]
- West, L.; Lowman, D.W.; Mora-Montes, H.M.; Grubb, S.; Murdoch, C.; Thornhill, M.H.; Gow, N.A.R.; Williams, D.; Haynes, K. Differential Virulence of Candida glabrata Glycosylation Mutants. J. Biol. Chem. 2013, 288, 22006–22018. [Google Scholar] [CrossRef] [PubMed]
- Briones-Martin-Del-Campo, M.; Orta-Zavalza, E.; Juarez-Cepeda, J.; Gutierrez-Escobedo, G.; Cañas-Villamar, I.; Castaño, I.; De Las Peñas, A. The Oxidative Stress Response of the Opportunistic Fungal Pathogen Candida glabrata. Rev. Iberoam. Micol. 2014, 31, 67–71. [Google Scholar] [CrossRef]
- Riedelberger, M.; Penninger, P.; Tscherner, M.; Hadriga, B.; Brunnhofer, C.; Jenull, S.; Stoiber, A.; Bourgeois, C.; Petryshyn, A.; Glaser, W.; et al. Type I Interferons Ameliorate Zinc Intoxication of Candida glabrata by Macrophages and Promote Fungal Immune Evasion. iScience 2020, 23, 101121. [Google Scholar] [CrossRef]
- Angoulvant, A.; Guitard, J.; Hennequin, C. Old and New Pathogenic Nakaseomyces Species: Epidemiology, Biology, Identification, Pathogenicity and Antifungal Resistance. FEMS Yeast Res. 2015, 16, fov114. [Google Scholar] [CrossRef]
- Köhler, J.R.; Acosta-Zaldívar, M.; Qi, W. Phosphate in Virulence of Candida albicans and Candida glabrata. J. Fungi 2020, 6, 40. [Google Scholar] [CrossRef]
- Gutiérrez-Escobedo, G.; Hernández-Carreón, O.; Morales-Rojano, B.; Revuelta-Rodríguez, B.; Vázquez-Franco, N.; Castaño, I.; De Las Peñas, A. Candida glabrata Peroxiredoxins, Tsa1 and Tsa2, and Sulfiredoxin, Srx1, Protect against Oxidative Damage and Are Necessary for Virulence. Fungal Genet. Biol. 2020, 135, 103287. [Google Scholar] [CrossRef]
- Martínez-Pastor, M.T.; Puig, S. Adaptation to Iron Deficiency in Human Pathogenic Fungi. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118797. [Google Scholar] [CrossRef] [PubMed]
- Seider, K.; Brunke, S.; Schild, L.; Jablonowski, N.; Wilson, D.; Majer, O.; Barz, D.; Haas, A.; Kuchler, K.; Schaller, M.; et al. The Facultative Intracellular Pathogen Candida glabrata Subverts Macrophage Cytokine Production and Phagolysosome Maturation. J. Immunol. 2011, 187, 3072–3086. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Fox, E.P.; Nett, J.E.; Sorrells, T.R.; Mitrovich, Q.M.; Hernday, A.D.; Tuch, B.B.; Andes, D.R.; Johnson, A.D. A Recently Evolved Transcriptional Network Controls Biofilm Development in Candida albicans. Cell 2012, 148, 126–138. [Google Scholar] [CrossRef]
- Beyer, R.; Jandric, Z.; Zutz, C.; Gregori, C.; Willinger, B.; Jacobsen, I.D.; Kovarik, P.; Strauss, J.; Schüller, C. Competition of Candida glabrata against Lactobacillus Is Hog1 Dependent. Cell. Microbiol. 2018, 20, e12943. [Google Scholar] [CrossRef]
- De Las Peñas, A.; Juárez-Cepeda, J.; López-Fuentes, E.; Briones-Martín-del-Campo, M.; Gutiérrez-Escobedo, G.; Castaño, I. Local and Regional Chromatin Silencing in Candida glabrata: Consequences for Adhesion and the Response to Stress. FEMS Yeast Res. 2015, 15, fov056. [Google Scholar] [CrossRef]
- Rai, M.N.; Sharma, V.; Balusu, S.; Kaur, R. An Essential Role for Phosphatidylinositol 3-Kinase in the Inhibition of Phagosomal Maturation, Intracellular Survival and Virulence in Candida glabrata. Cell. Microbiol. 2015, 17, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive Candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef]
- Riedelberger, M.; Penninger, P.; Tscherner, M.; Seifert, M.; Jenull, S.; Brunnhofer, C.; Scheidl, B.; Tsymala, I.; Bourgeois, C.; Petryshyn, A.; et al. Type I Interferon Response Dysregulates Host Iron Homeostasis and Enhances Candida glabrata Infection. Cell Host Microbe 2020, 27, 454–466.e8. [Google Scholar] [CrossRef]
- Rasheed, M.; Battu, A.; Kaur, R. Aspartyl Proteases in Candida glabrata are Required for Suppression of the Host Innate Immune Response. J. Biol. Chem. 2018, 293, 6410–6433. [Google Scholar] [CrossRef]
- Gabaldón, T.; Gómez-Molero, E.; Bader, O. Molecular Typing of Candida glabrata. Mycopathologia 2020, 185, 755–764. [Google Scholar] [CrossRef]
- He, C.; Song, Y.; Chang, X.K. Pathogenicity and drug-resistance analysis of Candida glabrata in patients with oral candidiasis. Zhonghua Yi Xue Za Zhi 2020, 100, 1778–1782. [Google Scholar] [CrossRef]
- Cai, S.; Xu, J.; Shao, Y.; Gong, J.; Zhao, F.; He, L.; Shan, X. Rapid Identification of the Candida glabrata Species Complex by High-Resolution Melting Curve Analysis. J. Clin. Lab. Anal. 2020, 34, e23226. [Google Scholar] [CrossRef] [PubMed]
- Smyth, J.; Mullen, C.C.; Jack, L.; Collier, A.; Bal, A.M. Diabetes, Malignancy and Age as Predictors of Candida glabrata Bloodstream Infection: A Re-Evaluation of the Risk Factors. J. Mycol. Med. 2018, 28, 547–550. [Google Scholar] [CrossRef]
- Rosenwald, A.G.; Arora, G.; Ferrandino, R.; Gerace, E.L.; Mohammednetej, M.; Nosair, W.; Rattila, S.; Subic, A.Z.; Rolfes, R. Identification of Genes in Candida glabrata Conferring Altered Responses to Caspofungin, a Cell Wall Synthesis Inhibitor. G3 Genes Genomes Genet. 2016, 6, 2893–2907. [Google Scholar] [CrossRef]
- Subramanya, S.H.; Baral, B.P.; Sharan, N.K.; Nayak, N.; Metok, Y.; Sathian, B.; Bairy, I.; Gokhale, S. Antifungal Susceptibility and Phenotypic Virulence Markers of Candida Species Isolated from Nepal. BMC Res. Notes 2017, 10, 543. [Google Scholar] [CrossRef] [PubMed]
