
Latency, Integration, and Reactivation of Human Herpesvirus-6


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. The Discovery of HHV-6
2. Biological Differences between HHV-6A and HHV-6B
3. Epidemiology of HHV-6
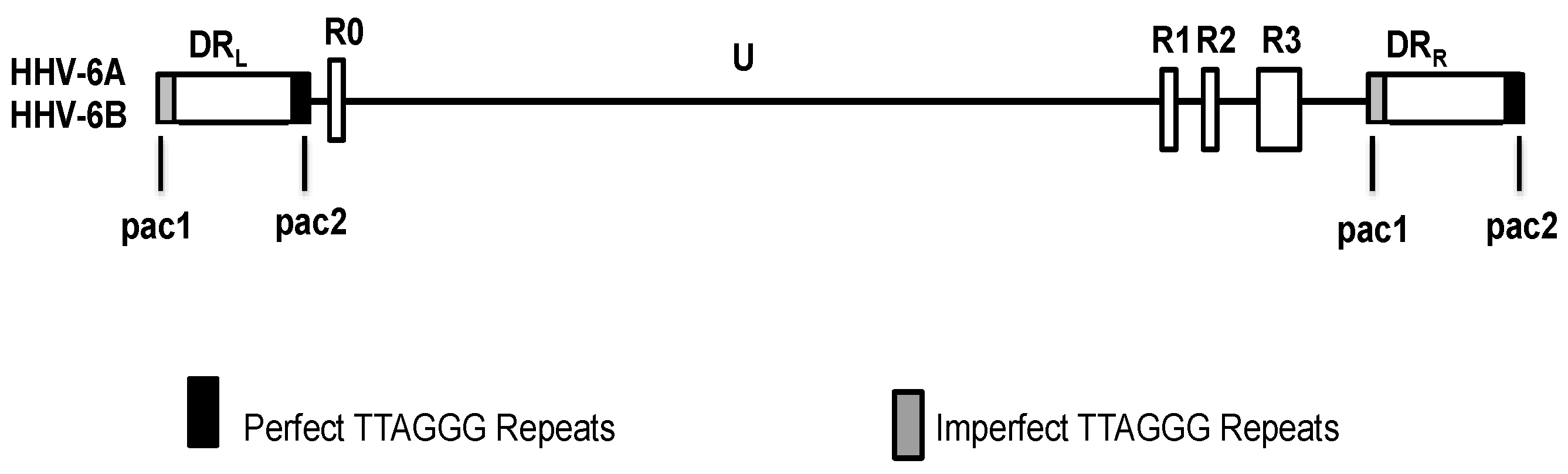
4. HHV-6 Genome Structure
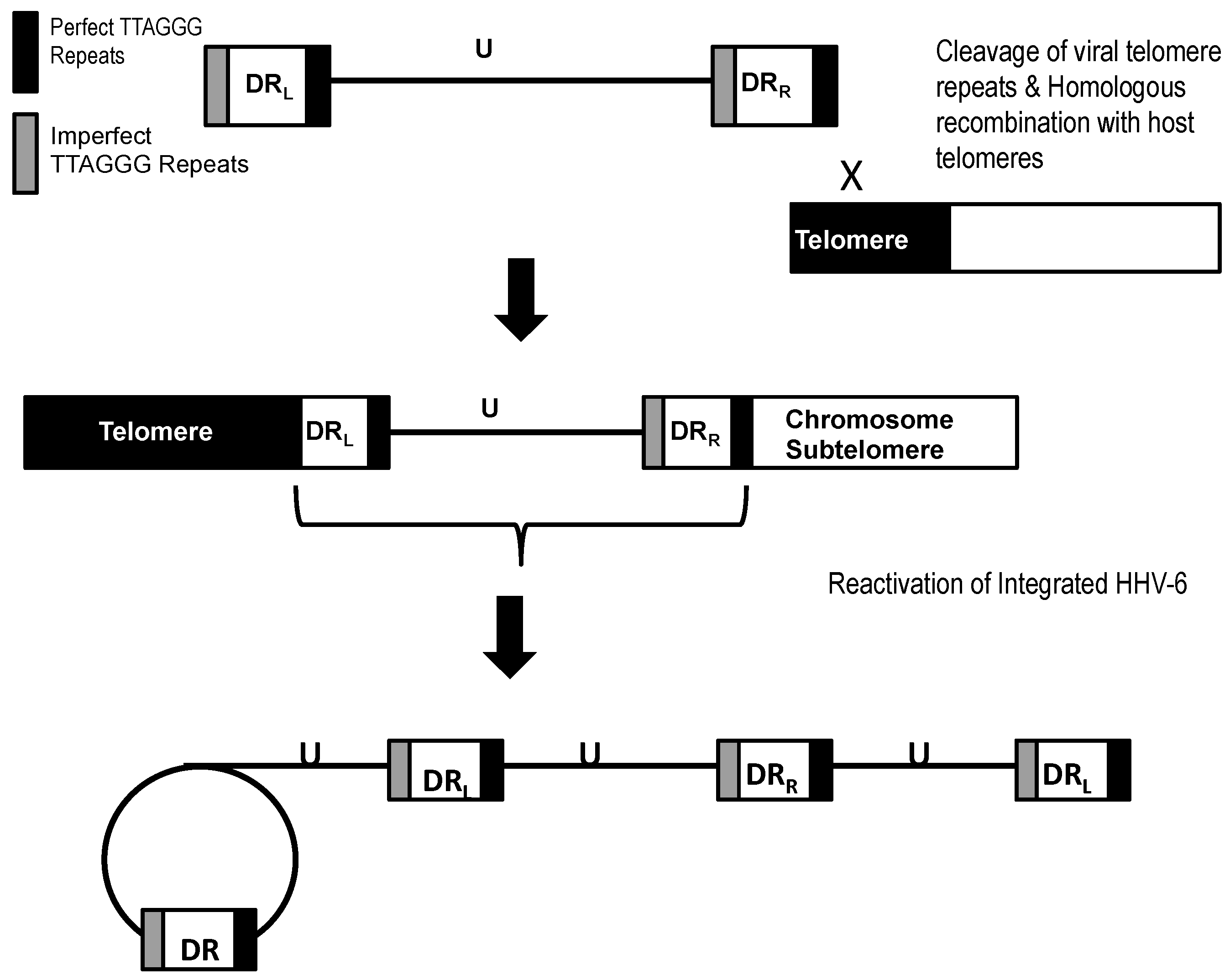
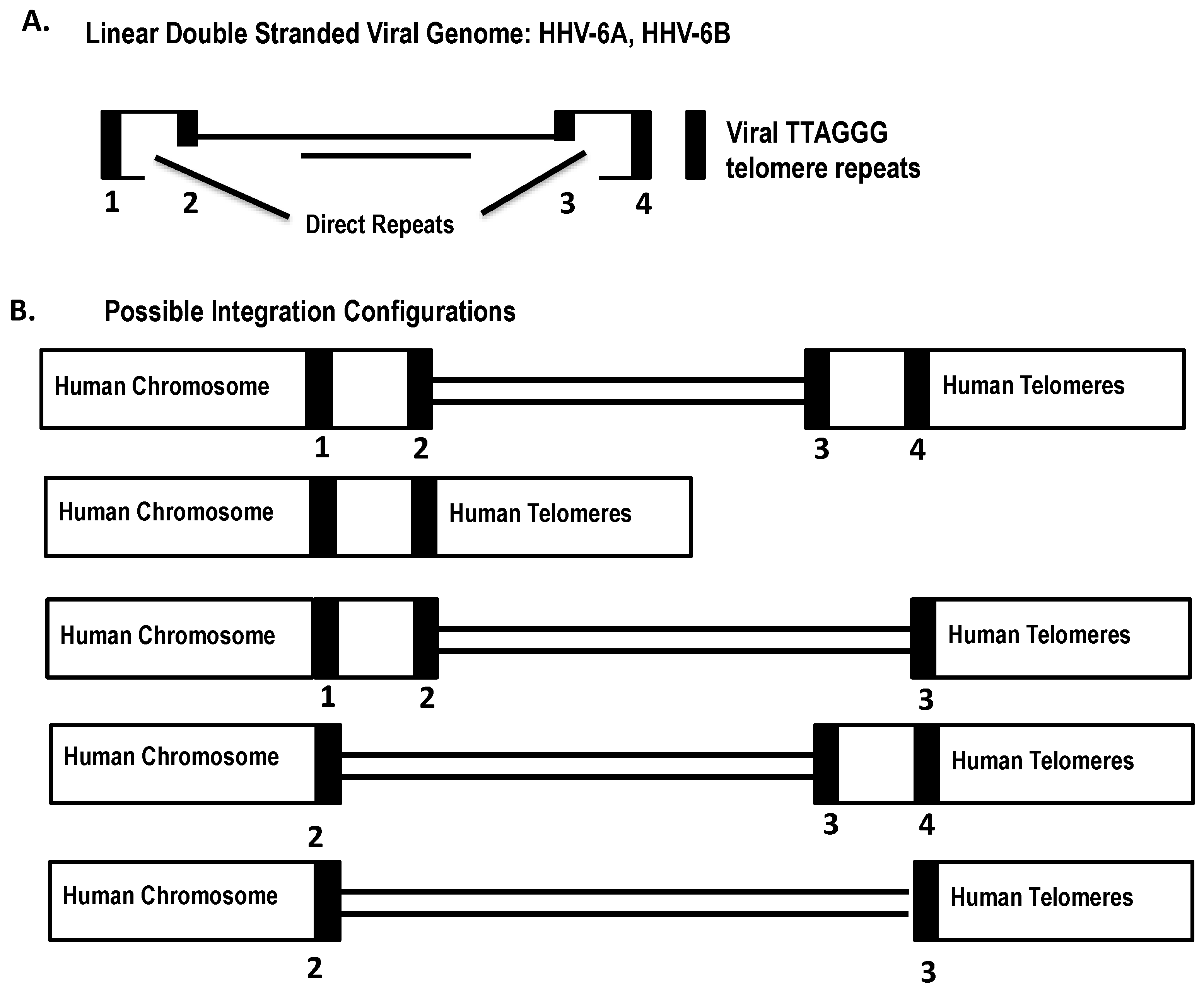
5. Integration of HHV-6 in Telomeres during Latency
6. Structure and Orientation of the Telomere-Integrated Viral Genome
7. Role of HHV-6 Telomeres and U94 in Integration
8. In Vitro Reactivation of Integrated HHV-6 from Infected Cells Harboring Latent HHV-6
9. Superinfection of iciHHV-6 Individuals with a Second Virus and Reactivation of iciHHV-6
10. Hypomethylation of Subtelomere Regions, HHV-6B Integration and Pathogenesis
11. Future of the Field of HHV-6
Conflicts of Interest
References
- Salahuddin, S.Z.; Ablashi, D.V.; Markham, P.D.; Josephs, S.F.; Sturzenegger, S.; Kaplan, M.; Halligan, G.; Biberfeld, P.; Wong-Staal, F.; Kramarsky, B.; et al. Isolation of a new virus, HBLV, in patients with lymphoproliferative disorders. Science 1986, 234, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Lusso, P.; Malnati, M.; De Maria, A.; Balotta, C.; DeRocco, S.E.; Markham, P.D.; Gallo, R.C. Productive infection of CD4+ and CD8+ mature human T cell populations and clones by human herpesvirus 6. Transcriptional down-regulation of CD3. J. Immunol. 1991, 147, 685–691. [Google Scholar] [PubMed]
- Lusso, P.; Markham, P.D.; Tschachler, E.; di Marzo Veronese, F.; Salahuddin, S.Z.; Ablashi, D.V.; Pahwa, S.; Krohn, K.; Gallo, R.C. In vitro cellular tropism of human B-lymphotropic virus (human herpesvirus-6). J. Exp. Med. 1988, 167, 1659–1670. [Google Scholar] [CrossRef]
- Takahashi, K.; Sonoda, S.; Higashi, K.; Kondo, T.; Takahashi, H.; Takahashi, M.; Yamanishi, K. Predominant CD4 T-lymphocyte tropism of human herpesvirus 6-related virus. J. Virol. 1989, 63, 3161–3163. [Google Scholar] [PubMed]
- Yasukawa, M.; Yakushijin, Y.; Furukawa, M.; Fujita, S. Specificity analysis of human CD4+ T-cell clones directed against human herpesvirus 6 (HHV-6), HHV-7, and human cytomegalovirus. J. Virol. 1993, 67, 6259–6264. [Google Scholar] [PubMed]
- Schirmer, E.C.; Wyatt, L.S.; Yamanishi, K.; Rodriguez, W.J.; Frenkel, N. Differentiation between two distinct classes of viruses now classified as human herpesvirus 6. Proc. Natl. Acad. Sci. USA 1991, 88, 5922–5926. [Google Scholar] [CrossRef] [PubMed]
- Ablashi, D.V.; Balachandran, N.; Josephs, S.F.; Hung, C.L.; Krueger, G.R.; Kramarsky, B.; Salahuddin, S.Z.; Gallo, R.C. Genomic polymorphism, growth properties, and immunologic variations in human herpesvirus-6 isolates. Virology 1991, 184, 545–552. [Google Scholar] [CrossRef]
