Aptamers as Delivery Agents of siRNA and Chimeric Formulations for the Treatment of Cancer

 ,
,
Abstract

1. Introduction
2. Aptamer in the Delivery of Therapeutic Nanoparticles Containing siRNA, shRNA, and miRNA
3. Cancer Immunotherapy Using Aptamers-siRNA
4. Tumor-Targeted Aptamers Complexed Directly to siRNAs and Other Agents for Cancer Therapy
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Singh, A.; Trivedi, P.; Jain, N.K. Advances in siRNA delivery in cancer therapy. Artif. Cells Nanomed. Biotechnol. 2018, 46, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Mishra, D.K.; Balekar, N.; Mishra, P.K. Nanoengineered strategies for siRNA delivery: From target assessment to cancer therapeutic efficacy. Drug Deliv. Transl. Res. 2017, 7, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.S. Improving cancer immunotherapy through nanotechnology. Nat. Rev. Cancer 2019, 19, 587–602. [Google Scholar] [CrossRef] [PubMed]
- Nanjwade, B.K.; Sarkar, A.B.; Srichana, T. Design and Characterization of Nanoparticulate Drug Delivery. In Characterization and Biology of Nanomaterials for Drug Delivery; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Wang, X.X.; Li, Y.B.; Yao, H.J.; Ju, R.J.; Zhang, Y.; Li, R.J.; Yu, Y.; Zhang, L.; Lu, W.L. The use of mitochondrial targeting resveratrol liposomes modified with a dequalinium polyethylene glycol-distearoylphosphatidyl ethanolamine conjugate to induce apoptosis in resistant lung cancer cells. Biomaterials 2011, 32, 5673–5687. [Google Scholar] [CrossRef] [PubMed]
- Faraji, A.H.; Wipf, P. Nanoparticles in cellular drug delivery. Bioorg. Med. Chem. 2009, 17, 2950–2962. [Google Scholar] [CrossRef]
- Bhushan, B. Springer Handbook of Nanotechnology; Bhushan, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 978-3-662-54355-9. [Google Scholar]
- Yao, V.J.; D’Angelo, S.; Butler, K.S.; Theron, C.; Smith, T.L.; Marchiò, S.; Gelovani, J.G.; Sidman, R.L.; Dobroff, A.S.; Brinker, C.J.; et al. Ligand-targeted theranostic nanomedicines against cancer. J. Control. Release 2016, 240, 267–286. [Google Scholar] [CrossRef] [PubMed]
- Guaragna, A.; Chiaviello, A.; Paolella, C.; DAlonzo, D.; Palumbo, G.; Palumbo, G. Synthesis and evaluation of folate-based chlorambucil delivery systems for tumor-targeted chemotherapy. Bioconjug. Chem. 2012, 23, 84–96. [Google Scholar] [CrossRef]
- Eloy, J.O.; Petrilli, R.; Raspantini, G.L.; Lee, R.J. Targeted Liposomes for siRNA Delivery to Cancer. Curr. Pharm. Des. 2018, 24, 2664–2672. [Google Scholar] [CrossRef]
- Kim, M.W.; Jeong, H.Y.; Kang, S.J.; Choi, M.J.; You, Y.M.; Im, C.S.; Lee, T.S.; Song, I.H.; Lee, C.G.; Rhee, K.J.; et al. Cancer-targeted Nucleic Acid Delivery and Quantum Dot Imaging Using EGF Receptor Aptamer-conjugated Lipid Nanoparticles. Sci. Rep. 2017, 7, 9474. [Google Scholar] [CrossRef]
- Guo, P.; Zhang, C.; Chen, C.; Garver, K.; Trottier, M. Inter-RNA interaction of phage θ29 pRNA to form a hexameric complex for viral DNA transportation. Mol. Cell 1998, 2, 149–155. [Google Scholar] [CrossRef]
- Hu, J.; Xiao, F.; Hao, X.; Bai, S.; Hao, J. Inhibition of monocyte adhesion to brain-derived endothelial cells by dual functional RNA chimeras. Mol. Ther. Nucleic Acids 2014, 3, e209. [Google Scholar] [CrossRef] [PubMed]