- Márquez, F.; Iturrieta, I.; Calvo, M.; Urrutia, M.; Godoy-Martínez, P. Epidemiología y Susceptibilidad Antifúngica de Especies Causantes de Candidemia En La Ciudad de Valdivia, Chile. Rev. Chil. Infectol. 2017, 34, 441–446. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rasheed, M.; Battu, A.; Kaur, R. Host-Pathogen Interaction in Candida glabrata Infection: Current Knowledge and Implications for Antifungal Therapy. Expert Rev. Anti-Infect. Ther. 2020, 18, 1093–1103. [Google Scholar] [CrossRef]
- Borman, A.M.; Muller, J.; Walsh-Quantick, J.; Szekely, A.; Patterson, Z.; Palmer, M.D.; Fraser, M.; Johnson, E.M. Fluconazole Resistance in Isolates of Uncommon Pathogenic Yeast Species from the United Kingdom. Antimicrob. Agents Chemother. 2019, 63, e00211-19. [Google Scholar] [CrossRef] [PubMed]
- Mashaly, G.; Shrief, R. Candida glabrata Complex from Patients with Healthcare-Associated Infections in Mansoura University Hospitals, Egypt: Distribution, Antifungal Susceptibility and Effect of Fluconazole and Polymyxin B Combination. Germs 2019, 9, 125–132. [Google Scholar] [CrossRef]
- Miranda-Cadena, K.; Marcos-Arias, C.; Mateo, E.; Aguirre, J.M.; Quindós, G.; Eraso, E. Prevalence and Antifungal Susceptibility Profiles of Candida glabrata, Candida parapsilosis and Their Close-Related Species in Oral Candidiasis. Arch. Oral Biol. 2018, 95, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Marín Martínez, E.M.; Aller García, A.I.; Martín-Mazuelos, E. Epidemiology, risk factors and in vitro susceptibility in candidaemia due to non-Candida albicans species. Rev. Iberoam. Micol. 2016, 33, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.R.; Jimenez Ortigosa, C.; Shor, E.; Perlin, D.S. Genetic Drivers of Multidrug Resistance in Candida glabrata. Front. Microbiol. 2016, 7, 1995. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xia, R.; Zhang, Q.; Bai, C.; Li, Z.; Zhang, P. Evaluation of Candidemia in Epidemiology and Risk Factors among Cancer Patients in a Cancer Center of China: An 8-Year Case-Control Study. BMC Infect. Dis. 2017, 17, 536. [Google Scholar] [CrossRef] [PubMed]
- Ngouana, T.K.; Toghueo, R.M.K.; Kenfack, I.F.; Lachaud, L.; Nana, A.K.; Tadjou, L.; Kouanfack, C.; Boyom, F.F.; Bertout, S. Epidemiology and Antifungal Susceptibility Testing of Non-albicans Candida Species Colonizing Mucosae of HIV-Infected Patients in Yaoundé (Cameroon). J. Mycol. Med. 2019, 29, 233–238. [Google Scholar] [CrossRef]
- Waikhom, S.D.; Afeke, I.; Kwawu, G.S.; Mbroh, H.K.; Osei, G.Y.; Louis, B.; Deku, J.G.; Kasu, E.S.; Mensah, P.; Agede, C.Y.; et al. Prevalence of Vulvovaginal Candidiasis among Pregnant Women in the Ho Municipality, Ghana: Species Identification and Antifungal Susceptibility of Candida Isolates. BMC Pregnancy Childbirth 2020, 20, 266. [Google Scholar] [CrossRef]
- Arastehfar, A.; Daneshnia, F.; Salehi, M.-R.; Zarrinfar, H.; Khodavaisy, S.; Haas, P.-J.; Roudbary, M.; Najafzadeh, M.-J.; Zomorodian, K.; Charsizadeh, A.; et al. Molecular Characterization and Antifungal Susceptibility Testing of Candida nivariensis from Blood Samples—An Iranian Multicentre Study and a Review of the Literature. J. Med. Microbiol. 2019, 68, 770–777. [Google Scholar] [CrossRef]
- Alobaid, K.; Khan, Z. Epidemiologic Characteristics of Adult Candidemic Patients in a Secondary Hospital in Kuwait: A Retrospective Study. J. Mycol. Med. 2019, 29, 35–38. [Google Scholar] [CrossRef]
- Al-Baqsami, Z.F.; Ahmad, S.; Khan, Z. Antifungal Drug Susceptibility, Molecular Basis of Resistance to Echinocandins and Molecular Epidemiology of Fluconazole Resistance among Clinical Candida glabrata Isolates in Kuwait. Sci. Rep. 2020, 10, 6238. [Google Scholar] [CrossRef]
- Gülmez, D.; Alp, S.; Gursoy, G.; Ayaz, C.M.; Dogan, O.; Arikan-Akdagli, S.; Akova, M. Mixed Fungaemia: An 18-Year Report from a Tertiary-Care University Hospital and a Systematic Review. Clin. Microbiol. Infect. 2020, 26, 833–841. [Google Scholar] [CrossRef]
- Aldejohann, A.M.; Herz, M.; Martin, R.; Walther, G.; Kurzai, O. Emergence of Resistant Candida glabrata in Germany. JAC Antimicrob. Resist. 2021, 3, dlab122. [Google Scholar] [CrossRef]
- Coste, A.T.; Kritikos, A.; Li, J.; Khanna, N.; Goldenberger, D.; Garzoni, C.; Zehnder, C.; Boggian, K.; Neofytos, D.; Riat, A.; et al. Emerging Echinocandin-Resistant Candida albicans and glabrata in Switzerland. Infection 2020, 48, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.; Borman, A.M.; Thorn, R.; Lawrance, L.M. Resistance to Echinocandin Antifungal Agents in the United Kingdom in Clinical Isolates of Candida glabrata: Fifteen Years of Interpretation and Assessment. Med. Mycol. 2020, 58, 219–226. [Google Scholar] [CrossRef]
- Szweda, P.; Gucwa, K.; Romanowska, E.; Dzierz Anowska-Fangrat, K.; Naumiuk, Ł.; Brillowska-Da Browska, A.; Wojciechowska-Koszko, I.; Milewski, S. Mechanisms of Azole Resistance among Clinical Isolates of Candida glabrata in Poland. J. Med. Microbiol. 2015, 64, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Medina, N.; Soto-Debrán, J.C.; Seidel, D.; Akyar, I.; Badali, H.; Barac, A.; Bretagne, S.; Cag, Y.; Cassagne, C.; Castro, C.; et al. MixInYeast: A multicenter study on mixed yeast infections. J. Fungi 2021, 7, 13. [Google Scholar] [CrossRef]
