Impact of Viral Inflammation on the Expression of Renal Drug Transporters in Pregnant Rats

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. RNA Extraction and Quantitative Real Time Polymerase Chain Reaction
2.3. Membrane Protein Extraction and Western Blot Analysis
2.4. Data and Statistical Analysis
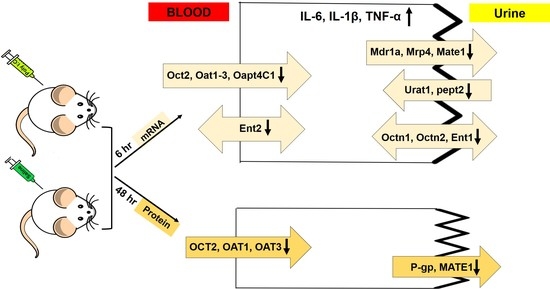
3. Results
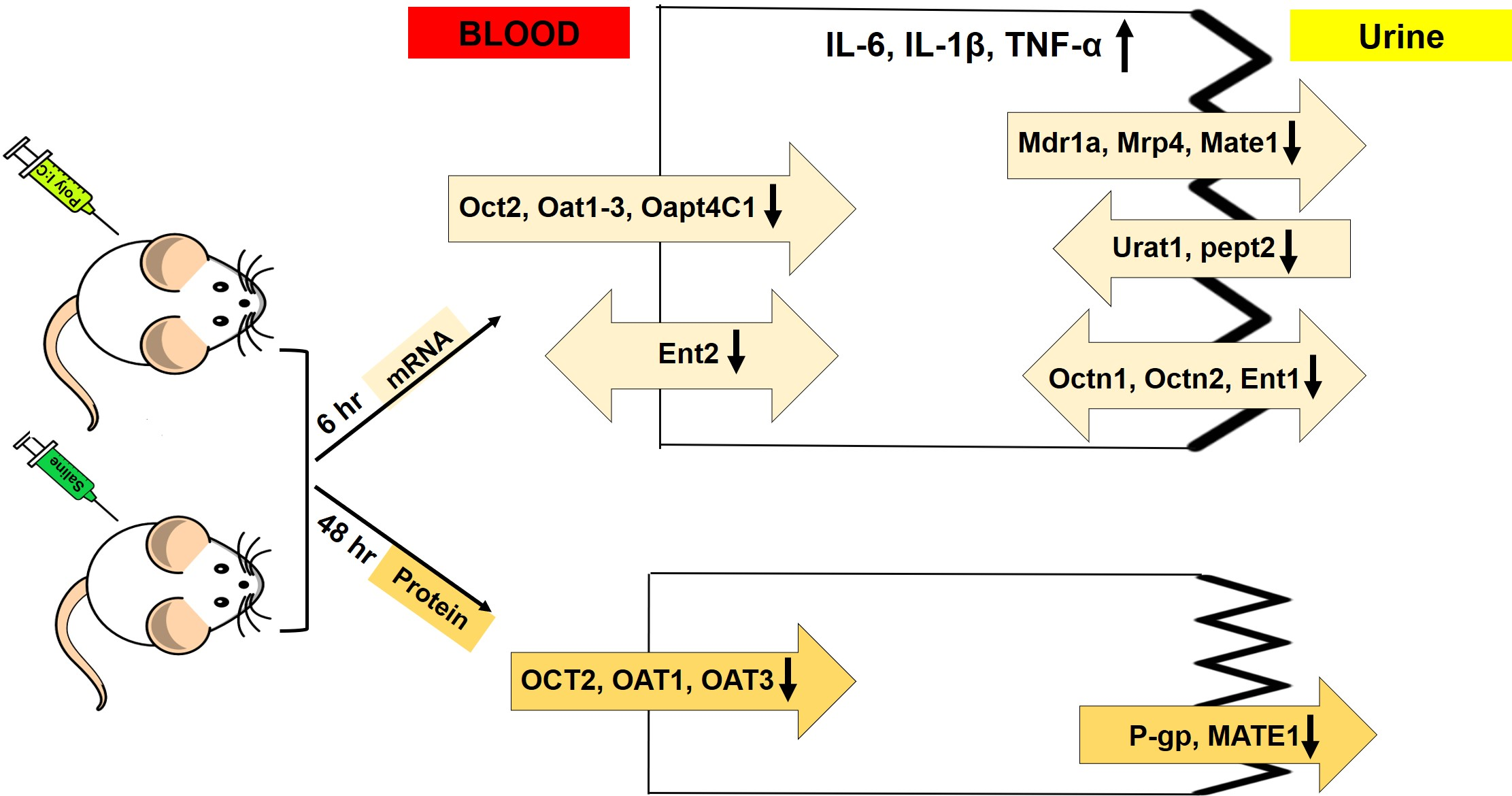
3.1. Effect of Poly I:C on the Inflammatory Response
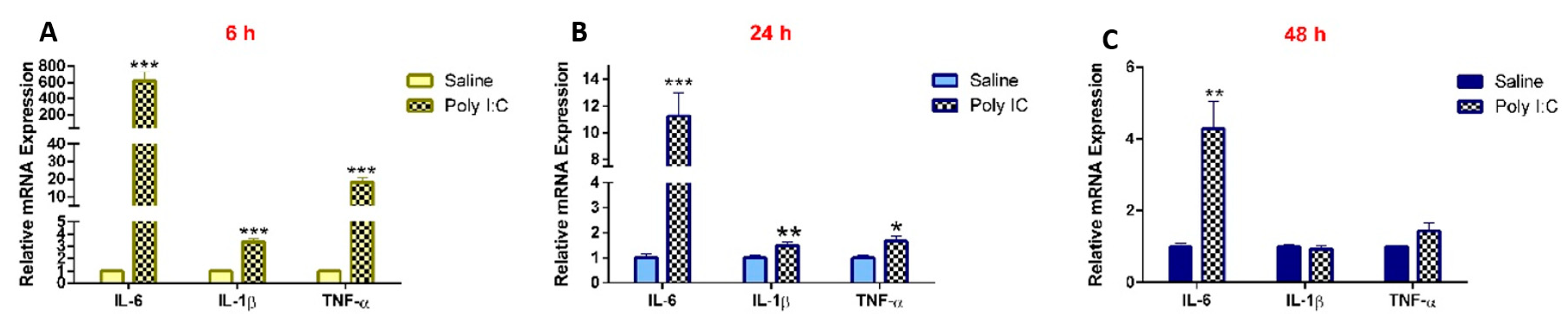
3.2. Effect of Poly I:C on the mRNA Expression of Renal Drug Transporters
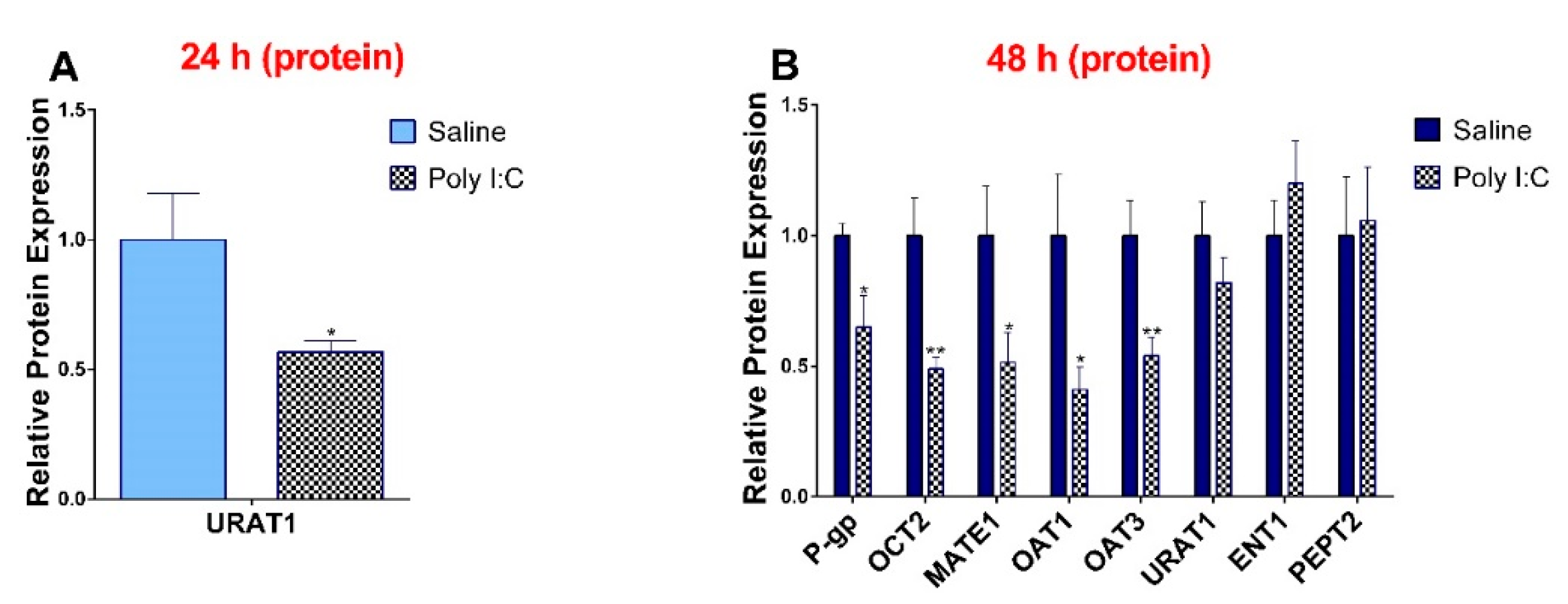
3.3. Effect of Poly I:C on the Protein Expression of Renal Drug Transporters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Racicot, K.; Mor, G. Risks associated with viral infections during pregnancy. J. Clin. Investig. 2017, 127, 1591–1599. [Google Scholar] [CrossRef]
- Abualsunun, W.A.; Piquette-Miller, M. Involvement of Nuclear Factor kappaB, not Pregnane X Receptor, in Inflammation-Mediated Regulation of Hepatic Transporters. Drug Metab. Dispos. 2017, 45, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Sukhai, M.; Yong, A.; Kalitsky, J.; Piquette-Miller, M. Inflammation and interleukin-6 mediate reductions in the hepatic expression and transcription of the mdr1a and mdr1b Genes. Mol. Cell Biol. Res. Commun. 2000, 4, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Sukhai, M.; Yong, A.; Pak, A.; Piquette-Miller, M. Decreased expression of P-glycoprotein in interleukin-1beta and interleukin-6 treated rat hepatocytes. Inflamm. Res. 2001, 50, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, G.; Kim, H.; Piquette-Miller, M. Regulation of the hepatic multidrug resistance gene expression by endotoxin and inflammatory cytokines in mice. Int. Immunopharmacol. 2001, 1, 189–199. [Google Scholar] [CrossRef]
- Lee, G.; Piquette-Miller, M. Influence of IL-6 on MDR and MRP-mediated multidrug resistance in human hepatoma cells. Can. J. Physiol. Pharmacol. 2001, 79, 876–884. [Google Scholar] [CrossRef]
- Christensen, H.; Hermann, M. Immunological response as a source to variability in drug metabolism and transport. Front. Pharmacol. 2012, 3, 8. [Google Scholar] [CrossRef]