- Ablashi, D.; Agut, H.; Alvarez-Lafuente, R.; Clark, D.A.; Dewhurst, S.; DiLuca, D.; Flamand, L.; Frenkel, N.; Gallo, R.; Gompels, U.A.; et al. Classification of HHV-6A and HHV-6B as distinct viruses. Arch. Virol. 2014, 159, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Liszewski, M.K.; Post, T.W.; Atkinson, J.P. Membrane cofactor protein (MCP or CD46): Newest member of the regulators of complement activation gene cluster. Annu. Rev. Immunol. 1991, 9, 431–455. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Serada, S.; Kawabata, A.; Ota, M.; Hayashi, E.; Naka, T.; Yamanishi, K.; Mori, Y. CD134 is a cellular receptor specific for human herpesvirus-6B entry. Proc. Natl. Acad. Sci. USA 2013, 110, 9096–9099. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Mori, Y. Determinants of human CD134 essential for entry of human herpesvirus 6B. J. Virol. 2015, 89, 10125–10129. [Google Scholar] [CrossRef] [PubMed]
- Jasirwan, C.; Furusawa, Y.; Tang, H.; Maeki, T.; Mori, Y. Human herpesvirus-6A gQ1 and gQ2 are critical for human CD46 usage. Microbiol. Immunol. 2014, 58, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.S.; Bundgaard, B.B.; Biltoft, M.; Rossen, L.S.; Hollsberg, P. Divergent tropism of HHV-6AGS and HHV-6BPl1 in T cells expressing different CD46 isoform patterns. Virology 2017, 502, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Achour, A.; Malet, I.; Le Gal, F.; Dehee, A.; Gautheret-Dejean, A.; Bonnafous, P.; Agut, H. Variability of gB and gH genes of human herpesvirus-6 among clinical specimens. J. Med. Virol. 2008, 80, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Ward, K.N.; Gray, J.J.; Fotheringham, M.W.; Sheldon, M.J. IgG antibodies to human herpesvirus-6 in young children: Changes in avidity of antibody correlate with time after infection. J. Med. Virol. 1993, 39, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Bates, M.; Monze, M.; Bima, H.; Kapambwe, M.; Clark, D.; Kasolo, F.C.; Gompels, U.A. Predominant human herpesvirus 6 variant a infant infections in an HIV-1 endemic region of sub-Saharan Africa. J. Med. Virol. 2009, 81, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Tembo, J.; Kabwe, M.; Chilukutu, L.; Chilufya, M.; Mwaanza, N.; Chabala, C.; Zumla, A.; Bates, M. Prevalence and risk factors for betaherpesvirus DNAemia in children >3 weeks and <2 years of age admitted to a large referral hospital in sub-saharan africa. Clin. Infect. Dis. 2015, 60, 423–431. [Google Scholar]
- Wang, F.Z.; Dahl, H.; Ljungman, P.; Linde, A. Lymphoproliferative responses to human herpesvirus-6 variant a and variant b in healthy adults. J. Med. Virol. 1999, 57, 134–139. [Google Scholar] [CrossRef]
