Evaluation of CpG-ODN-Adjuvanted Toxoplasma gondii Virus-Like Particle Vaccine upon One, Two, and Three Immunizations


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mice and Parasite
2.2. VLP Vaccine and Reagents
2.3. Immunization and T. gondii ME49 Infection
2.4. Sample Preparation
2.5. ELISA for Antibody Responses and Cytokine Levels
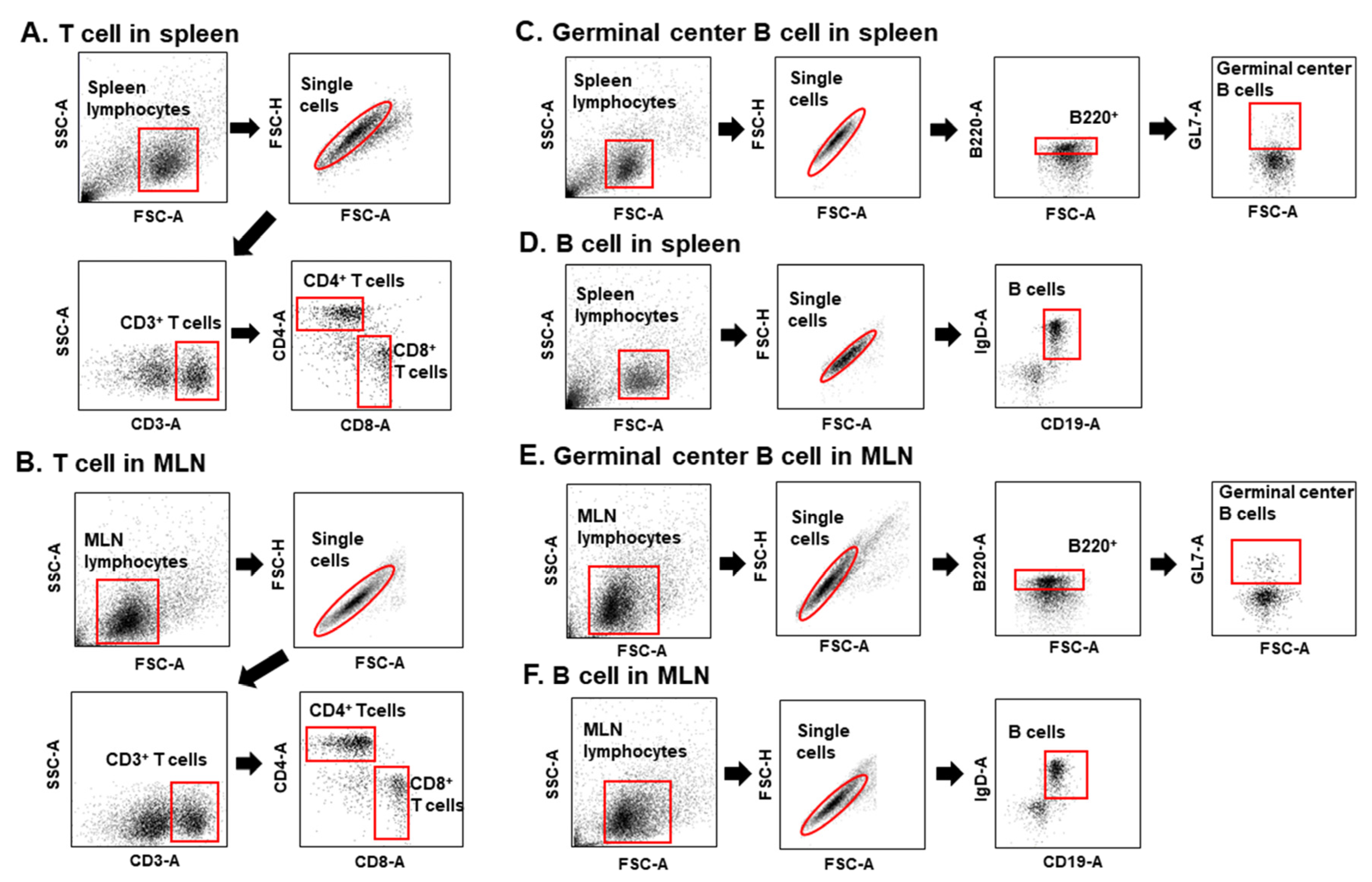
2.6. Flow Cytometry
2.7. Statistical Analysis
3. Results
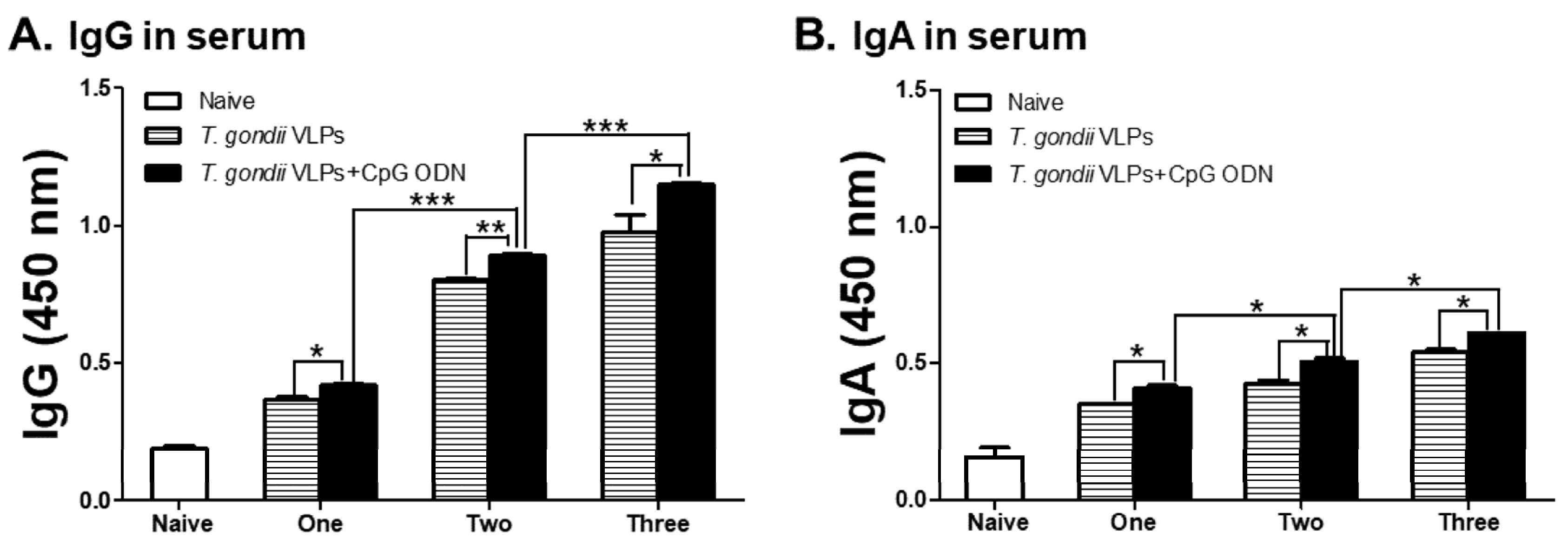
3.1. T. gondii-Specific IgG and IgA Antibody Responses in Sera
3.2. T. gondii—Specific IgG and IgA Antibody Responses in Mucosal Samples
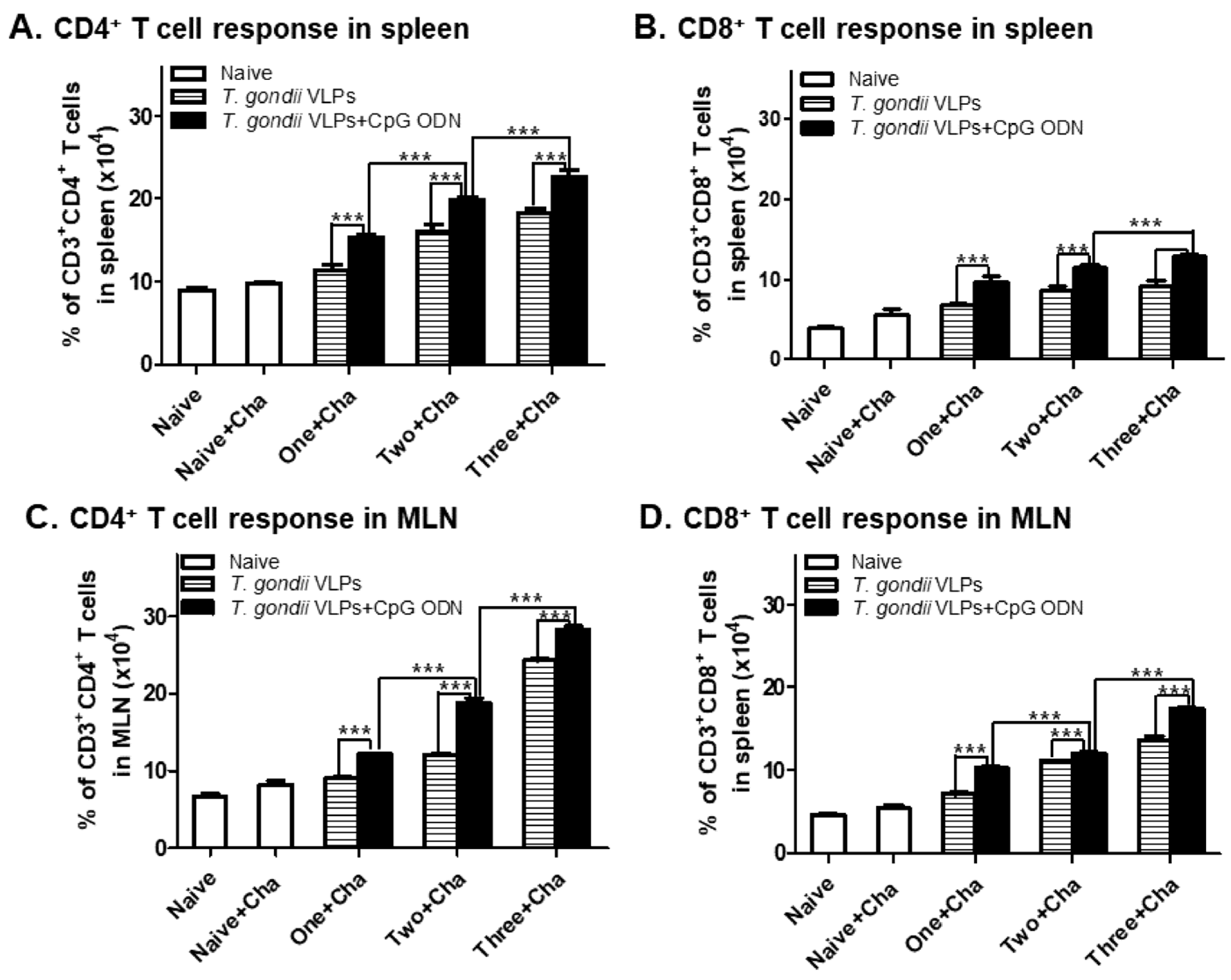
3.3. Immunization with T. gondii VLPs Alone or Adjuvanted VLPs Induces CD4+ and CD8+ T-Cell Expansion
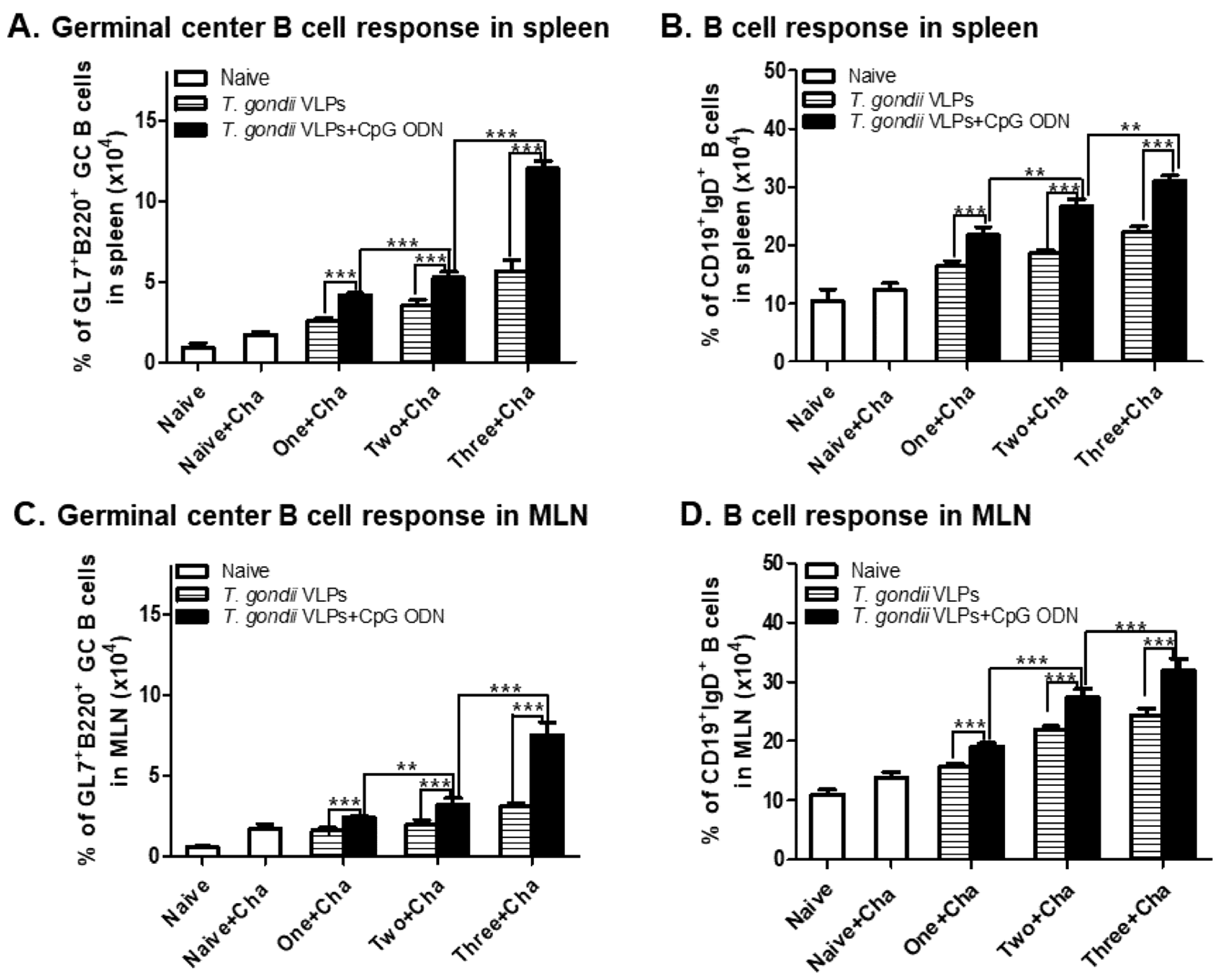
3.4. CpG-ODN-Adjuvanted VLP Vaccination Induced Germinal Center (GC) B-Cell and B-Cell Responses
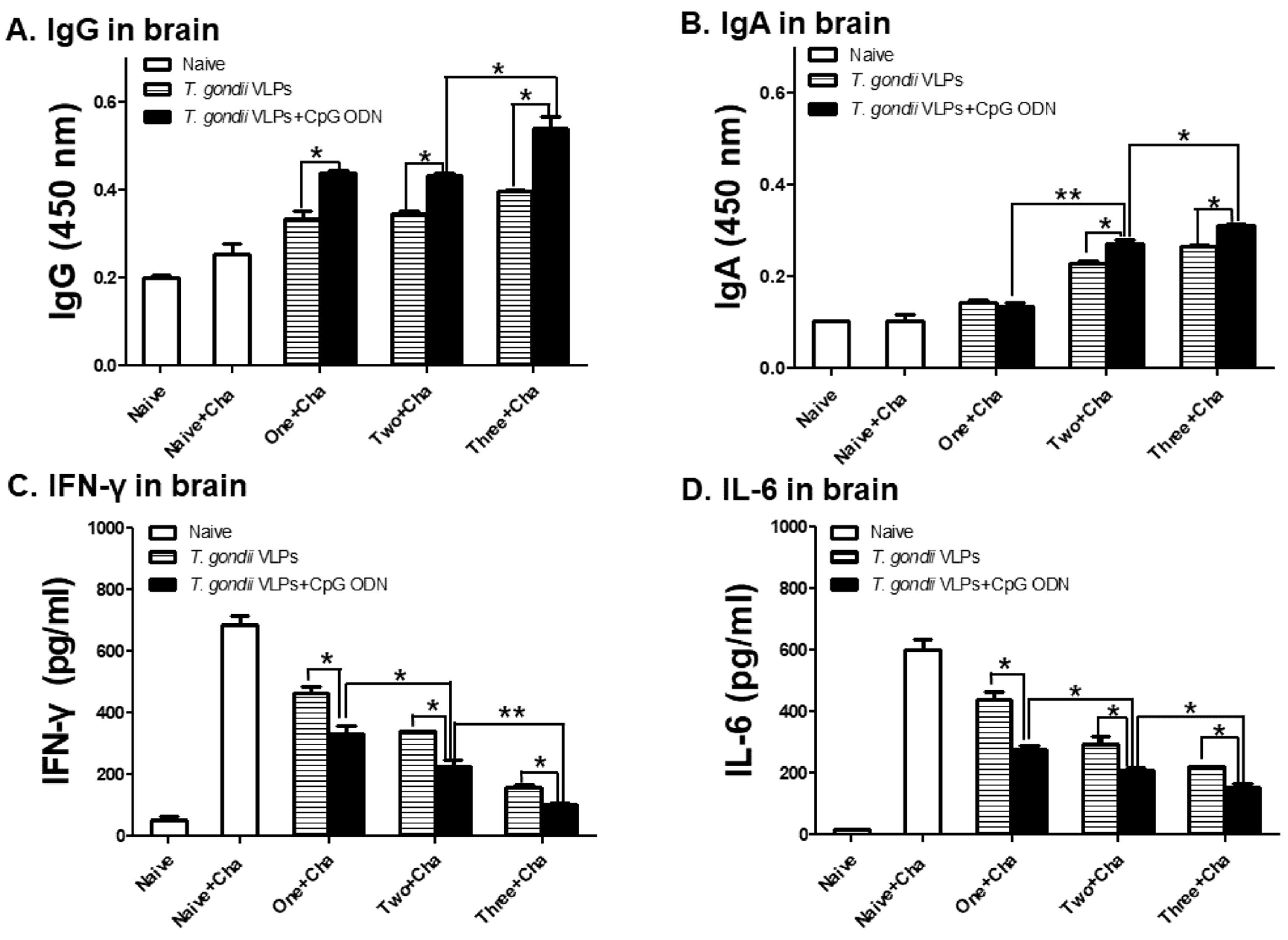
3.5. Antibody and Pro-Inflammatory Cytokine Responses in the Brain
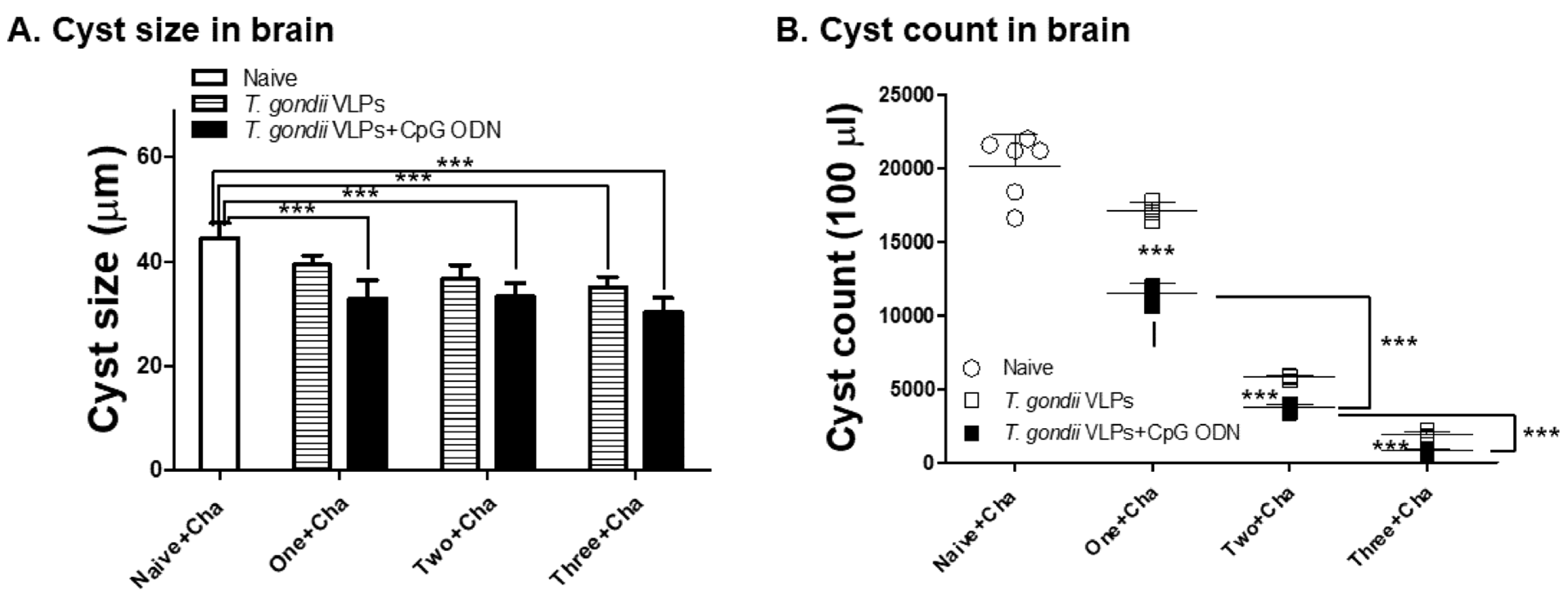
3.6. Adjuvanted T. gondii VLP Vaccination Significantly Reduced Cyst Counts upon Challenge Infection with T. gondii ME49
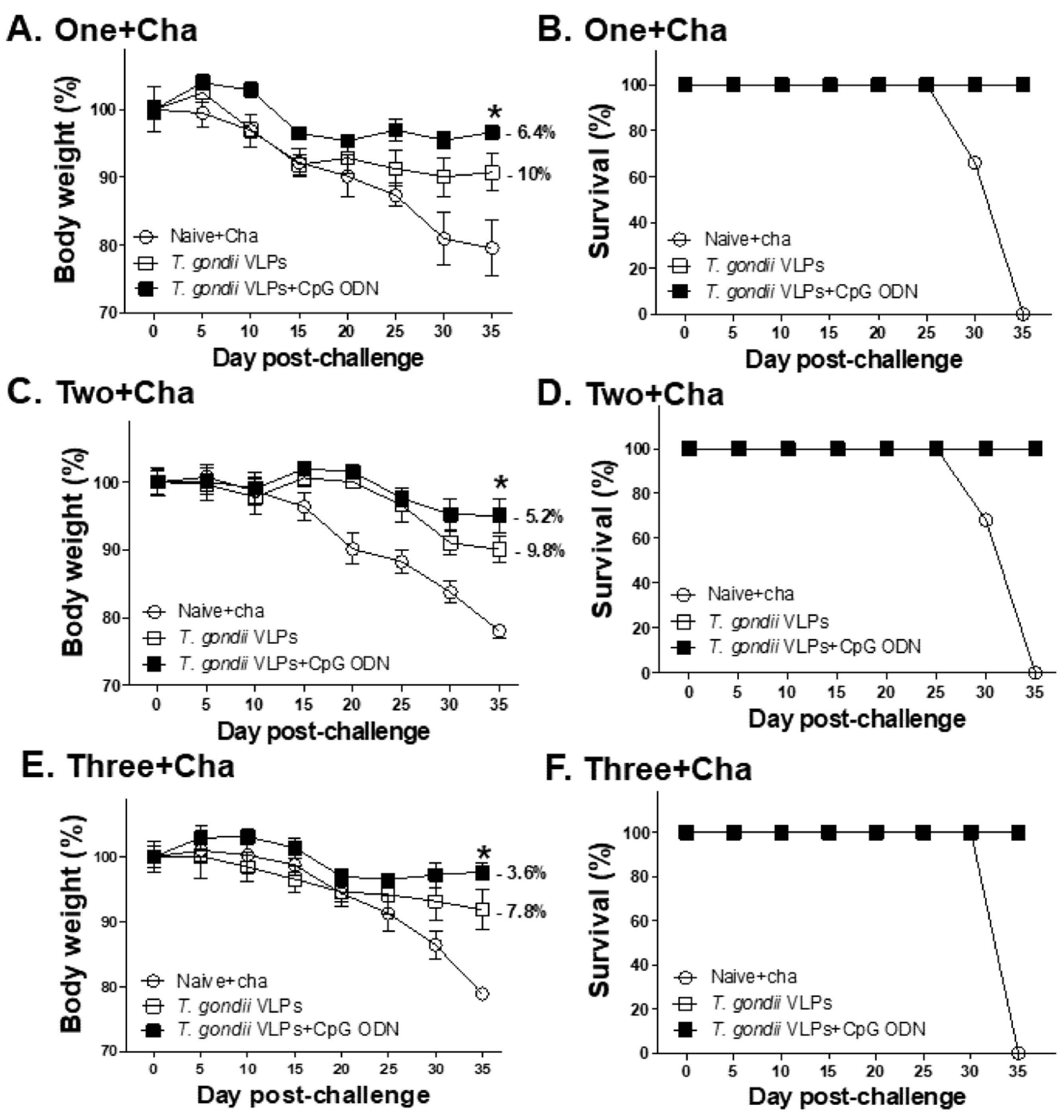
3.7. Protective Efficacy Induced by T. gondii VLPs with or without Adjuvant CpG-ODN
4. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Black, M.W.; Boothroyd, J.C. Lytic Cycle of Toxoplasma gondii. Microbiol. Mol. Biol. Rev. 2000, 64, 607–623. [Google Scholar] [CrossRef]
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From Animals to Humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef]
- Dubey, J.P. The history and life cycle of Toxoplasma gondii. In Toxoplasma gondii; Elsevier: London, UK, 2020; pp. 1–19. [Google Scholar] [CrossRef]