- Fardel, O.; Le Vee, M. Regulation of human hepatic drug transporter expression by pro-inflammatory cytokines. Expert Opin. Drug Metab. Toxicol. 2009, 5, 1469–1481. [Google Scholar] [CrossRef]
- Petrovic, V.; Teng, S.; Piquette-Miller, M. Regulation of drug transporters during infection and inflammation. Mol. Interv. 2007, 7, 99–111. [Google Scholar] [CrossRef]
- Morgan, E.T.; Goralski, K.B.; Piquette-Miller, M.; Renton, K.W.; Robertson, G.R.; Chaluvadi, M.R.; Charles, K.A.; Clarke, S.J.; Kacevska, M.; Liddle, C.; et al. Regulation of drug-metabolizing enzymes and transporters in infection, inflammation, and cancer. Drug Metab. Dispos. 2008, 36, 205–216. [Google Scholar] [CrossRef]
- Bianchi, F.; Pretto, S.; Tagliabue, E.; Balsari, A.; Sfondrini, L. Exploiting poly (I:C) to induce cancer cell apoptosis. Cancer Biol. Ther. 2017, 18, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Gabay, C.; Kushner, I. Acute-phase proteins and other systemic responses to inflammation. N. Engl. J. Med. 1999, 340, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Haslett, C. Resolution of acute inflammation and the role of apoptosis in the tissue fate of granulocytes. Clin. Sci. 1992, 83, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, V.; Piquette-Miller, M. Impact of polyinosinic/polycytidylic acid on placental and hepatobiliary drug transporters in pregnant rats. Drug Metab. Dispos. 2010, 38, 1760–1766. [Google Scholar] [CrossRef] [PubMed]
- Pour, N.K.; Piquette-Miller, M. Dysregulation of Renal Transporters in a Rodent Model of Viral Infection. Clin. Pharmacol. Ther. 2018, 103, 1096. [Google Scholar]
- McColl, E.R.; Piquette-Miller, M. Poly(I:C) alters placental and fetal brain amino acid transport in a rat model of maternal immune activation. Am. J. Reprod. Immunol. 2019, 81, e13115. [Google Scholar] [CrossRef]
- Shuster, D.L.; Bammler, T.K.; Beyer, R.P.; Macdonald, J.W.; Tsai, J.M.; Farin, F.M.; Hebert, M.F.; Thummel, K.E.; Mao, Q. Gestational age-dependent changes in gene expression of metabolic enzymes and transporters in pregnant mice. Drug Metab. Dispos. 2013, 41, 332–342. [Google Scholar] [CrossRef]
- Aghaeepour, N.; Ganio, E.A.; McIlwain, D.; Tsai, A.S.; Tingle, M.; Van Gassen, S.; Gaudilliere, D.K.; Baca, Q.; McNeil, L.; Okada, R.; et al. An immune clock of human pregnancy. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef]
