
Virus Resistance Is Not Costly in a Marine Alga Evolving under Multiple Environmental Stressors


Abstract
:1. Introduction
2. Materials and Methods
2.1. Susceptible and Resistant Lines
2.2. Culturing Conditions
2.3. Testing RP Lines for Viral Production
2.4. Testing Resistance Type Using OtV5 Inoculation
2.5. Population Growth Rates, Cell Size and Cell Chlorophyll Content after Evolution
2.6. Competition Assay
3. Results
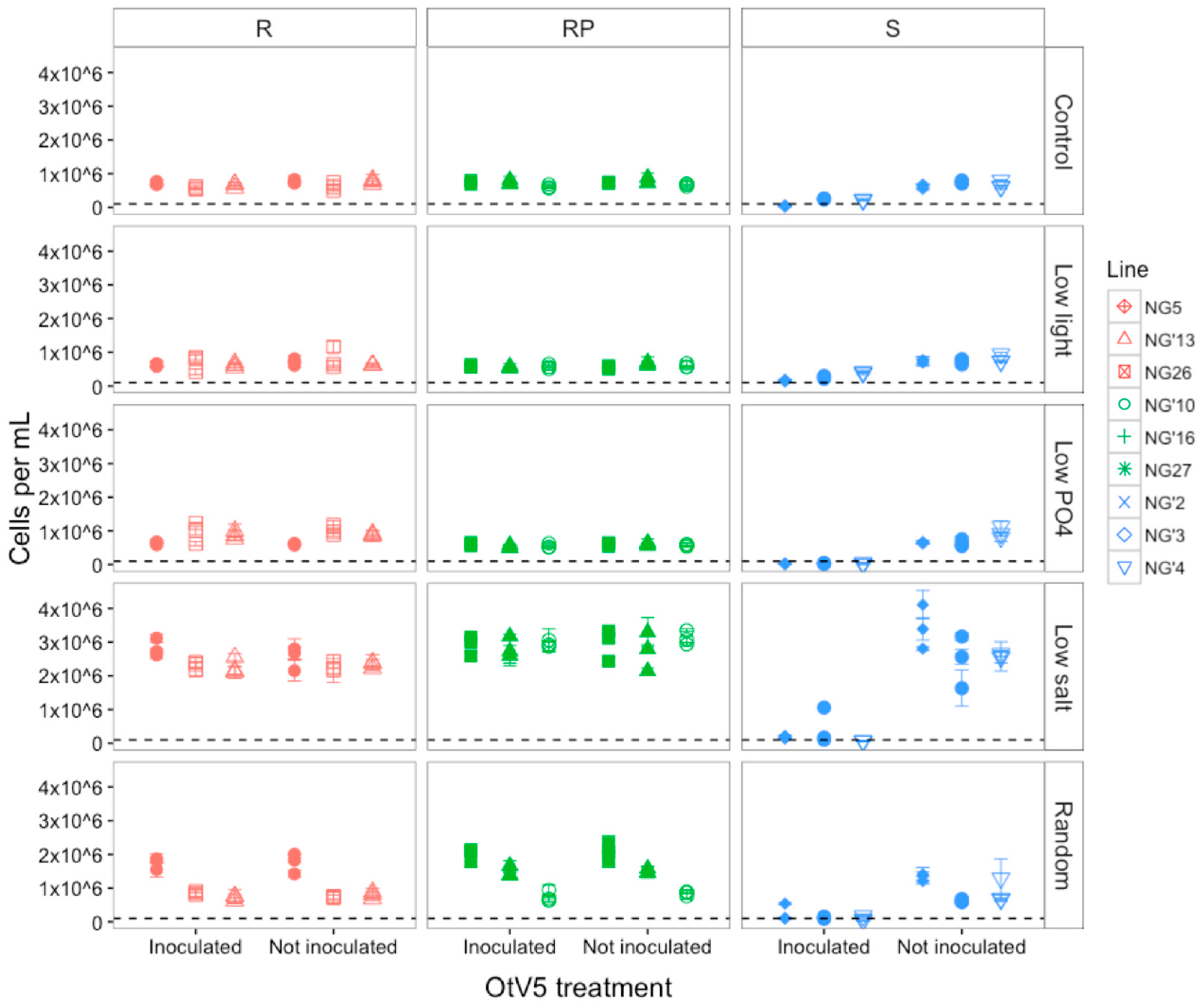