- Afonin, K.A.; Viard, M.; Koyfman, A.Y.; Martins, A.N.; Kasprzak, W.K.; Panigaj, M.; Desai, R.; Santhanam, A.; Grabow, W.W.; Jaeger, L.; et al. Multifunctional RNA Nanoparticles. Nano Lett. 2014, 14, 5662–5671. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Ebright, J.N.; Stovall, G.M.; Chen, X.; Nguyen, H.H.; Singh, A.; Syrett, A.; Ellington, A.D. Technical and biological issues relevant to cell typing with aptamers. J. Proteome Res. 2009, 8, 2438–2448. [Google Scholar] [CrossRef] [PubMed]
- Panigaj, M.; Reiser, J. Aptamer guided delivery of nucleic acid-based nanoparticles. DNA RNA Nanotechnol. 2016, 2, 42–52. [Google Scholar] [CrossRef]
- Zhao, N.; Bagaria, H.G.; Wong, M.S.; Zu, Y. A nanocomplex that is both tumor cell-selective and cancer gene-specific for anaplastic large cell lymphoma. J. Nanobiotechnol. 2011, 9, 2. [Google Scholar] [CrossRef]
- Powell, D.; Chandra, S.; Dodson, K.; Shaheen, F.; Wiltz, K.; Ireland, S.; Syed, M.; Dash, S.; Wiese, T.; Mandal, T.; et al. Aptamer-functionalized hybrid nanoparticle for the treatment of breast cancer. Eur. J. Pharm. Biopharm. 2017, 114, 108–118. [Google Scholar] [CrossRef]
- Yan, W.; Tao, M.; Jiang, B.; Yao, M.; Jun, Y.; Dai, W.; Tang, Z.; Gao, Y.; Zhang, L.; Chen, X.; et al. Overcoming Drug Resistance in Colon Cancer by Aptamer-Mediated Targeted Co-Delivery of Drug and siRNA Using Grapefruit-Derived Nanovectors. Cell. Physiol. Biochem. 2018, 50, 79–91. [Google Scholar] [CrossRef]
- Wilner, S.E.; Wengerter, B.; Maier, K.; Borba Magalhães, M.D.L.; Del Amo, D.S.; Pai, S.; Opazo, F.; Rizzoli, S.O.; Yan, A.; Levy, M. An RNA alternative to human transferrin: A new tool for targeting human cells. Mol. Ther. Nucleic Acids 2012, 1, e21. [Google Scholar] [CrossRef]
- Alshaer, W.; Hillaireau, H.; Vergnaud, J.; Mura, S.; Deloménie, C.; Sauvage, F.; Ismail, S.; Fattal, E. Aptamer-guided siRNA-loaded nanomedicines for systemic gene silencing in CD-44 expressing murine triple-negative breast cancer model. J. Control. Release 2018, 271, 98–106. [Google Scholar] [CrossRef]
- Kim, E.; Jung, Y.; Choi, H.; Yang, J.; Suh, J.S.; Huh, Y.M.; Kim, K.; Haam, S. Prostate cancer cell death produced by the co-delivery of Bcl-xL shRNA and doxorubicin using an aptamer-conjugated polyplex. Biomaterials 2010, 31, 4592–4599. [Google Scholar] [CrossRef]
- Wu, X.; Ding, B.; Gao, J.; Wang, H.; Fan, W.; Wang, X.; Zhang, W.; Wang, X.; Ye, L.; Zhang, M.; et al. Second-generation aptamer-conjugated PSMA-targeted delivery system for prostate cancer therapy. Int. J. Nanomed. 2011, 6, 1747–1756. [Google Scholar]
- Xu, X.; Wu, J.; Liu, Y.; Saw, P.E.; Tao, W.; Yu, M.; Zope, H.; Si, M.; Victorious, A.; Rasmussen, J.; et al. Multifunctional Envelope-Type siRNA Delivery Nanoparticle Platform for Prostate Cancer Therapy. ACS Nano 2017, 11, 2618–2627. [Google Scholar] [CrossRef]
- Lv, T.; Li, Z.; Xu, L.; Zhang, Y.; Chen, H.; Gao, Y. Chloroquine in combination with aptamer-modified nanocomplexes for tumor vessel normalization and efficient erlotinib/Survivin shRNA co-delivery to overcome drug resistance in EGFR-mutated non-small cell lung cancer. Acta Biomater. 2018, 76, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Dassie, J.P.; Liu, X.Y.; Thomas, G.S.; Whitaker, R.M.; Thiel, K.W.; Stockdale, K.R.; Meyerholz, D.K.; McCaffrey, A.P.; McNamara, J.O.; Giangrande, P.H. Systemic administration of optimized aptamer-siRNA chimeras promotes regression of PSMA-expressing tumors. Nat. Biotechnol. 2009, 27, 839–846. [Google Scholar] [CrossRef]