- Schmidt, C.; Hocherl, K.; Bucher, M. Regulation of renal glucose transporters during severe inflammation. Am. J. Physiol. Ren. Physiol. 2007, 292, F804–F811. [Google Scholar] [CrossRef]
- Schmidt, C.; Hocherl, K.; Schweda, F.; Kurtz, A.; Bucher, M. Regulation of renal sodium transporters during severe inflammation. J. Am. Soc. Nephrol. 2007, 18, 1072–1083. [Google Scholar] [CrossRef] [PubMed]
- Zamek-Gliszczynski, M.J.; Taub, M.E.; Chothe, P.P.; Chu, X.; Giacomini, K.M.; Kim, R.B.; Ray, A.S.; Stocker, S.L.; Unadkat, J.D.; Wittwer, M.B.; et al. Transporters in Drug Development: 2018 ITC Recommendations for Transporters of Emerging Clinical Importance. Clin. Pharmacol. Ther. 2018, 104, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Ivanyuk, A.; Livio, F.; Biollaz, J.; Buclin, T. Renal Drug Transporters and Drug Interactions. Clin. Pharmacokinet. 2017, 56, 825–892. [Google Scholar] [CrossRef]
- Morrissey, K.M.; Stocker, S.L.; Wittwer, M.B.; Xu, L.; Giacomini, K.M. Renal transporters in drug development. Annu. Rev. Pharmacol. Toxicol. 2016, 53, 503–529. [Google Scholar] [CrossRef]
- Yin, J.; Wang, J. Renal drug transporters and their significance in drug-drug interactions. Acta Pharm. Sin. B 2016, 6, 363–373. [Google Scholar] [CrossRef]
- Mihai, S.; Codrici, E.; Popescu, I.D.; Enciu, A.M.; Albulescu, L.; Necula, L.G.; Mambet, C.; Anton, G.; Tanase, C. Inflammation-Related Mechanisms in Chronic Kidney Disease Prediction, Progression, and Outcome. J. Immunol. Res. 2018, 2018, 2180373. [Google Scholar] [CrossRef]
- Naud, J.; Dumayne, C.; Nolin, T.D.; Leblond, F.A.; Pichette, V. Drug pharmacokinetics in renal failure: What’s new? Nephrol. Ther. 2015, 11, 144–151. [Google Scholar] [CrossRef]
- Ando, H.; Nishio, Y.; Ito, K.; Nakao, A.; Wang, L.; Zhao, Y.L.; Kitaichi, K.; Takagi, K.; Hasegawa, T. Effect of endotoxin on P-glycoprotein-mediated biliary and renal excretion of rhodamine-123 in rats. Antimicrob. Agents Chemother. 2001, 45, 3462–3467. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Morisaki, T.; Sugimoto, W.; Yokoo, K.; Sato, D.; Nonoguchi, H.; Tomita, K.; Terada, T.; Inui, K.; Hamada, A.; et al. Altered pharmacokinetics of cationic drugs caused by down-regulation of renal rat organic cation transporter 2 (Slc22a2) and rat multidrug and toxin extrusion 1 (Slc47a1) in ischemia/reperfusion-induced acute kidney injury. Drug Metab. Dispos. 2008, 36, 649–654. [Google Scholar] [CrossRef]