- Hull, C.M.; Parker, J.E.; Bader, O.; Weig, M.; Gross, U.; Warrilow, A.G.S.; Kelly, D.E.; Kelly, S.L. Facultative Sterol Uptake in an Ergosterol-Deficient Clinical Isolate of Candida glabrata Harboring a Missense Mutation in ERG11 and Exhibiting Cross-Resistance to Azoles and Amphotericin B. Antimicrob. Agents Chemother. 2012, 56, 4223–4232. [Google Scholar] [CrossRef] [PubMed]
- Colombo, A.L.; Júnior, J.N.; Guinea, J. Emerging Multidrug-Resistant Candida Species. Curr. Opin. Infect. Dis. 2017, 30, 528–538. [Google Scholar] [CrossRef]
- Montagna, M.T.; Lovero, G.; Coretti, C.; Martinelli, D.; De Giglio, O.; Iatta, R.; Balbino, S.; Rosato, A.; Caggiano, G. Susceptibility to Echinocandins of Candida spp. Strains Isolated in Italy Assessed by European Committee for Antimicrobial Susceptibility Testing and Clinical Laboratory Standards Institute Broth Microdilution Methods. BMC Microbiol. 2015, 15, 106. [Google Scholar] [CrossRef] [PubMed]
- Vale-Silva, L.; Beaudoing, E.; Tran, V.D.T.; Sanglard, D. Comparative Genomics of Two Sequential Candida glabrata Clinical Isolates. G3 Genes Genomes Genet. 2017, 7, 2413–2426. [Google Scholar] [CrossRef] [PubMed]
- Simonicova, L.; Moye-Rowley, W.S. Functional Information from Clinically-Derived Drug Resistant Forms of the Candida glabrata Pdr1 Transcription Factor. PLoS Genet. 2020, 16, e1009005. [Google Scholar] [CrossRef]
- López-Soria, L.M.; Bereciartua, E.; Santamaría, M.; Soria, L.M.; Hernández-Almaraz, J.L.; Mularoni, A.; Nieto, J.; Montejo, M. First case report of catheter-related fungemia by Candida nivariensis in the Iberian Peninsula. Rev. Iberoam. Micol. 2013, 30, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.; Selmecki, A.; Forche, A.; Diogo, D.; Bougnoux, M.-E.; d’Enfert, C.; Berman, J.; Sanglard, D. Genotypic evolution of azole resistance mechanisms in sequential Candida albicans isolates. Eukaryot. Cell 2007, 6, 1889–1904. [Google Scholar] [CrossRef]
- Morio, F.; Pagniez, F.; Besse, M.; Gay-andrieu, F.; Miegeville, M.; Pape, P.L. Deciphering azole resistance mechanisms with a focus on transcription factor-encoding genes TAC1, MRR1 and UPC2 in a set of fluconazole-resistant clinical isolates of Candida albicans. Int. J. Antimicrob. Agents 2013, 42, 410–415. [Google Scholar] [CrossRef]
- Nagayoshi, Y.; Miyazaki, T.; Shimamura, S.; Nakayama, H.; Minematsu, A.; Yamauchi, S.; Takazono, T.; Nakamura, S.; Yanagihara, K.; Kohno, S.; et al. Unexpected Effects of Azole Transporter Inhibitors on Antifungal Susceptibility in Candida glabrata and Other Pathogenic Candida Species. PLoS ONE 2017, 12, e0180990. [Google Scholar] [CrossRef]
- Hou, X.; Xiao, M.; Wang, H.; Yu, S.-Y.; Zhang, G.; Zhao, Y.; Xu, Y.-C. Profiling of PDR1 and MSH2 in Candida glabrata Bloodstream Isolates from a Multicenter Study in China. Antimicrob. Agents Chemother. 2018, 62, e00153-18. [Google Scholar] [CrossRef]
- Culakova, H.; Dzugasova, V.; Perzelova, J.; Gbelska, Y.; Subik, J. Mutation of the CgPDR16 Gene Attenuates Azole Tolerance and Biofilm Production in Pathogenic Candida glabrata. Yeast 2013, 30, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Farahyar, S.; Zaini, F.; Kordbacheh, P.; Rezaie, S.; Falahati, M.; Safara, M.; Raoofian, R.; Hatami, K.; Mohebbi, M.; Heidari, M. Expression of Efflux Pumps and Fatty Acid Activator One Genes in Azole Resistant Candida glabrata Isolated from Immunocompromised Patients. Acta Med. Iran. 2016, 54, 458–464. [Google Scholar]
- Yu, S.-J.; Chang, Y.-L.; Chen, Y.-L. Deletion of ADA2 Increases Antifungal Drug Susceptibility and Virulence in Candida glabrata. Antimicrob. Agents Chemother. 2018, 62, e01924-17. [Google Scholar] [CrossRef] [PubMed]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.D.; Johnson, M.D.; Pfeiffer, C.D.; Jiménez-Ortigosa, C.; Catania, J.; Booker, R.; Castanheira, M.; Messer, S.A.; Perlin, D.S.; Pfaller, M.A. Increasing Echinocandin Resistance in Candida glabrata: Clinical Failure Correlates with Presence of FKS Mutations and Elevated Minimum Inhibitory Concentrations. Clin. Infect. Dis. 2013, 56, 1724–1732. [Google Scholar] [CrossRef] [PubMed]
- Duxbury, S.J.N.; Bates, S.; Beardmore, R.E.; Gudelj, I. Evolution of Drug-Resistant and Virulent Small Colonies in Phenotypically Diverse Populations of the Human Fungal Pathogen Candida glabrata. Proc. Biol. Sci. 2020, 287, 761. [Google Scholar] [CrossRef]
- Bordallo-Cardona, M.Á.; Marcos-Zambrano, L.J.; Sánchez-Carrillo, C.; de la Pedrosa, E.G.G.; Cantón, R.; Bouza, E.; Escribano, P.; Guinea, J. Mutant Prevention Concentration and Mutant Selection Window of Micafungin and Anidulafungin in Clinical Candida glabrata Isolates. Antimicrob. Agents Chemother. 2018, 62, e01982-17. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Arendrup, M.; Cantón, E.; Cordoba, S.; Dannaoui, E.; García-Rodríguez, J.; Gonzalez, G.M.; Govender, N.P.; Martin-Mazuelos, E.; Lackner, M.; et al. Multicenter Study of Method-Dependent Epidemiological Cutoff Values for Detection of Resistance in Candida spp. and Aspergillus spp. to Amphotericin B and Echinocandins for the Etest Agar Diffusion Method. Antimicrob. Agents Chemother. 2017, 61, e01792-16. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Alastruey-Izquierdo, A.; Healey, K.R.; Johnson, M.E.; Perlin, D.S.; Edlind, T.D. Fks1 and Fks2 Are Functionally Redundant but Differentially Regulated in Candida glabrata: Implications for Echinocandin Resistance. Antimicrob. Agents Chemother. 2012, 56, 6304–6309. [Google Scholar] [CrossRef]