- Bagalkot, V.; Gao, X. SiRNA-aptamer chimeras on nanoparticles: Preserving targeting functionality for effective gene silencing. ACS Nano 2011, 5, 8131–8139. [Google Scholar] [CrossRef] [PubMed]
- Jafari, R.; Majidi Zolbanin, N.; Majidi, J.; Atyabi, F.; Yousefi, M.; Jadidi-Niaragh, F.; Aghebati-Maleki, L.; Shanehbandi, D.; Soltani Zangbar, M.-S.; Rafatpanah, H. Anti-Mucin1 Aptamer-Conjugated Chitosan Nanoparticles for Targeted Co-Delivery of Docetaxel and IGF-1R siRNA to SKBR3 Metastatic Breast Cancer Cells. Iran. Biomed. J. 2019, 23, 21–33. [Google Scholar] [CrossRef]
- Dong, J.; Cao, Y.; Shen, H.; Ma, Q.; Mao, S.; Li, S.; Sun, J. EGFR aptamer-conjugated liposome-polycation-DNA complex for targeted delivery of SATB1 small interfering RNA to choriocarcinoma cells. Biomed. Pharmacother. 2018, 107, 849–859. [Google Scholar] [CrossRef]
- Ayatollahi, S.; Salmasi, Z.; Hashemi, M.; Askarian, S.; Oskuee, R.K.; Abnous, K.; Ramezani, M. Aptamer-targeted delivery of Bcl-xL shRNA using alkyl modified PAMAM dendrimers into lung cancer cells. Int. J. Biochem. Cell Biol. 2017, 92, 210–217. [Google Scholar] [CrossRef]
- Taghavi, S.; Nia, A.H.; Abnous, K.; Ramezani, M. Polyethylenimine-functionalized carbon nanotubes tagged with AS1411 aptamer for combination gene and drug delivery into human gastric cancer cells. Int. J. Pharm. 2017, 516, 301–312. [Google Scholar] [CrossRef]
- Xu, Y.; Pang, L.; Wang, H.; Xu, C.; Shah, H.; Guo, P.; Shu, D.; Qian, S.Y. Specific delivery of delta-5-desaturase siRNA via RNA nanoparticles supplemented with dihomo-γ-linolenic acid for colon cancer suppression. Redox Biol. 2019, 21, 101085. [Google Scholar] [CrossRef]
- Li, L.; Hu, X.; Zhang, M.; Ma, S.; Yu, F.; Zhao, S.; Liu, N.; Wang, Z.; Wang, Y.; Guan, H.; et al. Dual Tumor-Targeting Nanocarrier System for siRNA Delivery Based on pRNA and Modified Chitosan. Mol. Ther. Nucleic Acids 2017, 8, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhao, H.; Guo, L.; Wang, Y.; Song, J.; Zhao, X.; Li, C.; Hao, L.; Wang, D.; Tang, J. Ultrasound-mediated nanobubble destruction (UMND) facilitates the delivery of A10-3.2 aptamer targeted and siRNA-loaded cationic nanobubbles for therapy of prostate cancer. Drug Deliv. 2018, 25, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, D.; Xu, Z.; Huang, C.; Zhang, C.; He, B.; Mao, C.; Wang, G.; Qian, H. Targeted Delivery of Rab26 siRNA with Precisely Tailored DNA Prism for Lung Cancer Therapy. ChemBioChem 2019, 20, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Jeelani, S.; Jagat Reddy, R.C.; Maheswaran, T.; Asokan, G.S.; Dany, A.; Anand, B. Theranostics: A treasured tailor for tomorrow. J. Pharm. Bioallied Sci. 2014, 6 (Suppl. 1), S6. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.W.; Jeong, H.Y.; Kang, S.J.; Jeong, I.H.; Choi, M.J.; You, Y.M.; Im, C.S.; Song, I.H.; Lee, T.S.; Lee, J.S.; et al. Anti-EGF receptor aptamer-guided co-delivery of anti-cancer siRNAs and quantum dots for theranostics of triple-negative breast cancer. Theranostics 2019, 9, 837. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, P.; Kim, S.; Kang, H.C.; Shim, M.S. Targeted siRNA delivery using aptamer-siRNA chimeras and aptamer-conjugated nanoparticles. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1543. [Google Scholar] [CrossRef]