- Schneider, R.; Sauvant, C.; Betz, B.; Otremba, M.; Fischer, D.; Holzinger, H.; Wanner, C.; Galle, J.; Gekle, M. Downregulation of organic anion transporters OAT1 and OAT3 correlates with impaired secretion of para-aminohippurate after ischemic acute renal failure in rats. Am. J. Physiol. Renal Physiol. 2007, 292, F1599–F1605. [Google Scholar] [CrossRef]
- Lindsay, R.S.; Loeken, M.R. Metformin use in pregnancy: Promises and uncertainties. Diabetologia 2017, 60, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Goswami, S.; Giacomini, K.M.; Altman, R.B.; Klein, T.E. Metformin pathways: Pharmacokinetics and pharmacodynamics. Pharmacogenet. Genomics 2012, 22, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Berns, J.S.; Kasbekar, N. Highly active antiretroviral therapy and the kidney: An update on antiretroviral medications for nephrologists. Clin. J. Am. Soc. Nephrol. 2006, 1, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Alam, C.; Whyte-Allman, S.K.; Omeragic, A.; Bendayan, R. Role and modulation of drug transporters in HIV-1 therapy. Adv. Drug Deliv. Rev. 2016, 103, 121–143. [Google Scholar] [CrossRef] [PubMed]
- Anger, G.J.; Piquette-Miller, M. Pharmacokinetic studies in pregnant women. Clin. Pharmacol. Ther. 2008, 83, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Field, R.; Campion, S.; Warren, C.; Murray, C.; Cunningham, C. Systemic challenge with the TLR3 agonist poly I:C induces amplified IFNalpha/beta and IL-1beta responses in the diseased brain and exacerbates chronic neurodegeneration. Brain Behav. Immun. 2010, 24, 996–1007. [Google Scholar] [CrossRef]
- Murray, C.; Griffin, É.W.; O’Loughlin, E.; Lyons, A.; Sherwin, E.; Ahmed, S.; Stevenson, N.J.; Harkin, A.; Cunningham, C. Interdependent and independent roles of type I interferons and IL-6 in innate immune, neuroinflammatory and sickness behaviour responses to systemic poly I:C. Brain Behav. Immun. 2015, 48, 274–286. [Google Scholar] [CrossRef]
- Lever, A.R.; Park, H.; Mulhern, T.J.; Jackson, G.R.; Comolli, J.C.; Borenstein, J.T.; Hayden, P.J.; Prantil-Baun, R. Comprehensive evaluation of poly(I:C) induced inflammatory response in an airway epithelial model. Physiol. Rep. 2015, 3, e12334. [Google Scholar] [CrossRef]