- Jayavasu, C.; Balachandra, K.; Wongchuree, S.; Kositanont, U.; Warachit, P. The latency rate of human herpesvirus 6 (HHV6) in positive and negative human immunodeficiency virus (HIV) infection of intravenous drug users (IVDU). Asian Pac. J. Allergy Immunol. 1997, 15, 29–33. [Google Scholar] [PubMed]
- Yamanishi, K.; Okuno, T.; Shiraki, K.; Takahashi, M.; Kondo, T.; Asano, Y.; Kurata, T. Identification of human herpesvirus-6 as a causal agent for exanthem subitum. Lancet 1988, 1, 1065–1067. [Google Scholar] [CrossRef]
- Tanaka, K.; Kondo, T.; Torigoe, S.; Okada, S.; Mukai, T.; Yamanishi, K. Human herpesvirus 7: Another causal agent for roseola (exanthem subitum). J Pediatr 1994, 125, 1–5. [Google Scholar] [CrossRef]
- Torigoe, S.; Kumamoto, T.; Koide, W.; Taya, K.; Yamanishi, K. Clinical manifestations associated with human herpesvirus 7 infection. Arch Dis Child 1995, 72, 518–519. [Google Scholar] [CrossRef] [PubMed]
- Ablashi, D.V.; Lapps, W.; Kaplan, M.; Whitman, J.E.; Richert, J.R.; Pearson, G.R. Human herpesvirus-6 (HHV-6) infection in multiple sclerosis: A preliminary report. Mult. Scler. 1998, 4, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Ben-Fredj, N.; Ben-Selma, W.; Rotola, A.; Nefzi, F.; Benedetti, S.; Frih-Ayed, M.; Di Luca, D.; Aouni, M.; Caselli, E. Prevalence of human herpesvirus U94/Rep antibodies and DNA in tunisian multiple sclerosis patients. J. Neurovirol. 2013, 19, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Soldan, S.S.; Berti, R.; Salem, N.; Secchiero, P.; Flamand, L.; Calabresi, P.A.; Brennan, M.B.; Maloni, H.W.; McFarland, H.F.; Lin, H.C.; et al. Association of human herpes virus 6 (HHV-6) with multiple sclerosis: Increased IgM response to HHV-6 early antigen and detection of serum HHV-6 DNA. Nat. Med. 1997, 3, 1394–1397. [Google Scholar] [CrossRef] [PubMed]
- Ablashi, D.V.; Eastman, H.B.; Owen, C.B.; Roman, M.M.; Friedman, J.; Zabriskie, J.B.; Peterson, D.L.; Pearson, G.R.; Whitman, J.E. Frequent HHV-6 reactivation in multiple sclerosis (MS) and chronic fatigue syndrome (CFS) patients. J. Clin. Virol. 2000, 16, 179–191. [Google Scholar] [CrossRef]
- Komaroff, A.L. Is human herpesvirus-6 a trigger for chronic fatigue syndrome? J. Clin. Virol. 2006, 37 (Suppl. S1), S39–S46. [Google Scholar] [CrossRef]
- Montoya, J.G.; Neely, M.N.; Gupta, S.; Lunn, M.R.; Loomis, K.S.; Pritchett, J.C.; Polsky, B.; Medveczky, P.G. Antiviral therapy of two patients with chromosomally-integrated human herpesvirus-6A presenting with cognitive dysfunction. J. Clin. Virol. 2012, 55, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Isegawa, Y.; Mukai, T.; Nakano, K.; Kagawa, M.; Chen, J.; Mori, Y.; Sunagawa, T.; Kawanishi, K.; Sashihara, J.; Hata, A.; et al. Comparison of the complete DNA sequences of human herpesvirus 6 variants A and B. J. Virol. 1999, 73, 8053–8063. [Google Scholar] [PubMed]