- Behlke, M.A. Chemical modification of siRNAs for in vivo use. Oligonucleotides 2008, 18, 305–320. [Google Scholar] [CrossRef]
- Keefe, A.D.; Cload, S.T. SELEX with modified nucleotides. Curr. Opin. Chem. Biol. 2008, 12, 448–456. [Google Scholar] [CrossRef]
- Mohamed, M.; Abu Lila, A.S.; Shimizu, T.; Alaaeldin, E.; Hussein, A.; Sarhan, H.A.; Szebeni, J.; Ishida, T. PEGylated liposomes: Immunological responses. Sci. Technol. Adv. Mater. 2019, 20, 710–724. [Google Scholar] [CrossRef]
- Lächelt, U.; Wagner, E. Nucleic Acid Therapeutics Using Polyplexes: A Journey of 50 Years (and Beyond). Chem. Rev. 2015, 115, 11043–11078. [Google Scholar] [CrossRef]
- Diao, Y.; Liu, J.; Ma, Y.; Su, M.; Zhang, H.; Hao, X. A specific aptamer-cell penetrating peptides complex delivered siRNA efficiently and suppressed prostate tumor growth in vivo. Cancer Biol. Ther. 2016, 17, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, N.; Kanwar, J.R.; Athalya, P.K.; Janakiraman, N.; Khetan, V.; Kanwar, R.K.; Eluchuri, S.; Krishnakumar, S. EpCAM aptamer mediated cancer cell specific delivery of EpCAM siRNA using polymeric nanocomplex. J. Biomed. Sci. 2015, 22, 4. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.C.; Mozumdar, S.; Huang, L. Lipid-based systemic delivery of siRNA. Adv. Drug Deliv. Rev. 2009, 61, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Hu-Lieskovan, S.; Wargo, J.A.; Ribas, A. Primary, Adaptive, and Acquired Resistance to Cancer Immunotherapy. Cell 2017, 168, 707–723. [Google Scholar] [CrossRef]
- Pastor, F.; Berraondo, P.; Etxeberria, I.; Frederick, J.; Sahin, U.; Gilboa, E.; Melero, I. An rna toolbox for cancer immunotherapy. Nat. Rev. Drug Discov. 2018, 17, 751–767. [Google Scholar] [CrossRef]
- Pratico, E.D.; Sullenger, B.A.; Nair, S.K. Identification and characterization of an agonistic aptamer against the T cell costimulatory receptor, OX40. Nucleic Acid Ther. 2013, 23, 35–43. [Google Scholar] [CrossRef]
- Rajagopalan, A.; Berezhnoy, A.; Schrand, B.; Puplampu-Dove, Y.; Gilboa, E. Aptamer-Targeted Attenuation of IL-2 Signaling in CD8+ T Cells Enhances Antitumor Immunity. Mol. Ther. 2017, 25, 54–61. [Google Scholar] [CrossRef]
- Puplampu-Dove, Y.; Gefen, T.; Rajagopalan, A.; Muheramagic, D.; Schrand, B.; Gilboa, E. Potentiating tumor immunity using aptamer-targeted RNAi to render CD8+ T cells resistant to TGFβ inhibition. Oncoimmunology 2018, 7, e1349588. [Google Scholar] [CrossRef]
- Song, P.; Chou, Y.K.; Zhang, X.; Meza-Romero, R.; Yomogida, K.; Benedek, G.; Chu, C.Q. CD4 aptamer-RORγt shRNA chimera inhibits IL-17 synthesis by human CD4 + T cells. Biochem. Biophys. Res. Commun. 2014, 452, 1040–1045. [Google Scholar] [CrossRef]
- Soldevilla, M.M.; Villanueva, H.; Bendandi, M.; Inoges, S.; López-Díaz de Cerio, A.; Pastor, F. 2-fluoro-RNA oligonucleotide CD40 targeted aptamers for the control of B lymphoma and bone-marrow aplasia. Biomaterials 2015, 67, 274–285. [Google Scholar] [CrossRef]