- Kumar, A.; Zhang, J.; Yu, F.S. Toll-like receptor 3 agonist poly(I:C)-induced antiviral response in human corneal epithelial cells. Immunology 2006, 117, 11–21. [Google Scholar] [CrossRef]
- Hasan, M.; Ruksznis, C.; Wang, Y.; Leifer, C.A. Antimicrobial peptides inhibit polyinosinic-polycytidylic acid-induced immune responses. J. Immunol. 2011, 187, 5653–5659. [Google Scholar] [CrossRef]
- Merrell, M.D.; Nyagode, B.A.; Clarke, J.D.; Cherrington, N.J.; Morgan, E.T. Selective and cytokine-dependent regulation of hepatic transporters and bile acid homeostasis during infectious colitis in mice. Drug Metab. Dispos. 2014, 42, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Geier, A.; Dietrich, C.G.; Voigt, S.; Kim, S.K.; Gerloff, T.; Kullak-Ublick, G.A.; Lorenzen, J.; Matern, S.; Gartung, C. Effects of proinflammatory cytokines on rat organic anion transporters during toxic liver injury and cholestasis. Hepatology 2003, 38, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Le Vee, M.; Lecureur, V.; Stieger, B.; Fardel, O. Regulation of drug transporter expression in human hepatocytes exposed to the proinflammatory cytokines tumor necrosis factor-alpha or interleukin-6. Drug Metab. Dispos. 2009, 37, 685–693. [Google Scholar] [CrossRef]
- Abualsunun, W.A.; Piquette-Miller, M. STAT3 is involved in IL-6-Mediated Downregulation of Hepatic Transporters in Mice. J. Pharm. Pharm. Sci. 2018, 21, 325s–334s. [Google Scholar] [CrossRef]
- Copeland, S.; Warren, H.S.; Lowry, S.F.; Calvano, S.E.; Remick, D. Inflammation and the Host Response to Injury Investigators. Acute inflammatory response to endotoxin in mice and humans. Clin. Diagn. Lab. Immunol. 2005, 12, 60–67. [Google Scholar]
- Roe, A.L.; Warren, G.; Hou, G.; Howard, G.; Shedlofsky, S.I.; Blouin, R.A. The effect of high dose endotoxin on CYP3A2 expression in the rat. Pharm. Res. 1998, 15, 1603–1608. [Google Scholar] [CrossRef]
- Poloyac, S.M.; Tosheva, R.T.; Gardner, B.M.; Shedlofsky, S.I.; Blouin, R.A. The effect of endotoxin administration on the pharmacokinetics of chlorzoxazone in humans. Clin. Pharmacol. Ther. 1999, 66, 554–562. [Google Scholar] [CrossRef]