- Winstanley, P. Drug Treatment of Toxoplasmic Encephalitis in Acquired Immunodeficiency Syndrome. Postgrad. Med. J. 1995, 71, 404–408. [Google Scholar] [CrossRef][Green Version]
- McCabe, R.; Chirurgi, V. Issues in Toxoplasmosis. Infect. Dis. Clin. N. Am. 1993, 7, 587. [Google Scholar] [PubMed]
- Alday, P.H.; Doggett, J.S. Drugs in Development for Toxoplasmosis: Advances, Challenges, and Current Status. Drug Des. Dev. Ther. 2017, 11, 273–293. [Google Scholar] [CrossRef] [PubMed]
- Lanjewar, D.N.; Surve, K.V.; Maheshwari, M.B.; Shenoy, B.P.; Hira, S.K. Toxoplasmosis of the Central Nervous System in the Acquired Immunodeficiency Syndrome. Indian J. Pathol. Microbiol. 1998, 41, 147–151. [Google Scholar]
- Martins-Duarte, E.S.; Urbina, J.A.; de Souza, W.; Vommaro, R.C. Antiproliferative Activities of Two Novel Quinuclidine Inhibitors against Toxoplasma gondii Tachyzoites In Vitro. J. Antimicrob. Chemother. 2006, 58, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Singla, L.D.; Zhou, H. Vaccines against Toxoplasma gondii: Status, Challenges and Future Directions. Hum. Vaccines Immunother. 2012, 8, 1305–1308. [Google Scholar] [CrossRef]
- Angus, C.; Klivington-Evans, D.; Dubey, J.; Kovacs, J.A. Immunization with a DNA Plasmid Encoding the SAG1 (P30) Protein of Toxoplasma gondii is Immunogenic and Protective in Rodents. J. Infect. Dis. 2000, 181, 317–324. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, A.; Li, Y.; Li, X.; Cong, H.; He, S.; Zhou, H. Immunization with a DNA Vaccine Encoding Toxoplasma gondii Superoxide Dismutase (TgSOD) Induces Partial Immune Protection Against Acute Toxoplasmosis in BALB/C Mice. BMC Infect. Dis. 2017, 17, 403. [Google Scholar] [CrossRef]