- Gompels, U.A.; Macaulay, H.A. Characterization of human telomeric repeat sequences from human herpesvirus 6 and relationship to replication. J. Gen. Virol. 1995, 76 Pt 2, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, G.; Dambaugh, T.R.; Stamey, F.R.; Dewhurst, S.; Inoue, N.; Pellett, P.E. Human herpesvirus 6B genome sequence: Coding content and comparison with human herpesvirus 6A. J. Virol. 1999, 73, 8040–8052. [Google Scholar] [PubMed]
- Thomson, B.J.; Dewhurst, S.; Gray, D. Structure and heterogeneity of the a sequences of human herpesvirus 6 strain variants U1102 and Z29 and identification of human telomeric repeat sequences at the genomic termini. J. Virol. 1994, 68, 3007–3014. [Google Scholar] [PubMed]
- Gravel, A.; Hall, C.B.; Flamand, L. Sequence analysis of transplacentally acquired human herpesvirus 6 DNA is consistent with transmission of a chromosomally integrated reactivated virus. J. Infect. Dis. 2013, 207, 1585–1589. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Kondo, T.; Okuno, T.; Takahashi, M.; Yamanishi, K. Latent human herpesvirus 6 infection of human monocytes/macrophages. J. Gen. Virol. 1991, 72 Pt 6, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Luppi, M.; Barozzi, P.; Morris, C.; Maiorana, A.; Garber, R.; Bonacorsi, G.; Donelli, A.; Marasca, R.; Tabilio, A.; Torelli, G. Human herpesvirus 6 latently infects early bone marrow progenitors in vivo. J. Virol. 1999, 73, 754–759. [Google Scholar] [PubMed]
- Arbuckle, J.H.; Medveczky, M.M.; Luka, J.; Hadley, S.H.; Luegmayr, A.; Ablashi, D.; Lund, T.C.; Tolar, J.; De Meirleir, K.; Montoya, J.G.; et al. The latent human herpesvirus-6A genome specifically integrates in telomeres of human chromosomes in vivo and in vitro. Proc. Natl. Acad. Sci. USA 2010, 107, 5563–5568. [Google Scholar] [CrossRef] [PubMed]
- Arbuckle, J.H.; Pantry, S.N.; Medveczky, M.M.; Prichett, J.; Loomis, K.S.; Ablashi, D.; Medveczky, P.G. Mapping the telomere integrated genome of human herpesvirus 6A and 6B. Virology 2013, 442, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Asano, Y.; Akimoto, S.; Ozaki, T.; Iwasaki, T.; Kurata, T.; Goshima, F.; Nishiyama, Y. Latent infection of human herpesvirus 6 in astrocytoma cell line and alteration of cytokine synthesis. J. Med. Virol. 2002, 66, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Ahlqvist, J.; Fotheringham, J.; Akhyani, N.; Yao, K.; Fogdell-Hahn, A.; Jacobson, S. Differential tropism of human herpesvirus 6 (HHV-6) variants and induction of latency by HHV-6A in oligodendrocytes. J. Neurovirol. 2005, 11, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Luppi, M.; Marasca, R.; Barozzi, P.; Ferrari, S.; Ceccherini-Nelli, L.; Batoni, G.; Merelli, E.; Torelli, G. Three cases of human herpesvirus-6 latent infection: Integration of viral genome in peripheral blood mononuclear cell DNA. J. Med. Virol. 