- Nozari, A.; Berezovski, M.V. Aptamers for CD Antigens: From Cell Profiling to Activity Modulation. Mol. Ther. Nucleic Acids 2017, 6, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Kruspe, S.; Giangrande, P.H. Aptamer-siRNA chimeras: Discovery, progress, and future prospects. Biomedicines 2017, 5, 45. [Google Scholar] [CrossRef] [PubMed]
- Strebhardt, K.; Ullrich, A. Paul Ehrlich’s magic bullet concept: 100 Years of progress. Nat. Rev. Cancer 2008, 8, 473–480. [Google Scholar] [CrossRef] [PubMed]
- McNamara, J.O.; Andrechek, E.R.; Wang, Y.; Viles, K.D.; Rempel, R.E.; Gilboa, E.; Sullenger, B.A.; Giangrande, P.H. Cell type-specific delivery of siRNAs with aptamer-siRNA chimeras. Nat. Biotechnol. 2006, 24, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Esposito, C.L.; Nuzzo, S.; Catuogno, S.; Romano, S.; de Nigris, F.; de Franciscis, V. STAT3 Gene Silencing by Aptamer-siRNA Chimera as Selective Therapeutic for Glioblastoma. Mol. Ther. Nucleic Acids 2018, 10, 398–411. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Maihle, N.J.; Yu, X.; Tang, S.C.; Liu, H.Y. Synergistic Targeting HER2 and EGFR with Bivalent Aptamer-siRNA Chimera Efficiently Inhibits HER2-Positive Tumor Growth. Mol. Pharm. 2018, 15, 4801–4813. [Google Scholar] [CrossRef]
- Yu, X.; Ghamande, S.; Liu, H.; Xue, L.; Zhao, S.; Tan, W.; Zhao, L.; Tang, S.C.; Wu, D.; Korkaya, H.; et al. Targeting EGFR/HER2/HER3 with a Three-in-One Aptamer-siRNA Chimera Confers Superior Activity against HER2+ Breast Cancer. Mol. Ther. Nucleic Acids 2018, 10, 317–330. [Google Scholar] [CrossRef]
- Jeong, H.; Lee, S.H.; Hwang, Y.; Yoo, H.; Jung, H.; Kim, S.H.; Mok, H. Multivalent Aptamer–RNA Conjugates for Simple and Efficient Delivery of Doxorubicin/siRNA into Multidrug-Resistant Cells. Macromol. Biosci. 2017, 17, 1600343. [Google Scholar] [CrossRef]
- Xiang, D.; Shigdar, S.; Bean, A.G.; Bruce, M.; Yang, W.; Mathesh, M.; Wang, T.; Yin, W.; Tran, P.H.L.; Shamaileh, H.A.; et al. Transforming doxorubicin into a cancer stem cell killer via EpCAM aptamer-mediated delivery. Theranostics 2017, 7, 4071. [Google Scholar] [CrossRef]
- Alshamaileh, H.; Wang, T.; Xiang, D.; Yin, W.; Tran, P.H.L.; Barrero, R.A.; Zhang, P.Z.; Li, Y.; Kong, L.; Liu, K.; et al. Aptamer-mediated survivin RNAi enables 5-fluorouracil to eliminate colorectal cancer stem cells. Sci. Rep. 2017, 7, 5898. [Google Scholar] [CrossRef]
- Dou, X.Q.; Wang, H.; Zhang, J.; Wang, F.; Xu, G.L.; Xu, C.C.; Xu, H.H.; Xiang, S.S.; Fu, J.; Song, H.F. Aptamer–drug conjugate: Targeted delivery of doxorubicin in a HER3 aptamer-functionalized liposomal delivery system reduces cardiotoxicity. Int. J. Nanomed. 2018, 13, 763–776. [Google Scholar] [CrossRef]
- Prusty, D.K.; Adam, V.; Zadegan, R.M.; Irsen, S.; Famulok, M. Supramolecular aptamer nano-constructs for receptor-mediated targeting and light-triggered release of chemotherapeutics into cancer cells. Nat. Commun. 2018, 9, 535. [Google Scholar] [CrossRef] [PubMed]