- Parmley, S.; Slifer, T.; Araujo, F. Protective Effects of Immunization with a Recombinant Cyst Antigen in Mouse Models of Infection with Toxoplasma gondii Tissue Cysts. J. Infect. Dis. 2002, 185, S90–S95. [Google Scholar] [CrossRef] [PubMed]
- Cuppari, A.F.; Sanchez, V.; Ledesma, B.; Frank, F.M.; Goldman, A.; Angel, S.O.; Martin, V. Toxoplasma gondii Protease Inhibitor-1 (TgPI-1) is a Novel Vaccine Candidate Against Toxoplasmosis. Vaccine 2008, 26, 5040–5045. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, E.V.; Bernardes, E.S.; Silva, N.M.; Mineo, J.R.; Panunto-Castelo, A.; Roque-Barreira, M. Immunization with MIC1 and MIC4 Induces Protective Immunity against Toxoplasma gondii. Microb. Infect. 2006, 8, 1244–1251. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Zhou, T.; Liang, X.; Ye, S.; Zhao, P.; Yang, J.; Zhou, Y.; Zhao, J.; Shen, B. A Lactate Fermentation Mutant of Toxoplasma Stimulates Protective Immunity against Acute and Chronic Toxoplasmosis. Front. Immunol. 2018, 9, 1814. [Google Scholar] [CrossRef]
- Gigley, J.P.; Fox, B.A.; Bzik, D.J. Long-Term Immunity to Lethal Acute Or Chronic Type II Toxoplasma gondii Infection is Effectively Induced in Genetically Susceptible C57BL/6 Mice by Immunization with an Attenuated Type I Vaccine Strain. Infect. Immun. 2009, 77, 5380–5388. [Google Scholar] [CrossRef]
- Kang, H.; Lee, S.; Chu, K.; Lee, D.; Quan, F. Virus-Like Particles Expressing Toxoplasma gondii Rhoptry Protein 18 Induces Better Protection Than Rhoptry Protein 4 against, T. gondii Infection. Korean J. Parasitol. 2018, 56, 429. [Google Scholar] [CrossRef]