- Vandeputte, P.; Tronchin, G.; Larcher, G.; Ernoult, E.; Bergès, T.; Chabasse, D.; Bouchara, J.P. A nonsense mutation in the ERG6 gene leads to reduced susceptibility to polyenes in a clinical isolate of Candida glabrata. Antimicrob. Agents Chemother. 2008, 52, 3701–3709. [Google Scholar] [CrossRef] [PubMed]
- Minematsu, A.; Miyazaki, T.; Shimamura, S.; Nishikawa, H.; Nakayama, H.; Takazono, T.; Saijo, T.; Yamamoto, K.; Imamura, Y.; Yanagihara, K.; et al. Vacuolar Proton-Translocating ATPase Is Required for Antifungal Resistance and Virulence of Candida glabrata. PLoS ONE 2019, 14, e0210883. [Google Scholar] [CrossRef]
- Roetzer, A.; Gregori, C.; Jennings, A.M.; Quintin, J.; Ferrandon, D.; Butler, G.; Kuchler, K.; Ammerer, G.; Schüller, C. Candida glabrata Environmental Stress Response Involves Saccharomyces cerevisiae Msn2/4 Orthologous Transcription Factors. Mol. Microbiol. 2008, 69, 603–620. [Google Scholar] [CrossRef] [PubMed]
- Fujita, S.; Senda, Y.; Okusi, T.; Ota, Y.; Takada, H.; Yamada, K.; Kawano, M. Catheter-related fungemia due to fluconazole-resistant Candida nivariensis. J. Clin. Microbiol. 2007, 45, 3459–3461. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Ma, B.; Cormack, B.P. A family of glycosylphosphatidylinositol-linked aspartyl proteases is required for virulence of Candida glabrata. Proc. Natl. Acad. Sci. USA 2007, 104, 7628–7633. [Google Scholar] [CrossRef]
- Sikora, M.; Dabkowska, M.; Swoboda-Kopec, E.; Jarzynka, S.; Netsvyetayeva, I.; Jaworska-Zaremba, M.; Pertkiewicz, M.; Mlynarczyk, G. Differences in Proteolytic Activity and Gene Profiles of Fungal Strains Isolated from the Total Parenteral Nutrition Patients. Folia Microbiol. 2011, 56, 143–148. [Google Scholar] [CrossRef]
- Cuenca-Estrella, M.; Gomez-Lopez, A.; Isla, G.; Rodriguez, D.; Almirante, B.; Pahissa, A.; Rodriguez-Tudela, J.L. Barcelona Candidemia Project Study Group Prevalence of Candida bracarensis and Candida nivariensis in a Spanish Collection of Yeasts: Comparison of Results from a Reference Centre and from a Population-Based Surveillance Study of Candidemia. Med. Mycol. 2011, 49, 525–529. [Google Scholar] [CrossRef]
- Galocha, M.; Pais, P.; Cavalheiro, M.; Pereira, D.; Viana, R.; Teixeira, M.C. Divergent Approaches to Virulence in C. albicans and C. glabrata: Two Sides of the Same Coin. Int. J. Mol. Sci. 2019, 20, 2345. [Google Scholar] [CrossRef]
- de Melo Riceto, É.B.; de Paula Menezes, R.; Penatti, M.P.; dos Santos Pedroso, R. Enzymatic and Hemolytic Activity in Different Candida Species. Rev. Iberoam. Micol. 2015, 32, 79–82. [Google Scholar] [CrossRef]
- Mohandas, V.; Ballal, M. Distribution of Candida Species in Different Clinical Samples and Their Virulence: Biofilm Formation, Proteinase and Phospholipase Production: A Study on Hospitalized Patients in Southern India. J. Glob. Infect. Dis. 2011, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Dabiri, S.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Comparative Analysis of Proteinase, Phospholipase, Hydrophobicity and Biofilm Forming Ability in Candida Species Isolated from Clinical Specimens. J. Mycol. Med. 2018, 28, 437–442. [Google Scholar] [CrossRef]
- Kalaiarasan, K.; Singh, R.; Chaturvedula, L. Changing Virulence Factors among Vaginal Non-albicans Candida Species. Indian J. Med. Microbiol. 2018, 36, 364–368. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, Epidemiology, Pathogenicity and Antifungal Resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef] [PubMed]
- Berila, N.; Hyroššová, P.; Šubík, J. Oxidative Stress Response and Virulence Factors in Candida glabrata Clinical Isolates. Folia Microbiol. 2011, 56, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Atalay, M.A.; Koc, A.N.; Demir, G.; Sav, H. Investigation of Possible Virulence Factors in Candida Strains Isolated from Blood Cultures. Niger. J. Clin. Pract. 2015, 18, 52–55. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.; Kastora, S.L.; Gomes-Gonçalves, A.; Azevedo, N.; Rodrigues, C.F.; Silva, S.; Demuyser, L.; Van Dijck, P.; Casal, M.; Brown, A.J.P.; et al. Transcriptional Responses of Candida glabrata Biofilm Cells to Fluconazole Are Modulated by the Carbon Source. npj Biofilms Microbiomes 2020, 6, 4. [Google Scholar] [CrossRef]
- Tam, P.; Gee, K.; Piechocinski, M.; Macreadie, I. Candida glabrata, Friend and Foe. J. Fungi 2015, 1, 277–292. [Google Scholar] [CrossRef]
- Kalkanci, A.; GüZel, A.B.; Khalil, I.I.J.; Aydin, M.; Ilkit, M.; Kuştimur, S. Yeast Vaginitis during Pregnancy: Susceptibility Testing of 13 Antifungal Drugs and Boric Acid and the Detection of Four Virulence Factors. Med. Mycol. 2012, 50, 585–593. [Google Scholar] [CrossRef]
- Alves, C.T.; Wei, X.-Q.; Silva, S.; Azeredo, J.; Henriques, M.; Williams, D.W. Candida albicans Promotes Invasion and Colonisation of Candida glabrata in a Reconstituted Human Vaginal Epithelium. J. Infect. 2014, 69, 396–407. [Google Scholar] [CrossRef]
- Valotteau, C.; Prystopiuk, V.; Cormack, B.P.; Dufrêne, Y.F. Atomic Force Microscopy Demonstrates That Candida glabrata Uses Three Epa Proteins To Mediate Adhesion to Abiotic Surfaces. mSphere 2019, 4, e00277-19. [Google Scholar] [CrossRef]