1993, 40, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Tanaka-Taya, K.; Sashihara, J.; Kurahashi, H.; Amo, K.; Miyagawa, H.; Kondo, K.; Okada, S.; Yamanishi, K. Human herpesvirus 6 (HHV-6) is transmitted from parent to child in an integrated form and characterization of cases with chromosomally integrated HHV-6 DNA. J. Med. Virol. 2004, 73, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.B.; Caserta, M.T.; Schnabel, K.; Shelley, L.M.; Marino, A.S.; Carnahan, J.A.; Yoo, C.; Lofthus, G.K.; McDermott, M.P. Chromosomal integration of human herpesvirus 6 is the major mode of congenital human herpesvirus 6 infection. Pediatrics 2008, 122, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Daibata, M.; Taguchi, T.; Nemoto, Y.; Taguchi, H.; Miyoshi, I. Inheritance of chromosomally integrated human herpesvirus 6 DNA. Blood 1999, 94, 1545–1549. [Google Scholar] [PubMed]
- Leong, H.N.; Tuke, P.W.; Tedder, R.S.; Khanom, A.B.; Eglin, R.P.; Atkinson, C.E.; Ward, K.N.; Griffiths, P.D.; Clark, D.A. The prevalence of chromosomally integrated human herpesvirus 6 genomes in the blood of UK blood donors. J. Med. Virol. 2007, 79, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Ljungman, P.; Wang, F.Z.; Clark, D.A.; Emery, V.C.; Remberger, M.; Ringden, O.; Linde, A. High levels of human herpesvirus 6 DNA in peripheral blood leucocytes are correlated to platelet engraftment and disease in allogeneic stem cell transplant patients. Br. J. Haematol. 2000, 111, 774–781. [Google Scholar] [PubMed]
- Clark, D.A.; Nacheva, E.P.; Leong, H.N.; Brazma, D.; Li, Y.T.; Tsao, E.H.; Buyck, H.C.; Atkinson, C.E.; Lawson, H.M.; Potter, M.N.; et al. Transmission of integrated human herpesvirus 6 through stem cell transplantation: Implications for laboratory diagnosis. J. Infect. Dis. 2006, 193, 912–916. [Google Scholar] [CrossRef] [PubMed]
- Ward, K.N.; Leong, H.N.; Nacheva, E.P.; Howard, J.; Atkinson, C.E.; Davies, N.W.; Griffiths, P.D.; Clark, D.A. Human herpesvirus 6 chromosomal integration in immunocompetent patients results in high levels of viral DNA in blood, sera, and hair follicles. J. Clin. Microbiol. 2006, 44, 1571–1574. [Google Scholar] [CrossRef] [PubMed]
- Caserta, M.T.; Hall, C.B.; Schnabel, K.; Lofthus, G.; Marino, A.; Shelley, L.; Yoo, C.; Carnahan, J.; Anderson, L.; Wang, H. Diagnostic assays for active infection with human herpesvirus 6 (HHV-6). J. Clin. Virol. 2010, 48, 55–57. [Google Scholar] [CrossRef] [PubMed]
- Gulve, N.; Frank, C.; Klepsch, M.; Prusty, B.K. Chromosomal integration of HHV-6A during non-productive viral infection. Sci. Rep. 2017, 7, 512. [Google Scholar] [CrossRef] [PubMed]