- Lee, S.H.; Chu, K.B.; Kang, H.J.; Quan, F.S. Virus-Like Particles Containing Multiple Antigenic Proteins of Toxoplasma gondii Induce Memory T Cell and B Cell Responses. PLoS ONE 2019, 14, e0220865. [Google Scholar] [CrossRef]
- Lee, S.; Kang, H.; Lee, D.; Kang, S.; Quan, F. Virus-Like Particle Vaccines Expressing Toxoplasma gondii Rhoptry Protein 18 and Microneme Protein 8 Provide Enhanced Protection. Vaccine 2018, 36, 5692–5700. [Google Scholar] [CrossRef]
- Lee, S.; Kang, H.; Lee, D.; Quan, F. Protective Immunity Induced by Incorporating Multiple Antigenic Proteins of Toxoplasma gondii into Influenza Virus-Like Particles. Front. Immunol. 2018, 9, 3073. [Google Scholar] [CrossRef]
- Li, T.; Wu, J.; Zhu, S.; Zang, G.; Li, S.; Lv, X.; Yue, W.; Qiao, Y.; Cui, J.; Shao, Y. A Novel C Type CpG Oligodeoxynucleotide Exhibits Immunostimulatory Activity in Vitro and Enhances Antitumor Effect in Vivo. Front. Immunol. 2020, 11, 8. [Google Scholar] [CrossRef]
- Vollmer, J.; Weeratna, R.; Payette, P.; Jurk, M.; Schetter, C.; Laucht, M.; Wader, T.; Tluk, S.; Liu, M.; Davis, H.L. Characterization of Three CpG Oligodeoxynucleotide Classes with Distinct Immunostimulatory Activities. Eur. J. Immunol. 2004, 34, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kim, A.; Lee, S.; Quan, F. Cross-Protection Induced by Toxoplasma gondii Virus-Like Particle Vaccine upon Intraperitoneal Route Challenge. Acta Trop. 2016, 164, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Lee, S.H.; Kim, A.R.; Quan, F.S. Virus-Like Nanoparticle Vaccine Confers Protection against Toxoplasma gondii. PLoS ONE 2016, 11, e0161231. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Chu, K.; Lee, S.; Kim, M.; Park, H.; Jin, H.; Moon, E.; Quan, F. Toxoplasma gondii Virus-Like Particle Vaccination Alleviates Inflammatory Response in the Brain upon T. gondii Infection. Parasite Immunol. 2020, 42, e12716. [Google Scholar] [CrossRef]