- Devault, A.; Gros, P. Two members of the mouse mdr gene family confer multidrug resistance with overlapping but distinct drug specificities. Mol. Cell. Biol. 1990, 10, 1652–1663. [Google Scholar] [CrossRef]
- Barrand, M.A.; Twentyman, P.R. Differential recognition of mdr1a and mdr1b gene products in multidrug resistant mouse tumour cell lines by different monoclonal antibodies. Br. J. Cancer 1992, 65, 239–245. [Google Scholar] [CrossRef][Green Version]
- Wen, J.; Luo, J.; Huang, W.; Tang, J.; Zhou, H.; Zhang, W. The Pharmacological and Physiological Role of Multidrug-Resistant Protein 4. J. Pharmacol. Exp. Ther. 2015, 354, 358–375. [Google Scholar] [CrossRef]
- Brcakova, E.; Fuksa, L.; Cermanova, J.; Kolouchova, G.; Hroch, M.; Hirsova, P.; Martinkova, J.; Staud, F.; Micuda, S. Alteration of methotrexate biliary and renal elimination during extrahepatic and intrahepatic cholestasis in rats. Biol. Pharm. Bull. 2009, 32, 1978–1985. [Google Scholar] [CrossRef][Green Version]
- Gibson, C.J.; Hossain, M.M.; Richardson, J.R.; Aleksunes, L.M. Inflammatory regulation of ATP binding cassette efflux transporter expression and function in microglia. J. Pharmacol. Exp. Ther. 2012, 343, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Khamdang, S.; Takeda, M.; Shimoda, M.; Noshiro, R.; Narikawa, S.; Huang, X.L.; Enomoto, A.; Piyachaturawat, P.; Endou, H. Interactions of human- and rat-organic anion transporters with pravastatin and cimetidine. J. Pharmacol. Sci. 2004, 94, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H.; Endou, H. The SLC22 drug transporter family. Pflugers Arch. 2004, 447, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Maeda, A.; Tsuruoka, S.; Kanai, Y.; Endou, H.; Saito, K.; Miyamoto, E.; Fujimura, A. Evaluation of the interaction between nonsteroidal anti-inflammatory drugs and methotrexate using human organic anion transporter 3-transfected cells. Eur. J. Pharmacol. 2008, 596, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Hocherl, K.; Schmidt, C.; Bucher, M. COX-2 inhibition attenuates endotoxin-induced downregulation of organic anion transporters in the rat renal cortex. Kidney Int. 2009, 75, 373–380. [Google Scholar] [CrossRef]