- Ohye, T.; Inagaki, H.; Ihira, M.; Higashimoto, Y.; Kato, K.; Oikawa, J.; Yagasaki, H.; Niizuma, T.; Takahashi, Y.; Kojima, S.; et al. Dual roles for the telomeric repeats in chromosomally integrated human herpesvirus-6. Sci. Rep. 2014, 4, 4559. [Google Scholar] [CrossRef] [PubMed]
- Wallaschek, N.; Sanyal, A.; Pirzer, F.; Gravel, A.; Mori, Y.; Flamand, L.; Kaufer, B.B. The telomeric repeats of human herpesvirus 6A (HHV-6A) are required for efficient virus integration. PLoS Pathog. 2016, 12, e1005666. [Google Scholar] [CrossRef] [PubMed]
- Gravel, A.; Dubuc, I.; Wallaschek, N.; Gilbert-Girard, S.; Collin, V.; Hall-Sedlak, R.; Jerome, K.R.; Mori, Y.; Carbonneau, J.; Boivin, G.; et al. Cell culture systems to study human herpesvirus 6A/B chromosomal integration. J. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Trempe, F.; Gravel, A.; Dubuc, I.; Wallaschek, N.; Collin, V.; Gilbert-Girard, S.; Morissette, G.; Kaufer, B.B.; Flamand, L. Characterization of human herpesvirus 6A/B U94 as atpase, helicase, exonuclease and DNA-binding proteins. Nucleic Acids Res. 2015, 43, 6084–6098. [Google Scholar] [CrossRef] [PubMed]
- Thomson, B.J.; Weindler, F.W.; Gray, D.; Schwaab, V.; Heilbronn, R. Human herpesvirus 6 (HHV-6) is a helper virus for adeno-associated virus type 2 (AAV-2) and the AAV-2 rep gene homologue in HHV-6 can mediate AAV-2 DNA replication and regulate gene expression. Virology 1994, 204, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Thomson, B.J.; Efstathiou, S.; Honess, R.W. Acquisition of the human adeno-associated virus type-2 rep gene by human herpesvirus type-6. Nature 1991, 351, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Wallaschek, N.; Gravel, A.; Flamand, L.; Kaufer, B.B. The putative U94 integrase is dispensable for human herpesvirus 6 (HHV-6) chromosomal integration. J. Gen. Virol. 2016, 97, 1899–1903. [Google Scholar] [CrossRef] [PubMed]
- Gardella, T.; Medveczky, P.; Sairenji, T.; Mulder, C. Detection of circular and linear herpesvirus DNA molecules in mammalian cells by gel electrophoresis. J. Virol. 1984, 50, 248–254. [Google Scholar] [PubMed]
- Pantry, S.N.; Medveczky, M.M.; Arbuckle, J.H.; Luka, J.; Montoya, J.G.; Hu, J.; Renne, R.; Peterson, D.; Pritchett, J.C.; Ablashi, D.V.; et al. Persistent human herpesvirus-6 infection in patients with an inherited form of the virus. J. Med. Virol. 2013, 85, 1940–1946. [Google Scholar] [CrossRef] [PubMed]
- Tweedy, J.; Spyrou, M.A.; Pearson, M.; Lassner, D.; Kuhl, U.; Gompels, U.A. Complete genome sequence of germline chromosomally integrated human herpesvirus 6A and analyses integration sites define a new human endogenous virus with potential to reactivate as an emerging infection. Viruses 2016, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Engdahl, E.; Dunn, N.; Niehusmann, P.; Wideman, S.; Wipfler, P.; Becker, A.J.; Ekstrom, T.J.; Almgren, M.; Fogdell-Hahn, A. Human herpesvirus 6B induces hypomethylation on chromosome 17p13.3, correlating with increased gene expression and virus integration. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Daibata, M.; Taguchi, T.; Kubonishi, I.; Taguchi, H.; Miyoshi, I. Lymphoblastoid cell lines with integrated human herpesvirus type 6. J. Hum. Virol. 1998, 1, 475–481. [Google Scholar] [PubMed]