- Kang, H.; Lee, S.; Kim, M.; Chu, K.; Lee, D.; Chopra, M.; Choi, H.; Park, H.; Jin, H.; Quan, F. Influenza Virus-Like Particles Presenting both Toxoplasma gondii ROP4 and ROP13 Enhance Protection against, T. gondii Infection. Pharmaceutics 2019, 11, 342. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Nguyen, M.T. Recent Advances of Vaccine Adjuvants for Infectious Diseases. Immune Netw. 2015, 15, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Pasquale, A.D.; Preiss, S.; Silva, F.T.D.; Garçon, N. Vaccine Adjuvants: From 1920 to 2015 and Beyond. Vaccines 2015, 3, 320–343. [Google Scholar] [CrossRef]
- Kang, H.J.; Chu, K.B.; Lee, S.H.; Kim, M.J.; Park, H.; Jin, H.; Quan, F.S. Virus-Like Particle Vaccine Containing Toxoplasma gondii Rhoptry Protein 13 Induces Protection against, T. gondii ME49 Infection in Mice. Korean J. Parasitol. 2019, 57, 543–547. [Google Scholar] [CrossRef]
- Mathur, S.; Walley, K.R.; Boyd, J.H. The Toll-Like Receptor 9 Ligand CPG-C Attenuates Acute Inflammatory Cardiac Dysfunction. Shock 2011, 36, 478–483. [Google Scholar] [CrossRef]
- Klinman, D.M. CpG DNA as a Vaccine Adjuvant. Expert Rev. Vaccines 2003, 2, 305–315. [Google Scholar] [CrossRef]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine Adjuvants: Understanding the Structure and Mechanism of Adjuvanticity. Vaccine 2019, 37, 3167–3178. [Google Scholar] [CrossRef] [PubMed]