- Erman, F.; Tuzcu, M.; Orhan, C.; Sahin, N.; Sahin, K. Effect of lycopene against cisplatin-induced acute renal injury in rats: Organic anion and cation transporters evaluation. Biol. Trace Elem. Res. 2014, 158, 90–95. [Google Scholar] [CrossRef]
- Hartmann, G.; Cheung, A.K.; Piquette-Miller, M. Inflammatory cytokines, but not bile acids, regulate expression of murine hepatic anion transporters in endotoxemia. J. Pharmacol. Exp. Ther. 2002, 303, 273–281. [Google Scholar] [CrossRef]
- Choi, M.K.; Kim, M.H.; Maeng, H.J.; Song, I.S. Contribution of CNT1 and ENT1 to ribavirin uptake in human hepatocytes. Arch. Pharm. Res. 2018, 38, 904–913. [Google Scholar] [CrossRef]
- Pour, N.K.; Piquette-Miller, M. Endotoxin Modulates the Expression of Renal Drug Transporters in HIV-1 Transgenic Rats. J. Pharm. Pharm. Sci. 2018, 21, 117s–129s. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Drug Transporter Name | Drug Substrate |
|---|---|---|
| ABCB1 | P-glycoprotein (P-gp) | Acebutolol, Apixaban, Betamethasone, Cimetidine, Ciprofloxacin, Cortisol, Diazoxide, Digoxin, Edoxaban, Ephedrine, Fimasartan, Fluvastatin, Furosemide, Glyburide, Lamivudine, Maraviroc, Metoclopramide, Rosuvastatin, Sitagliptin, Zidovudine |
| ABCC2 | Multidrug resistance-associated protein 2 (MRP2) | Actinomycin D, Ampicillin, Anagliptin, Azithromycin, Carvedilol, Cisplatin, Darunavir, Eluxadoline, Enalapril, Fexofenadine, Gemfibrozil, Indinavir, Methotrexate, Morphine, Ofloxacin, Paclitaxel, Paracetamol, Pitavastatin, Rosuvastatin, Talinolol, Tenofovir disoproxil fumarate, Thyroxine |
| ABCC4 | Multidrug resistance-associated protein 4 (MRP4) | Abacavir, Adefovir, Alacepril, Amoxicillin, Ampicillin, Anagliptin, Baclofen, Bumetanide, Captopril, Ceftazidime, Cisplatin, Gemfibrozil, Imatinib, Mefenamic acid, Nalidixic acid, Nateglinide, Norfloxacin, Penicillin, Pravastatin, Rosuvastatin, Tenofovir disoproxil fumarate, Tetracycline, Zidovudine |
| SLC22A2 | Organic cation transporter 2 (OCT2) | 6β-hydroxycortisol, Agmatine, Ceftobiprole, Dofetilide, Enoxacin, Entecavir, Gemifloxacin, Gentamycin, Glycopyrrolate, Ibrutinib, Mirabegron, Nadolol, Picoplatin, Pramipexole, Veliparib |
| SLC22A4 | Organic cation/carnitine transporter 1 (OCTN1) | Amisulpride, Doxorubicin, Ergothioneine, Gabapentin, Hydroxyurea, Metformin, Mitoxantrone, Phenformin |