- Bayraç, A.T.; Akça, O.E.; Eyidoğan, F.İ.; Öktem, H.A. Target-specific delivery of doxorubicin to human glioblastoma cell line via ssDNA aptamer. J. Biosci. 2018, 43, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Yan, Z.; Jin, K.; Pang, Q.; Jiang, T.; Lu, H.; Liu, X.; Pang, Z.; Yu, L.; Jiang, X. Precise glioblastoma targeting by AS1411 aptamer-functionalized poly (l-γ-glutamylglutamine)–paclitaxel nanoconjugates. J. Colloid Interface Sci. 2017, 490, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanyam, J.; Badrinarayanan, L.; Dhaka, B.; Gowda, H.; Pandey, A.; Subramanian, K.; Subadhra, L.B.; Elchuri, S. V EpCAM Aptamer siRNA chimeras: Therapeutic efficacy in epithelial cancer cells. bioRxiv 2019. [Google Scholar] [CrossRef]
- Li, H.; Yang, S.; Yu, G.; Shen, L.; Fan, J.; Xu, L.; Zhang, H.; Zhao, N.; Zeng, Z.; Hu, T.; et al. Aptamer internalization via endocytosis inducing s-phase arrest and priming maver-1 lymphoma cells for cytarabine chemotherapy. Theranostics 2017, 7, 1204–1213. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kratschmer, C.; Levy, M. Targeted Delivery of Auristatin-Modified Toxins to Pancreatic Cancer Using Aptamers. Mol. Ther. Nucleic Acids 2018, 10, 227–236. [Google Scholar] [CrossRef]
- Zolbanin, N.M.; Jafari, R.; Majidi, J.; Atyabi, F.; Yousefi, M.; Jadidi-Niaragh, F.; Aghebati-Maleki, L.; Shanehbandi, D.; Zangbar, M.S.S.; Nayebi, A.M. Targeted co-delivery of docetaxel and cMET siRNA for treatment of mucin1 overexpressing breast cancer cells. Adv. Pharm. Bull. 2018, 8, 383–393. [Google Scholar] [CrossRef]
- de Almeida, C.E.B.; Alves, L.N.; Rocha, H.F.; Cabral-Neto, J.B.; Missailidis, S. Aptamer delivery of siRNA, radiopharmaceutics and chemotherapy agents in cancer. Int. J. Pharm. 2017, 525, 334–342. [Google Scholar] [CrossRef]


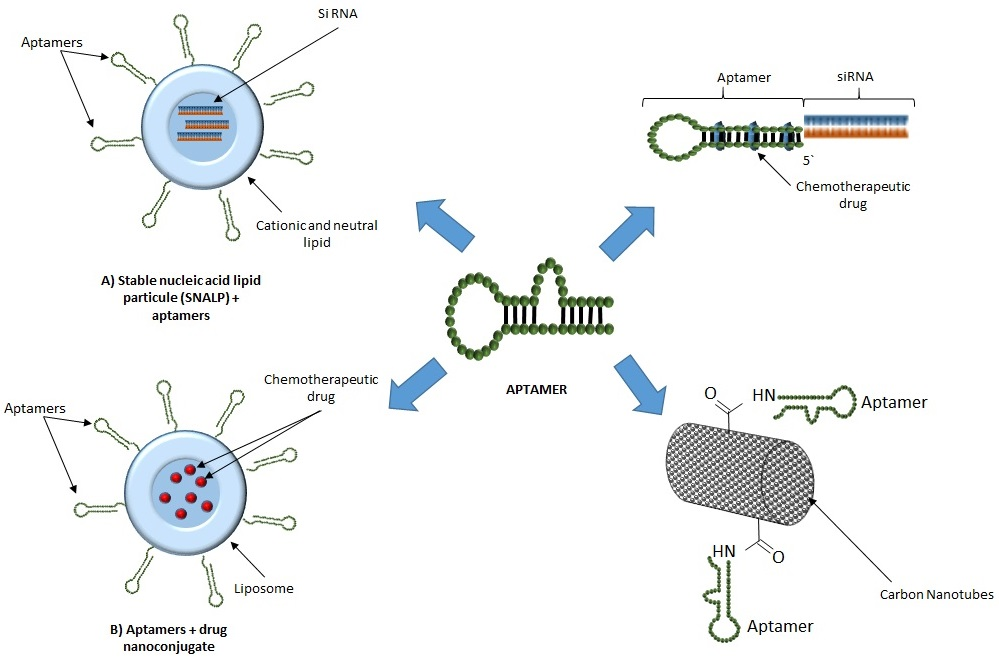{kind=link}
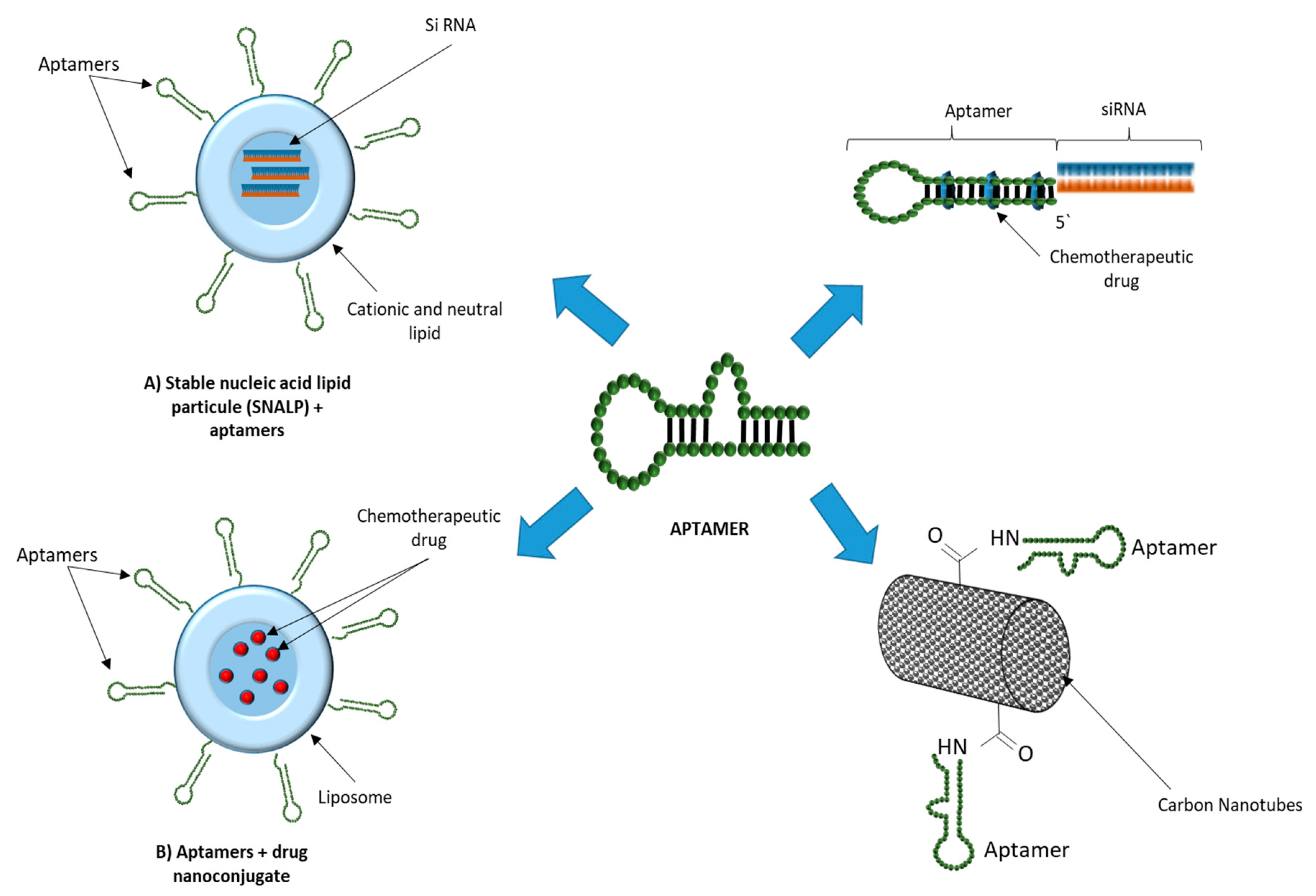{kind=link}
| siRNA/miRNA/shRNA Target Gene | Carrier Aptamer | Efficacy Tests | Ref. |
|---|---|---|---|
| Survivin | CD4 | In vitro knock down antiapoptosis factor survivin | [12] |
| ICAM 1 | FB4 (bind to transferrin receptors) | In vitro results showed a decreased of ICAM-1 expression and blocked the adhesion of monocytes | [13] |
| Human immunodeficiency virus (HIV)-1 | epidermal growth factor receptor (EGFR) | In vitro and in vivo gene silencing | [14] |
| Anaplastic lymphoma kinase (ALK) | CD30 | In vitro results of growth arrest and apoptosis | [17] |
| P-gp | A6 (bind to human epidermal growth factor receptor 2 (HER-2) receptors) | In vitro results about decreasing of resistance to chemotherapeutics | [18] |
| P-gp | LA1 (specific to multi-drug resistance (MDR)) | In vitro and in vivo anti-tumor activity | [19] |
| GFP | Transferrin | In vitro enhanced siRNA uptake and target gene knockdown | [20] |
| Luciferase | CD44 | In vitro and in vivo results of targeted gene silencing in CD44-positive breast cancer | [21] |
| P-gp | HER2 | In vitro confirmation of silencing disease-related genes in tumors | [18] |
| BCL-xL | Prostate-specific membrane antigen (PSMA) | In vitro anti-tumor activity | [22] |
| Genes in prostate cancer | PSMA | In vitro anticancer effect | [23] |
| Prohibitin 1 | PSMA | In vivo gene silencing in prostate cancer tumor cells | [24] |
| Survivin | EGFR | In vitro and in vivo antitumor effects | [25] |
| GFP | PSMA | In vitro GFP silencing/Fluorescence imaging (quantum dots) | [27] |