- Brunke, S.; Hube, B. Two Unlike Cousins: Candida albicans and C. glabrata Infection Strategies. Cell. Microbiol. 2013, 15, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Pärnänen, P.; Meurman, J.H.; Nikula-Ijäs, P. A Novel Candida glabrata Cell Wall Associated Serine Protease. Biochem. Biophys. Res. Commun. 2015, 457, 676–680. [Google Scholar] [CrossRef]
- Juárez-Cepeda, J.; Orta-Zavalza, E.; Cañas-Villamar, I.; Arreola-Gómez, J.; Pérez-Cornejo, G.P.; Hernández-Carballo, C.Y.; Gutiérrez-Escobedo, G.; Castaño, I.; De Las Peñas, A. The EPA2 Adhesin Encoding Gene Is Responsive to Oxidative Stress in the Opportunistic Fungal Pathogen Candida glabrata. Curr. Genet. 2015, 61, 529–544. [Google Scholar] [CrossRef]
- Rodrigues, C.; Rodrigues, M.; Silva, S.; Henriques, M. Candida glabrata Biofilms: How Far Have We Come? J. Fungi 2017, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- López-Fuentes, E.; Gutiérrez-Escobedo, G.; Timmermans, B.; Van Dijck, P.; De Las Peñas, A.; Castaño, I. Candida glabrata’s Genome Plasticity Confers a Unique Pattern of Expressed Cell Wall Proteins. J. Fungi 2018, 4, 67. [Google Scholar] [CrossRef]
- Llopis-Torregrosa, V.; Vaz, C.; Monteoliva, L.; Ryman, K.; Engstrom, Y.; Gacser, A.; Gil, C.; Ljungdahl, P.O.; Sychrová, H. Trk1-Mediated Potassium Uptake Contributes to Cell-Surface Properties and Virulence of Candida glabrata. Sci. Rep. 2019, 9, 7529. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Molero, E.; de Boer, A.D.; Dekker, H.L.; Moreno-Martínez, A.; Kraneveld, E.A.; Ichsan; Chauhan, N.; Weig, M.; de Soet, J.J.; de Koster, C.G.; et al. Proteomic Analysis of Hyperadhesive Candida glabrata Clinical Isolates Reveals a Core Wall Proteome and Differential Incorporation of Adhesins. FEMS Yeast Res. 2015, 15, fov098. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Gabaldón, T.; Schüller, C. From Saccharomyces Cerevisiae to Candida glabrata in a Few Easy Steps: Important Adaptations for an Opportunistic Pathogen. FEMS Microbiol. Lett. 2011, 314, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gabaldón, T.; Martin, T.; Marcet-Houben, M.; Durrens, P.; Bolotin-Fukuhara, M.; Lespinet, O.; Arnaise, S.; Boisnard, S.; Aguileta, G.; Atanasova, R.; et al. Comparative Genomics of Emerging Pathogens in the Candida glabrata Clade. BMC Genom. 2013, 14, 623. [Google Scholar] [CrossRef]
- Olson, M.L.; Jayaraman, A.; Kao, K.C. Relative Abundances of Candida albicans and Candida glabrata in in vitro Coculture Biofilms Impact Biofilm Structure and Formation. Appl. Environ. Microbiol. 2018, 84, e02769-17. [Google Scholar] [CrossRef]
- Cortés-Acosta, E.; Ibarra, J.A.; Ramírez-Saad, H.; Vargas-Mendoza, C.F.; Villa-Tanaca, L.; Hernández-Rodríguez, C. Polymorphism in the Regulatory Regions of Genes CgYPS1 and CgYPS7 Encoding Yapsins in Candida glabrata Is Associated with Changes in Expression Levels. FEMS Yeast Res. 2017, 17, fox077. [Google Scholar] [CrossRef][Green Version]
- Muñoz-Duarte, A.R.; Castrejón-Jiménez, N.S.; Baltierra-Uribe, S.L.; Pérez-Rangel, S.J.; Carapia-Minero, N.; Castañeda-Sánchez, J.I.; Luna-Herrera, J.; López-Santiago, R.; Rodríguez-Tovar, A.V.; García-Pérez, B.E. Candida glabrata Survives and Replicates in Human Osteoblasts. Pathog. Dis. 2016, 74, ftw030. [Google Scholar] [CrossRef]
- López-Fuentes, E.; Hernández-Hernández, G.; Castanedo, L.; Gutiérrez-Escobedo, G.; Oktaba, K.; De las Peñas, A.; Castaño, I. Chromatin Loop Formation Induced by a Subtelomeric Protosilencer Represses EPA Genes in Candida glabrata. Genetics 2018, 210, 113–128. [Google Scholar] [CrossRef]
- Desai, C.; Mavrianos, J.; Chauhan, N. Candida glabrata Pwp7p and Aed1p are Required for Adherence to Human Endothelial Cells. FEMS Yeast Res. 2011, 11, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo-Carvalho, M.H.; Ramos, L.D.; Barbedo, L.S.; Chaves, A.L.; Muramoto, I.A.; Santos, A.L.; Almeida-Paes, R.; Zancopé-Oliveira, R.M. First Description of Candida nivariensis in Brazil: Antifungal Susceptibility Profile and Potential Virulence Attributes. Mem. Inst. Oswaldo Cruz 2016, 111, 51–58. [Google Scholar] [CrossRef]
- Luo, G.; Samaranayake, L.P. Candida glabrata, an emerging fungal pathogen, exhibits superior relative cell surface hydrophobicity and adhesion to denture acrylic surfaces compared with Candida albicans. Apmis 2002, 110, 601–610. [Google Scholar] [CrossRef]
- Negri, M.; Gonçalves, V.; Silva, S.; Henriques, M.; Azeredo, J.; Oliveira, R. Crystal Violet Staining to Quantify Candida Adhesion to Epithelial Cells. Br. J. Biomed. Sci. 2010, 67, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Vieira de Melo, A.P.; Zuza-Alves, D.L.; da Silva-Rocha, W.P.; Ferreira Canário de Souza, L.B.; Francisco, E.C.; Salles de Azevedo Melo, A.; Maranhão Chaves, G. Virulence Factors of Candida spp. Obtained from Blood Cultures of Patients with Candidemia Attended at Tertiary Hospitals in Northeast Brazil. J. Mycol. Med. 2019, 29, 132–139. [Google Scholar] [CrossRef]
- Mutlu Sariguzel, F.; Berk, E.; Koc, A.N.; Sav, H.; Demir, G. Investigation of the Relationship between Virulence Factors and Genotype of Candida spp. Isolated from Blood Cultures. J. Infect. Dev. Ctries 2015, 9, 857–864. [Google Scholar] [CrossRef]
- Moreira, A.; Silva, S.; Botelho, C.; Sampaio, P.; Pais, C.; Henriques, M. Candida bracarensis: Evaluation of Virulence Factors and Its Tolerance to Amphotericin B and Fluconazole. Mycopathologia 2015, 180, 305–315. [Google Scholar] [CrossRef]