| SLC22A5 | Organic cation/carnitine transporter 1 (OCTN2) | Amisulpride, FAMT, Hydroxyurea, Pramipexole |
| SLC47A1 | Multidrug and toxin extrusion protein 1 (MATE1) | 6β-hydroxycortisol, Acyclovir, Alacepril, Dofetilide, Dopamine, Enoxacin, Ethambutol, Flecainide, Flutamide, Ganciclovir, Gemi-floxacin, Glycopyrrolate, Hydroxychloroquine, Lamivudine, Mesna, Tenofovir disoproxil fumarate, Tipiracil, Triamterene |
| SLC22A6 | Organic anion transporter 1 (OAT1) | 6-Mercaptopurine, Anagliptin, Avibactam, Cefmetazole, Cefoxitin, Cisplatin, Fimasartan, Fimasartan, Fleroxacin, Ganciclovir, Imipenem, Meropenem, Methotrexate, Norfloxacin, Ofloxacin, Oseltamivir, Oxazepam, Quinapril, Sacubitril, Tazobactam, Temocaprilate |
| SLC22A8 | Organic anion transporter 3 (OAT3) | 6β-hydroxycortisol, Abacavir, Acetylsalicylate, Allopurinol, Anagliptin, Bezafibrate, Captopril, Cidofovir, Cilostazol, Cisplatin, Edaravone, Eluxadoline, Empagliflozin, Entecavir, Fexofenadine, Fleroxacin, Ganciclovir, Glyburide, Imipenem, Lamotrigine, Leucovorin, Mesna, Morinidazole, Oseltamivir, Pemetrexed, Pitavastatin, Ranitidine, Safinamide, Sitagliptin, Tazobactam, Temocaprilate, Tetracycline, Topiramate, Topotecan, Valaciclovir |
| SLCO4C1 | Organic anion transporter polypeptide 4C1 (OATP4C1) | Estrone sulfate, Methotrexate, Sitagliptin |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karimian Pour, N.; McColl, E.R.; Piquette-Miller, M. Impact of Viral Inflammation on the Expression of Renal Drug Transporters in Pregnant Rats. Pharmaceutics 2019, 11, 624. https://doi.org/10.3390/pharmaceutics11120624
Karimian Pour N, McColl ER, Piquette-Miller M. Impact of Viral Inflammation on the Expression of Renal Drug Transporters in Pregnant Rats. Pharmaceutics. 2019; 11(12):624. https://doi.org/10.3390/pharmaceutics11120624
Chicago/Turabian StyleKarimian Pour, Navaz, Eliza R. McColl, and Micheline Piquette-Miller. 2019. "Impact of Viral Inflammation on the Expression of Renal Drug Transporters in Pregnant Rats" Pharmaceutics 11, no. 12: 624. https://doi.org/10.3390/pharmaceutics11120624
APA StyleKarimian Pour, N., McColl, E. R., & Piquette-Miller, M. (2019). Impact of Viral Inflammation on the Expression of Renal Drug Transporters in Pregnant Rats. Pharmaceutics, 11(12), 624. https://doi.org/10.3390/pharmaceutics11120624