- Jongert, E.; Roberts, C.W.; Gargano, N.; Förster-Waldl, E.; Petersen, E. Vaccines Against Toxoplasma gondii: Challenges and Opportunities. Memórias do Instituto Oswaldo Cruz 2009, 104, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.A.; Smith, B.F.; Guarino, A.J.; Blagburn, B.L.; Baker, H.J. The use of CpG as an Adjuvant to Toxoplasma gondii Vaccination. Parasitol. Res. 2004, 92, 313–316. [Google Scholar] [CrossRef]
- EL-Malky, M.A.; Al-Harthi, S.A.; Mohamed, R.T.; Bali, M.A.E.; Saudy, N.S. Vaccination with Toxoplasma Lysate Antigen and CpG Oligodeoxynucleotides: Comparison of Immune Responses in Intranasal Versus Intramuscular Administrations. Parasitol. Res. 2014, 113, 2277–2284. [Google Scholar] [CrossRef]
- Liu, S.; Shi, L.; Cheng, Y.; Fan, G.; Ren, H.; Yuan, Y. Evaluation of Protective Effect of Multi-Epitope DNA Vaccine Encoding Six Antigen Segments of Toxoplasma gondii in Mice. Parasitol. Res. 2009, 105, 267. [Google Scholar] [CrossRef] [PubMed]
- Anis, E.; Moerman, L.; Ginsberg, G.; Karakis, I.; Slater, P.E.; Warshavsky, B.; Gosinov, R.; Grotto, I.; Marva, E. Did Two Booster Doses for Schoolchildren Change the Epidemiology of Pertussis in Israel? J. Public Health Policy 2018, 39, 304–317. [Google Scholar] [CrossRef]
- Ogra, P.L.; Faden, H.; Welliver, R.C. Vaccination Strategies for Mucosal Immune Responses. Clin. Microbiol. Rev. 2001, 14, 430–445. [Google Scholar] [CrossRef]
- Cohen, S.; Denkers, E. Border Maneuvers: Deployment of Mucosal Immune Defenses Against Toxoplasma gondii. Mucosal Immunol. 2014, 7, 744–752. [Google Scholar] [CrossRef]
- Innes, E.A.; Hamilton, C.; Garcia, J.L.; Chryssafidis, A.; Smith, D. A One Health Approach to Vaccines Against Toxoplasma gondii. Food Waterborne Parasitol. 2019, 15, e00053. [Google Scholar] [CrossRef]