- Bairwa, G.; Rasheed, M.; Taigwal, R.; Sahoo, R.; Kaur, R. GPI (Glycosylphosphatidylinositol)-Linked Aspartyl Proteases Regulate Vacuole Homoeostasis in Candida glabrata. Biochem. J. 2014, 458, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Rapala-Kozik, M.; Bochenska, O.; Zajac, D.; Karkowska-Kuleta, J.; Gogol, M.; Zawrotniak, M.; Kozik, A. Extracellular Proteinases of Candida Species Pathogenic Yeasts. Mol. Oral Microbiol. 2018, 33, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Yagmur, G.; Sav, H.; Ziyade, N.; Elgormus, N.; Sen, S.; Akkoyun Bilgi, E.; Atan, Y.; Buyuk, Y.; Kiraz, N. Evaluation of Virulence Factors and Antifungal Susceptibility in Yeast Isolates from Postmortem Specimens. J. Forensic Sci. 2016, 61, 1000–1006. [Google Scholar] [CrossRef] [PubMed]
- Rossoni, R.D.; Barbosa, J.O.; Vilela, S.F.G.; dos Santos, J.D.; Jorge, A.O.C.; Junqueira, J.C. Correlation of Phospholipase and Proteinase Production of Candida with in vivo Pathogenicity in Galleria mellonella. Braz. J. Oral Sci. 2013, 12, 199–204. [Google Scholar] [CrossRef]
- Hacioglu, M.; Guzel, C.B.; Savage, P.B.; Tan, A.S.B. Antifungal Susceptibilities, in vitro Production of Virulence Factors and Activities of Ceragenins against Candida spp. Isolated from Vulvovaginal Candidiasis. Med. Mycol. 2019, 57, 291–299. [Google Scholar] [CrossRef]
- Sriphannam, C.; Nuanmuang, N.; Saengsawang, K.; Amornthipayawong, D.; Kummasook, A. Anti-Fungal Susceptibility and Virulence Factors of Candida spp. Isolated from Blood Cultures. J. Mycol. Med. 2019, 29, 325–330. [Google Scholar] [CrossRef]
- Barbosa, A.H.; Damasceno, J.L.; Casemiro, L.A.; Martins, C.H.G.; Pires, R.H.; Candido, R.C. Susceptibility to Oral Antiseptics and Virulence Factors Ex Vivo Associated with Candida spp. Isolated from Dental Prostheses. J. Prosthodont. 2019, 28, 398–408. [Google Scholar] [CrossRef]
- Pereira, C.A.; Domingues, N.; Araújo, M.I.D.S.A.; Junqueira, J.C.; Back-Brito, G.N.; Jorge, A.O.C. Production of Virulence Factors in Candida Strains Isolated from Patients with Denture Stomatitis and Control Individuals. Diagn. Microbiol. Infect. Dis. 2016, 85, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Tay, S.T.; Lotfalikhani, A.; Sabet, N.S.; Ponnampalavanar, S.; Sulaiman, S.; Na, S.L.; Ng, K.P. Occurrence and Characterization of Candida nivariensis from a Culture Collection of Candida glabrata Clinical Isolates in Malaysia. Mycopathologia 2014, 178, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Schaller, M.; Borelli, C.; Korting, H.C.; Hube, B. Hydrolytic enzymes as virulence factors of Candida albicans. Mycoses 2005, 48, 365–367. [Google Scholar] [CrossRef]
- Marcos-Arias, C.; Eraso, E.; Madariaga, L.; Aguirre, J.M.; Quindós, G. Phospholipase and Proteinase Activities of Candida Isolates from Denture Wearers. Mycoses 2011, 54, e10–e16. [Google Scholar] [CrossRef] [PubMed]
- Kumari, V.; Banerjee, T.; Kumar, P.; Pandey, S.; Tilak, R. Emergence of Non-albicans Candida among Candidal Vulvovaginitis Cases and Study of Their Potential Virulence Factors, from a Tertiary Care Center, North India. Indian J. Pathol. Microbiol. 2013, 56, 144–147. [Google Scholar] [CrossRef]
- Salari, S.; Sadat Seddighi, N.; Ghasemi Nejad Almani, P. Evaluation of Biofilm Formation Ability in Different Candida Strains and Anti-Biofilm Effects of Fe3O4-NPs Compared with Fluconazole: An in vitro Study. J. Mycol. Med. 2018, 28, 23–28. [Google Scholar] [CrossRef]
- Tellapragada, C.; Eshwara, V.K.; Johar, R.; Shaw, T.; Malik, N.; Bhat, P.V.; Kamath, A.; Mukhopadhyay, C. Antifungal Susceptibility Patterns, in vitro Production of Virulence Factors, and Evaluation of Diagnostic Modalities for the Speciation of Pathogenic Candida from Blood Stream Infections and Vulvovaginal Candidiasis. J. Pathog. 2014, 2014, 142864. [Google Scholar] [CrossRef]
- Furlaneto, M.C.; Góes, H.P.; Perini, H.F.; Dos Santos, R.C.; Furlaneto-Maia, L. How Much Do We Know about Hemolytic Capability of Pathogenic Candida Species? Folia Microbiol. 2018, 63, 405–412. [Google Scholar] [CrossRef]
- Luo, G.; Samaranayake, L.P.; Cheung, B.P.K.; Tang, G. Reverse transcriptase polymerase chain reaction (RT-PCR) detection of HLP gene expression in Candida glabrata and its possible role in in vitro haemolysin production. Apmis 2004, 112, 283–290. [Google Scholar] [CrossRef]
- Nevitt, T.; Thiele, D.J. Host Iron Withholding Demands Siderophore Utilization for Candida glabrata to Survive Macrophage Killing. PLoS Pathog. 2011, 7, e1001322. [Google Scholar] [CrossRef] [PubMed]
- Araújo, D.; Henriques, M.; Silva, S. Portrait of Candida Species Biofilm Regulatory Network Genes. Trends Microbiol. 2017, 25, 62–75. [Google Scholar] [CrossRef]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, R.; Li, Y.; Wang, Z.; Ishchuk, O.P.; Ahmad, K.M.; Wee, J.; Piskur, J.; Shapiro, J.A.; Gu, Z. Understand the Genomic Diversity and Evolution of Fungal Pathogen Candida glabrata by Genome-Wide Analysis of Genetic Variations. Methods 2020, 176, 82–90. [Google Scholar] [CrossRef]
- Pannanusorn, S.; Fernandez, V.; Römling, U. Prevalence of Biofilm Formation in Clinical Isolates of Candida Species Causing Bloodstream Infection. Mycoses 2013, 56, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Kunyeit, L.; Kurrey, N.K.; Anu-Appaiah, K.A.; Rao, R.P. Probiotic Yeasts Inhibit Virulence of Non-albicans Candida Species. mBio 2019, 10, e02307-19. [Google Scholar] [CrossRef]