- Daibata, M.; Taguchi, T.; Sawada, T.; Taguchi, H.; Miyoshi, I. Chromosomal transmission of human herpesvirus 6 DNA in acute lymphoblastic leukaemia. Lancet 1998, 352, 543–544. [Google Scholar] [CrossRef]
- Daibata, M.; Taguchi, T.; Taguchi, H.; Miyoshi, I. Integration of human herpesvirus 6 in a Burkitt's lymphoma cell line. Br. J. Haematol. 1998, 102, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Nacheva, E.P.; Ward, K.N.; Brazma, D.; Virgili, A.; Howard, J.; Leong, H.N.; Clark, D.A. Human herpesvirus 6 integrates within telomeric regions as evidenced by five different chromosomal sites. J. Med. Virol. 2008, 80, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Daibata, M.; Tohyama, M.; Batchelor, J.; Hashimoto, K.; Iijima, M. Chromosomal integration of human herpesvirus 6 DNA in anticonvulsant hypersensitivity syndrome. Br. J. Dermatol. 2008, 158, 640–642. [Google Scholar] [CrossRef] [PubMed]
- Troy, S.B.; Blackburn, B.G.; Yeom, K.; Caulfield, A.K.; Bhangoo, M.S.; Montoya, J.G. Severe encephalomyelitis in an immunocompetent adult with chromosomally integrated human herpesvirus 6 and clinical response to treatment with foscarnet plus ganciclovir. Clin. Infect. Dis. 2008, 47, e93–e96. [Google Scholar] [CrossRef] [PubMed]
- Barozzi, P.; Riva, G.; Vallerini, D.; Quadrelli, C.; Lagreca, I.; Eccheli, R.; Forghieri, F.; Coluccio, V.; Maccaferri, M.; Paolini, A.; et al. Circulating functional T cells specific to human herpes virus 6 (HHV6) antigens in individuals with chromosomally integrated hhv6. Clin. Microbiol. Infect. 2016, 22, 893–895. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.A. Clinical and laboratory features of human herpesvirus 6 chromosomal integration. Clin. Microbiol. Infect. 2016, 22, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Hidalgo-Bravo, A.; Zhang, E.; Cotton, V.E.; Mendez-Bermudez, A.; Wig, G.; Medina-Calzada, Z.; Neumann, R.; Jeffreys, A.J.; Winney, B.; et al. Human telomeres that carry an integrated copy of human herpesvirus 6 are often short and unstable, facilitating release of the viral genome from the chromosome. Nucleic Acids Res. 2014, 42, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.; Luppi, M.; McDonald, M.; Barozzi, P.; Torelli, G. Fine mapping of an apparently targeted latent human herpesvirus type 6 integration site in chromosome band 17p13.3. J. Med. Virol. 1999, 58, 69–75. [Google Scholar] [CrossRef]