| Insulin-like growth factor receptor 1 (IGF-1R) | MUC-1 | In vitro augment the targeting of pathways involved in tumorigenesis and metastasis | [28] |
| Special AT-rich sequence binding protein 1 (SATB1) | EGFR | In vitro and in vivo studies of gene expression decreased | [29] |
| BCL-xL | Nucleolin | in vitro gene silencing and apoptosis | [30] |
| BCL-xL | Nucelolin | In vitro gene silencing and tumoricidal efficacy | [31] |
| D5D | EpCAM | In vitro and in vivo inhibition of D5D expression | [32] |
| c-myc | Transferrin | In vitro and in vivo anti-tumor activity | [33] |
| FoxM1 | PSMA | In vitro and in vivo anti-tumor activity in prostate cancer cells and xenografts in mice | [34] |
| Rab26 | MUC-1 | In vitro anti-tumor activity | [35] |
| bcl-2 | EGFR | In vitro interference of tumor cell proliferation | [11] |
| bcl-2 and PKC-ι | EGFR | In vivo delivery and therapeutic efficacy | [37] |
| siRNA Target Gene | Carrier Aptamer | Efficacy Tests | Ref. |
|---|---|---|---|
| IL-2 Rα (CD25) | 41-BB | In vitro using CHO cells and murine CD8+ cells and in vivo using C57BL/6 mice adoptively transferred with OT-I cells and breast tumor 4T1 model | [49] |
| Smad-4 | 41-BB | In vitro using murine CD8+ cells and in vivo using breast tumor 4T1 model | [50] |
| RORγT | CD4 | In vitro using CD4+ Karpas 299 cells and primary human CD4+ T cells | [51] |
| SMG-1 | CD40 agonist | In vitro using human B- lymphocytes and in vivo using B-cell lymphoma-bearing mice | [52] |
| Carrier Aptamers | siRNA or Drug Delivered | Target Cell Lines | Main Effects | Ref. |
|---|---|---|---|---|
| PDGFRβ (Gint4.T) | STAT-3 siRNA | U87MG and T98G glioblastoma cells | Reduction of anti-apoptotic factors PARP and BCL-xL, inducing cell death in two glioblastoma lineages in vitro. Reduction of tumor growth rate and pro-tumoral factors in a xenograft mouse model of glioblastoma | [57] |
| HER-2 sense& antisense bivalent | EGFR siRNA | BT474 and SKBR3 breast cancer cells | Downregulation of EGFR in vitro and suppression of xenografted tumor growth in vivo | [58] |
| bivalent HER-2-HER-3 | EGFR siRNA | BT474 and SKBR3 breast cancer cells | Reduction of HER receptors, induction of cell cycle arrest and apoptosis in vitro and inhibition of tumor growth in vivo | [59] |
| MUC1 multiaptamer | BCL-2 siRNA and Doxorubicin (DOX) | MCF-7 breast cancer cells | More efficiency in delivering doxorubicin. BCL-2 siRNA acted synergistically with DOX increasing the sensitivity of cells for apoptosis | [60] |
| epithelial cell adhesion molecule (EpCAM) | Doxorubicin (DOX) | EpCAM positive HT29 colorectal cancer cells; SCOV-3 ovarian cancer cell line and T47D breast cancer cell line | Tumor growth control overcoming chemoresistance in both tumorsphere formation assay and in xenotransplanted mice | [61] |
| EpCAM | Survivin RNAi | EpCAM positive HT29 colorectal cancer cells | The combination of the surivin downregulation with the action of 5-fluorouacil cytotoxic chemotherapeutic could increase the lethality of colorectal cancer stem cells, as well as tumor control, in colorectal cancer xenograph model | [62] |
| HER-3ECD | Doxorubicin (DOX) | MCF-7 HER3+ and BT 474HER3+ breast cancer cells | Increase in the survival rate in a xenograft mouse model. Reduction of liver and cardio toxicity after chemotherapy when compared to the formulation without aptamer | [63] |