- Kodama, S.; Abe, N.; Hirano, T.; Suzuki, M. Safety and Efficacy of Nasal Application of CpG Oligodeoxynucleotide as a Mucosal Adjuvant. Laryngoscope 2006, 116, 331–335. [Google Scholar] [CrossRef]
- Krieg, A.M.; Yi, A.; Matson, S.; Waldschmidt, T.J.; Bishop, G.A.; Teasdale, R.; Koretzky, G.A.; Klinman, D.M. CpG Motifs in Bacterial DNA Trigger Direct B-Cell Activation. Nature 1995, 374, 546–549. [Google Scholar] [CrossRef]
- Gallichan, W.S.; Woolstencroft, R.N.; Guarasci, T.; McCluskie, M.J.; Davis, H.L.; Rosenthal, K.L. Intranasal Immunization with CpG Oligodeoxynucleotides as an Adjuvant Dramatically Increases IgA and Protection Against Herpes Simplex Virus-2 in the Genital Tract. J. Immunol. 2001, 166, 3451–3457. [Google Scholar] [CrossRef] [PubMed]
- McCluskie, M.J.; Weeratna, R.D.; Payette, P.J.; Davis, H.L. Parenteral and Mucosal Prime-Boost Immunization Strategies in Mice with Hepatitis B Surface Antigen and CpG DNA. FEMS Immunol. Med. Microbiol. 2002, 32, 179–185. [Google Scholar] [CrossRef]
- Meek, B.; Klaren, V.N.; van Haeringen, N.J.; Kijlstra, A.; Peek, R. IgA Antibodies to Toxoplasma gondii in Human Tears. Invest. Ophthalmol. Vis. Sci. 2000, 41, 2584–2590. [Google Scholar]
- Jenum, P.A.; Stray-Pedersen, B. Development of Specific Immunoglobulins G, M, and A Following Primary Toxoplasma gondii Infection in Pregnant Women. J. Clin. Microbiol. 1998, 36, 2907–2913. [Google Scholar] [CrossRef] [PubMed]
- Ruddle, N.H.; Akirav, E.M. Secondary Lymphoid Organs: Responding to Genetic and Environmental Cues in Ontogeny and the Immune Response. J. Immunol. 2009, 183, 2205–2212. [Google Scholar] [CrossRef]