- Ohye, T.; Kawamura, Y.; Inagaki, H.; Yoshikawa, A.; Ihira, M.; Yoshikawa, T.; Kurahashi, H. A simple cytogenetic method to detect chromosomally integrated human herpesvirus-6. J. Virol. Methods 2016, 228, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Strenger, V.; Aberle, S.W.; Nacheva, E.P.; Urban, C. Chromosomal integration of the HHV-6 genome in a patient with nodular sclerosis Hodgkin lymphoma. Br. J. Haematol. 2013, 161, 594–595. [Google Scholar] [CrossRef] [PubMed]
- Strenger, V.; Kayser, S.; Witte, K.E.; Lassner, D.; Schwinger, W.; Jahn, G.; Urban, C.; Feuchtinger, T. Individuals with inherited chromosomally integrated human herpes virus 6 (ciHHV-6) have functionally active HHV-6 specific T-cell immunity. Clin. Microbiol. Infect. 2016, 22. [Google Scholar] [CrossRef] [PubMed]
- Torelli, G.; Barozzi, P.; Marasca, R.; Cocconcelli, P.; Merelli, E.; Ceccherini-Nelli, L.; Ferrari, S.; Luppi, M. Targeted integration of human herpesvirus 6 in the p arm of chromosome 17 of human peripheral blood mononuclear cells in vivo. J. Med. Virol. 1995, 46, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Yagasaki, H.; Shichino, H.; Shimizu, N.; Ohye, T.; Kurahashi, H.; Yoshikawa, T.; Takahashi, S. Nine-year follow-up in a child with chromosomal integration of human herpesvirus 6 transmitted from an unrelated donor through the Japan marrow donor program. Transpl. Infect. Dis. 2015, 17, 160–161. [Google Scholar] [CrossRef] [PubMed]
- Gravel, A.; Dubuc, I.; Morissette, G.; Sedlak, R.H.; Jerome, K.R.; Flamand, L. Inherited chromosomally integrated human herpesvirus 6 as a predisposing risk factor for the development of angina pectoris. Proc. Natl. Acad. Sci. USA 2015, 112, 8058–8063. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.A.; Magaret, A.S.; Hall-Sedlak, R.; Mikhaylova, A.; Huang, M.L.; Sandmaier, B.M.; Hansen, J.A.; Jerome, K.R.; Zerr, D.M.; Boeckh, M. Outcomes of hematopoietic cell transplantation using donors or recipients with inherited chromosomally integrated HHV-6. Blood 2017. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pantry, S.N.; Medveczky, P.G. Latency, Integration, and Reactivation of Human Herpesvirus-6. Viruses 2017, 9, 194. https://doi.org/10.3390/v9070194
Pantry SN, Medveczky PG. Latency, Integration, and Reactivation of Human Herpesvirus-6. Viruses. 2017; 9(7):194. https://doi.org/10.3390/v9070194
Chicago/Turabian StylePantry, Shara N., and Peter G. Medveczky. 2017. "Latency, Integration, and Reactivation of Human Herpesvirus-6" Viruses 9, no. 7: 194. https://doi.org/10.3390/v9070194
APA StylePantry, S. N., & Medveczky, P. G. (2017). Latency, Integration, and Reactivation of Human Herpesvirus-6. Viruses, 9(7), 194. https://doi.org/10.3390/v9070194