- Cataldi, V.; Di Campli, E.; Fazii, P.; Traini, T.; Cellini, L.; Di Giulio, M. Candida Species Isolated from Different Body Sites and Their Antifungal Susceptibility Pattern: Cross-Analysis of Candida albicans and Candida glabrata Biofilms. Med. Mycol. 2017, 55, 624–634. [Google Scholar] [CrossRef]
- Pongrácz, J.; Benedek, K.; Juhász, E.; Iván, M.; Kristóf, K. In vitro Biofilm Production of Candida Bloodstream Isolates: Any Association with Clinical Characteristics? J. Med. Microbiol. 2016, 65, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Kraneveld, E.A.; de Soet, J.J.; Deng, D.M.; Dekker, H.L.; de Koster, C.G.; Klis, F.M.; Crielaard, W.; de Groot, P.W.J. Identification and Differential Gene Expression of Adhesin-like Wall Proteins in Candida glabrata Biofilms. Mycopathologia 2011, 172, 415–427. [Google Scholar] [CrossRef]
- de Groot, P.W.J.; Kraneveld, E.A.; Yin, Q.Y.; Dekker, H.L.; Gross, U.; Crielaard, W.; de Koster, C.G.; Bader, O.; Klis, F.M.; Weig, M. The cell wall of the human pathogen Candida glabrata: Differential incorporation of novel adhesin-like wall proteins. Eukaryot. Cell 2008, 7, 1951–1964. [Google Scholar] [CrossRef]
- Leiva-Peláez, O.; Gutiérrez-Escobedo, G.; López-Fuentes, E.; Cruz-Mora, J.; De Las Peñas, A.; Castaño, I. Molecular Characterization of the Silencing Complex SIR in Candida glabrata Hyperadherent Clinical Isolates. Fungal Genet. Biol. 2018, 118, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Costa, C.; Mil-Homens, D.; Romão, D.; de Carvalho, C.C.C.R.; Pais, P.; Mira, N.P.; Fialho, A.M.; Teixeira, M.C. The Multidrug Resistance Transporters CgTpo1_1 and CgTpo1_2 Play a Role in Virulence and Biofilm Formation in the Human Pathogen Candida glabrata. Cell. Microbiol. 2017, 19, e12686. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Cavalheiro, M.; Costa, C.; Takahashi-Nakaguchi, A.; Okamoto, M.; Chibana, H.; Teixeira, M.C. Screening the Drug:H+ Antiporter Family for a Role in Biofilm Formation in Candida glabrata. Front. Cell. Infect. Microbiol. 2020, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K.; Bhakt, P.; Kaur, R. Essential Role for the Phosphatidylinositol 3,5-Bisphosphate Synthesis Complex in Caspofungin Tolerance and Virulence in Candida glabrata. Antimicrob. Agents Chemother. 2019, 63, e00886-19. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Fernandes, L.; Henriques, M.; Silva, S. Environmental PH Modulates Biofilm Formation and Matrix Composition in Candida albicans and Candida glabrata. Biofouling 2020, 36, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Azevedo, N.M.; Henriques, M.; Silva, S. Hormones Modulate Candida Vaginal Isolates Biofilm Formation and Decrease Their Susceptibility to Azoles and Hydrogen Peroxide. Med. Mycol. 2020, 58, 341–350. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.; Azeredo, J. Silicone Colonization by Non-Candida albicans Candida Species in the Presence of Urine. J. Med. Microbiol. 2010, 59, 747–754. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Continent | Country | Number of Strains | Susceptibility and Resistance | References | |||
|---|---|---|---|---|---|---|---|
| Azoles | Echinocandins | Polyenes | Other | ||||
| Africa | Ethiopia | 17 | Clotrimazole R: 11.76% Itraconazole: R: 76.47% Ketoconazole: R: 41.18% | [14] | |||
| Cameroon | 33 | Fluconazole: R: 3.03% | [104] | ||||
| Ghana | 31 | Fluconazole: S: 19.4% ISDD: 41.9% R: 38.7% Voriconazole: S: 54.8% ISDD: 9.7% R: 35.5% | Nystatin ISDD: 12.9% R: 87.1% | [105] | |||
| America | Canada | 392 | Fluconazole: ISDD: 87.8% | [37] | |||
| Chile | 37 | Fluconazole: R: 6.6% Itraconazole R: 20% | Micafungin R: 10% | Amphotericin B R: 2.7% | [16] | ||
| Brazil | 38 Susceptibility tests were performed in 8 | Fluconazole R: 50% Miconazole R: 12.5% | Amphotericin B S: 100% Nystatin S: 100% | [51] | |||
| Asia | China | 73 | Fluconazole R: 6.8% Voriconazole R: 6.8% | Amphotericin B R: 100% 5-fluorocytosisin: R: 11% | [20] | ||
| India | 21 | Fluconazole S: 100% Itraconazole S: 100% Posaconazole S: 100% Ketoconazole S: 100% Voriconazole S: 100% | Caspofungin S: 100% | Amphotericin B S: 100% 5-fluorocytosisin S: 100% | [21] | ||
| India | 22 | High Resistance to clotrimazole, fluconazole, itraconazole, ketoconazole: 100% | High Resistance to nystatin: 100% | [70] | |||
| Iran | 4 (C. nivariensis) | Fluconazole S: 100% Itraconazole S: 100% Voriconazole S: 100% Posaconazole S: 100% | Micafungin S: 100% | Amphotericin B S: 100% 5-fluorocytosine S: 100% | [106] | ||
| Nepal | 9 | Fluconazole S: 66.6% ISDD: 11.1% R: 22.2% Voriconazole: ISDD: 88.8% R: 11.1% | Caspofungin S: 100% | Amphotericin B ISDD: 100% | [95] | ||
| Kuwait | 11 | Fluconazole: R: 64% ISDD: 36% | [107] | ||||
| Kuwait | 75 | Fluconazole: R: 48% ISDD: 52% | Micafungin: R: 4% ISDD: 2.67% Caspofungin: R: 5.33% | Amphotericin B R: 6.67% | [108] | ||
| Turkey | 12 | Fluconazole R: 100% | [109] | ||||
| Turkey | 83 | Fluconazole: R: 9.2% Itraconazole: R: 45.8% Voriconazole R: 43.4% | [71] | ||||
| Europe | Spain | 14 | Fluconazole: ISDD: 100% Itraconazole: R: 50% Voriconazole: S: 100% | Caspofungin: S: 92.9% ISDD: 7.1% Micafungin: S: 100% Anidulafungin: S: 85.7% ISDD: 14.3% | Amphotericin B S: 100% 5-fluorocytosisin: S: 100% | [101] | |
| Ireland | 21 | Fluconazole: R: 37% | Amphotericin B: R: 14% | [5] | |||