| cMET | Doxorubicin (DOX) | NCI-H1838 lung cancer cells | Better efficiency in the uptake by the target cell and a precise subcellular distribution of DOX to the cell nucleus, after endocytosis | [64] |
| Anti A-172 cells aptamer | Doxorubicin (DOX) | Glioblastoma multiforme tumor cells A-172 | Selective binding and inactivation of A-172 glioblastoma cells | [65] |
| Nucleolin (AS1411) | Paclitaxel (PCX) | Glioblastoma U87 MG cells and neo-vascular endothelial cells | Improvement of cell uptake and internalization, spheroid tumor penetration and cell growth inhibition. Enhanced accumulation of AS1411-PGG-PTX nanoconjugates in glioblastoma cells, retention time in circulation penetration in glioblastoma tumors. Increase survival of the tumor bearing mice and an efficient PCX delivery to glioblastoma tissue with consequently strong cytotoxic effect | [66] |
| EpCAM | PLK1, BCL2, and STAT3 siRNA | NCC RbC 51 retinoblastoma cells; MCF-7 breast cancer cells; Müller Glial Mio M1 cells; PMI 2650 Head and Neck cancer cells and HepG2 Hepatocellular cells | Cell death induction and tumor reduction on RB xenografts tumor model | [67] |
| HL-1 (anti Maver-I cells) | Cytarabine | Maver-I lymphoma cells | Aptamer treatment was able to sensitize synergistically lymphoma cells in S-phase to lethal effect of cytarabine | [68] |
| Waz (anti-transferrin receptor—TfR) | Auristatin modified toxins | Panc-1, MIA PaCa-2, and BxPC3 Pancreas Ductal Adenocarcinoma cells | Aptamer conjugates demonstrated to be toxic to cell lines in different extends | [69] |
| EGFR | Auristatin modified toxins | Panc-1, MIA PaCa-2 and BxPC3 Pancreas Ductal Adenocarcinoma cells | Aptamer-toxin conjugates demonstrated to be toxic to cell lines in different extends | [69] |
| MUC1 | cMET siRNA and Docetaxel | SKBR3 breast cancer cells | Nanoparticle chitosan increased specificity and cellular uptake of the nanoconjugate and successful silencing was confirmed | [70] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dinis Ano Bom, A.P.; da Costa Neves, P.C.; Bonacossa de Almeida, C.E.; Silva, D.; Missailidis, S. Aptamers as Delivery Agents of siRNA and Chimeric Formulations for the Treatment of Cancer. Pharmaceutics 2019, 11, 684. https://doi.org/10.3390/pharmaceutics11120684
Dinis Ano Bom AP, da Costa Neves PC, Bonacossa de Almeida CE, Silva D, Missailidis S. Aptamers as Delivery Agents of siRNA and Chimeric Formulations for the Treatment of Cancer. Pharmaceutics. 2019; 11(12):684. https://doi.org/10.3390/pharmaceutics11120684
Chicago/Turabian StyleDinis Ano Bom, Ana Paula, Patrícia Cristina da Costa Neves, Carlos Eduardo Bonacossa de Almeida, Dilson Silva, and Sotiris Missailidis. 2019. "Aptamers as Delivery Agents of siRNA and Chimeric Formulations for the Treatment of Cancer" Pharmaceutics 11, no. 12: 684. https://doi.org/10.3390/pharmaceutics11120684
APA StyleDinis Ano Bom, A. P., da Costa Neves, P. C., Bonacossa de Almeida, C. E., Silva, D., & Missailidis, S. (2019). Aptamers as Delivery Agents of siRNA and Chimeric Formulations for the Treatment of Cancer. Pharmaceutics, 11(12), 684. https://doi.org/10.3390/pharmaceutics11120684