- Estato, V.; Stipursky, J.; Gomes, F.; Mergener, T.C.; Frazão-Teixeira, E.; Allodi, S.; Tibiriçá, E.; Barbosa, H.S.; Adesse, D. The Neurotropic Parasite Toxoplasma gondii Induces Sustained Neuroinflammation with Microvascular Dysfunction in Infected Mice. Am. J. Pathol. 2018, 188, 2674–2687. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.S.; Shin, J.; Yang, J.; Jung, B.; Lee, S.H.; Shin, E. Characteristics of Infection Immunity Regulated by Toxoplasma gondii to Maintain Chronic Infection in the Brain. Front. Immunol. 2018, 9, 158. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.-J.; Chu, K.-B.; Kim, M.-J.; Park, H.; Jin, H.; Lee, S.-H.; Moon, E.-K.; Quan, F.-S. Evaluation of CpG-ODN-Adjuvanted Toxoplasma gondii Virus-Like Particle Vaccine upon One, Two, and Three Immunizations. Pharmaceutics 2020, 12, 989. https://doi.org/10.3390/pharmaceutics12100989
Kang H-J, Chu K-B, Kim M-J, Park H, Jin H, Lee S-H, Moon E-K, Quan F-S. Evaluation of CpG-ODN-Adjuvanted Toxoplasma gondii Virus-Like Particle Vaccine upon One, Two, and Three Immunizations. Pharmaceutics. 2020; 12(10):989. https://doi.org/10.3390/pharmaceutics12100989
Chicago/Turabian StyleKang, Hae-Ji, Ki-Back Chu, Min-Ju Kim, Hyunwoo Park, Hui Jin, Su-Hwa Lee, Eun-Kyung Moon, and Fu-Shi Quan. 2020. "Evaluation of CpG-ODN-Adjuvanted Toxoplasma gondii Virus-Like Particle Vaccine upon One, Two, and Three Immunizations" Pharmaceutics 12, no. 10: 989. https://doi.org/10.3390/pharmaceutics12100989
APA StyleKang, H.-J., Chu, K.-B., Kim, M.-J., Park, H., Jin, H., Lee, S.-H., Moon, E.-K., & Quan, F.-S. (2020). Evaluation of CpG-ODN-Adjuvanted Toxoplasma gondii Virus-Like Particle Vaccine upon One, Two, and Three Immunizations. Pharmaceutics, 12(10), 989. https://doi.org/10.3390/pharmaceutics12100989