| Poland | 445 | Fluconazole: ISDD: 100% Itraconazole: R: 41% Posaconazole: R: 50% Voriconazole: S: 83% | Caspofungin, anidulafungin and micafungin: S: 100% | Amphotericin B: S: 100% | [57] | ||
| Greece | 34 | Fluconazole: ISDD: 94% R: 6% | Anidulafungin: S: 97% R: 3% Caspofungin S: 88% ISDD: 3% R: 9% Micafungin S: 97% R: 3% | [24] | |||
| Germany | 176 | Fluconazole: R: 38% | Anidulafungin: S: 52% R: 48% | Combined resistance to fluconazole and echinocandins: 14% | [110] | ||
| Switzerland | 5 | Fluconazole: ISDD: 100% | Caspofungin, anidulafungin and micafungin R: 100% | [111] | |||
| United Kingdom | 7225 | Caspofungin, anidulafungin and micafungin: R: 0.55% | [112] | ||||
| Poland | 81 | Fluconazole: R: 22.2% ISDD: 77.7% Fluconazole and voriconazole: R: 1.2% Voriconazole R: 7.4% Cross-Resistance to other azoles: R: 18.5% | Caspofungin, anidulafungin and micafungin: S: 100% | Amphotericin B: S: 100% 5-fluorocytosine: S: 93.8% ISDD: 3.7% R: 2.5% | [113] | ||
| Poland, France, Greece, Germany, Italy, Czech Republic, Spain, Austria, Serbia, Iran, India, Thailand, United States | 64 | Fluconazole: R: 1.6% Itraconazole: R: 1.6% Isavuconazole S: 100% Posaconazole: R: 3.1% Voriconazole R: 3.1% | Caspofungin, anidulafungin and micafungin S: 100% | Amphotericin B: S: 100% 5-fluorocytosine: S: 100% | [114] | ||
| Germany Spain | 4 1 | Fluconazole: R: 100% Voriconazole: R: 100% | Amphotericin B: R: 100% | [115] | |||
| Jerusalem | 176 | Fluconazole: ISDD: 81.25% R: 4% Voriconazole: R: 4.7% | Caspofungin: R: 33.6% | Amphotericin B S: 100% | [11] | ||
| Oceania | Australia | 35 | Fluconazole: R: 22.8% | Caspofungin: R: 17.1% | [3] | ||
| Yeast | Drug on Which Resistance Is Generated | Antifungal Resistance | References | |||
|---|---|---|---|---|---|---|
| Genes and Proteins Involved | Mechanisms Involved | Result | ||||
| Complex C. glabrata | Azoles | Multiple drugs | Mutations (polymorphisms, deletions, etc.) in the Pdr1 gene of the PDR1 transcription factor and TAC1 transcription factor | Overexpression and activation of ATP-binding cassettes [CDR1, CDR2 (also designated PDH1, SNQ2, FAA1)] | Drugs transported to the outside of the cell | [8,113,117,118,119, 120,121,122,123, 124,126] |
| Decreased cell surface hydrophobicity during biofilm formation | [125] | |||||
| Modification of biological transport pathways of hydrophobic compounds and lipid metabolism | [126] | |||||
| Azoles | Mitochondrial dysfunction associated with the development of mitochondrial DNA-deficient “small mutants.” | Drugs transported to the outside of the cell | [64,120,124] | |||
| Azoles | Deletion of CgADA2 | Positive regulation of adherence factors | Thermotolerance and hypervirulence | [127] | ||
| Echinocandins | Mutations in FKS1 and FKS2 in the hot spots HS1 and HS2 (Examples: FKS1 S629P, FKS2 F659 FKS2 S663P y) | Altered conformation of the 1,3-β-glucan-synthase subunits Fks1p and Fks2p | Reduced affinity of echinocandins for β-1,3 glucan. | [108,110,111,129,130,131,132,133] | ||
| Azoles/polyenes | Mutations in the ERG6 and ERG11 proteins | Alteration of sterol 14-α-demethylase | Less ergosterol content with cell membrane modification. | [134] | ||
| Ergosterol exchanged for generated exogenous sterols | [115] | |||||
| C. nivariensis | Azoles | Increased expression of YPS1, AWP3, EPA1, ERG11, CDR1, and CDR2 genes | Antifungal resistance and increased virulence | [86] | ||
| Increased mRNA expression of ERG11, CDR1, and CDR2 | Overexpression and activation of CDRs | Increased antifungal resistance | [102] | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frías-De-León, M.G.; Hernández-Castro, R.; Conde-Cuevas, E.; García-Coronel, I.H.; Vázquez-Aceituno, V.A.; Soriano-Ursúa, M.A.; Farfán-García, E.D.; Ocharán-Hernández, E.; Rodríguez-Cerdeira, C.; Arenas, R.; et al. Candida glabrata Antifungal Resistance and Virulence Factors, a Perfect Pathogenic Combination. Pharmaceutics 2021, 13, 1529. https://doi.org/10.3390/pharmaceutics13101529
Frías-De-León MG, Hernández-Castro R, Conde-Cuevas E, García-Coronel IH, Vázquez-Aceituno VA, Soriano-Ursúa MA, Farfán-García ED, Ocharán-Hernández E, Rodríguez-Cerdeira C, Arenas R, et al. Candida glabrata Antifungal Resistance and Virulence Factors, a Perfect Pathogenic Combination. Pharmaceutics. 2021; 13(10):1529. https://doi.org/10.3390/pharmaceutics13101529
Chicago/Turabian StyleFrías-De-León, María Guadalupe, Rigoberto Hernández-Castro, Esther Conde-Cuevas, Itzel H. García-Coronel, Víctor Alfonso Vázquez-Aceituno, Marvin A. Soriano-Ursúa, Eunice D. Farfán-García, Esther Ocharán-Hernández, Carmen Rodríguez-Cerdeira, Roberto Arenas, and et al. 2021. "Candida glabrata Antifungal Resistance and Virulence Factors, a Perfect Pathogenic Combination" Pharmaceutics 13, no. 10: 1529. https://doi.org/10.3390/pharmaceutics13101529
APA StyleFrías-De-León, M. G., Hernández-Castro, R., Conde-Cuevas, E., García-Coronel, I. H., Vázquez-Aceituno, V. A., Soriano-Ursúa, M. A., Farfán-García, E. D., Ocharán-Hernández, E., Rodríguez-Cerdeira, C., Arenas, R., Robledo-Cayetano, M., Ramírez-Lozada, T., Meza-Meneses, P., Pinto-Almazán, R., & Martínez-Herrera, E. (2021). Candida glabrata Antifungal Resistance and Virulence Factors, a Perfect Pathogenic Combination. Pharmaceutics, 13(10), 1529. https://doi.org/10.3390/pharmaceutics13101529