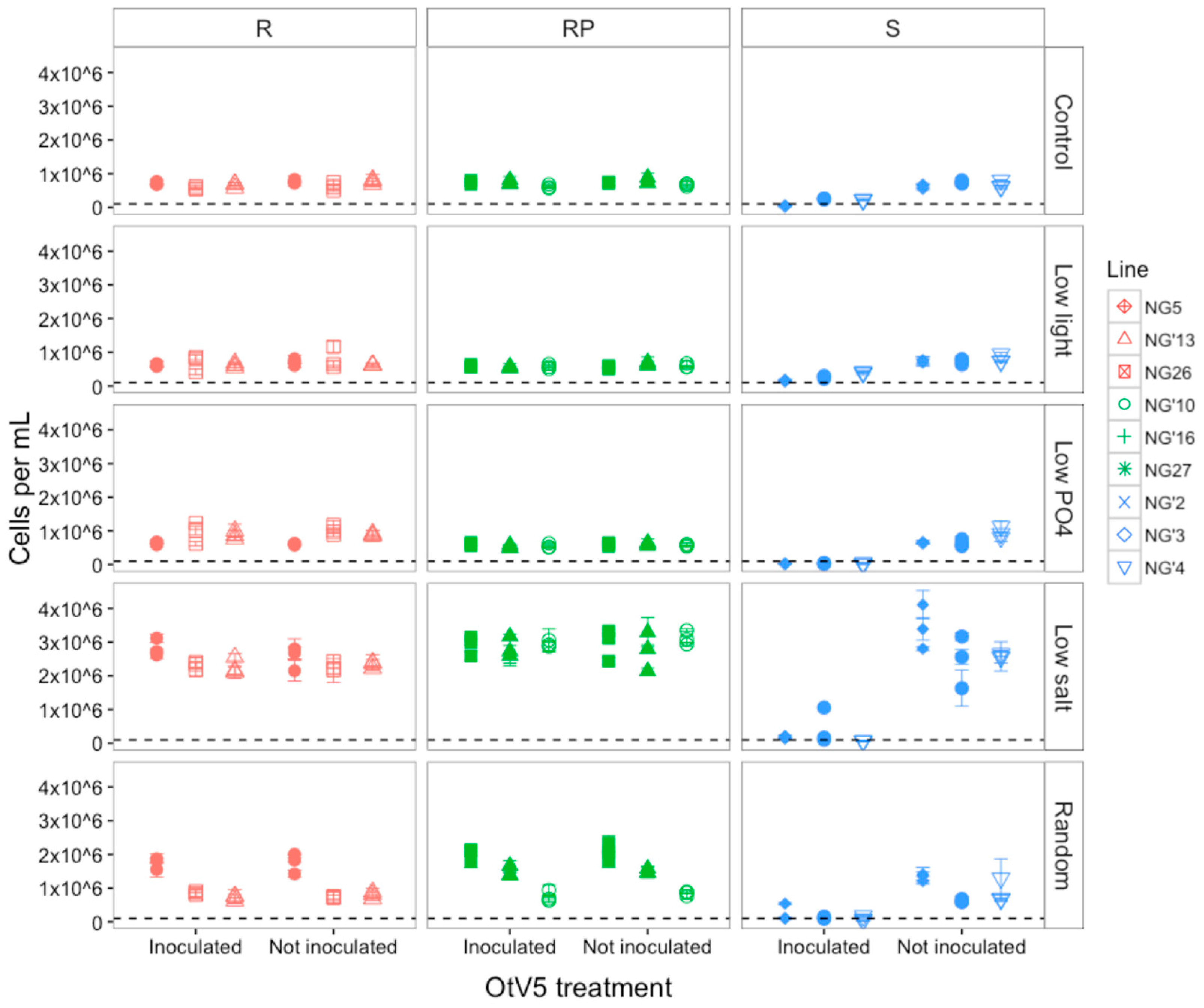
3.1. Susceptibility to OtV5 after Evolution
3.1.1. Host Resistance Type Was Maintained during Evolution
3.1.2. OtV5-Mediated Lysis Decreased in Susceptible Populations
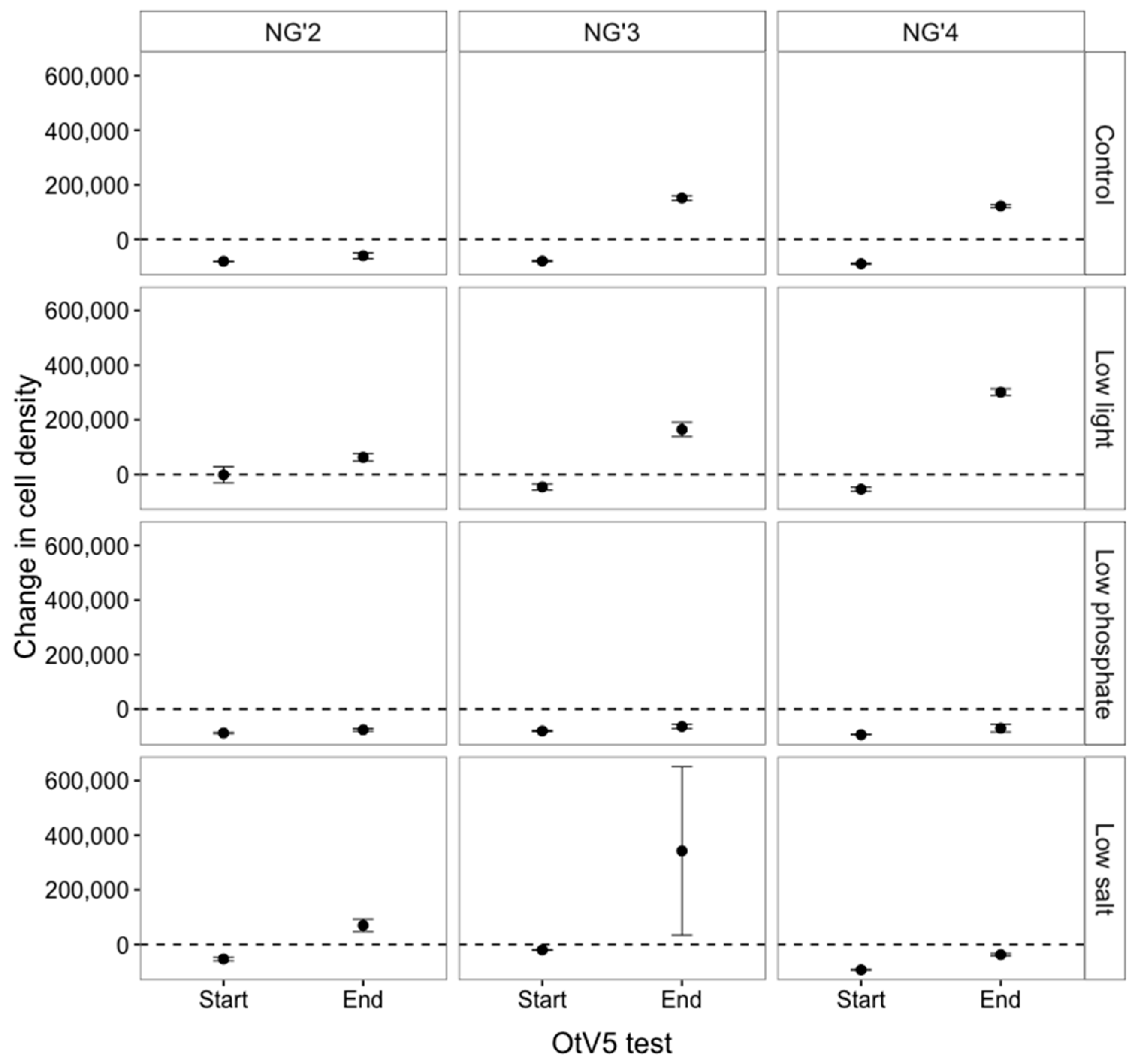
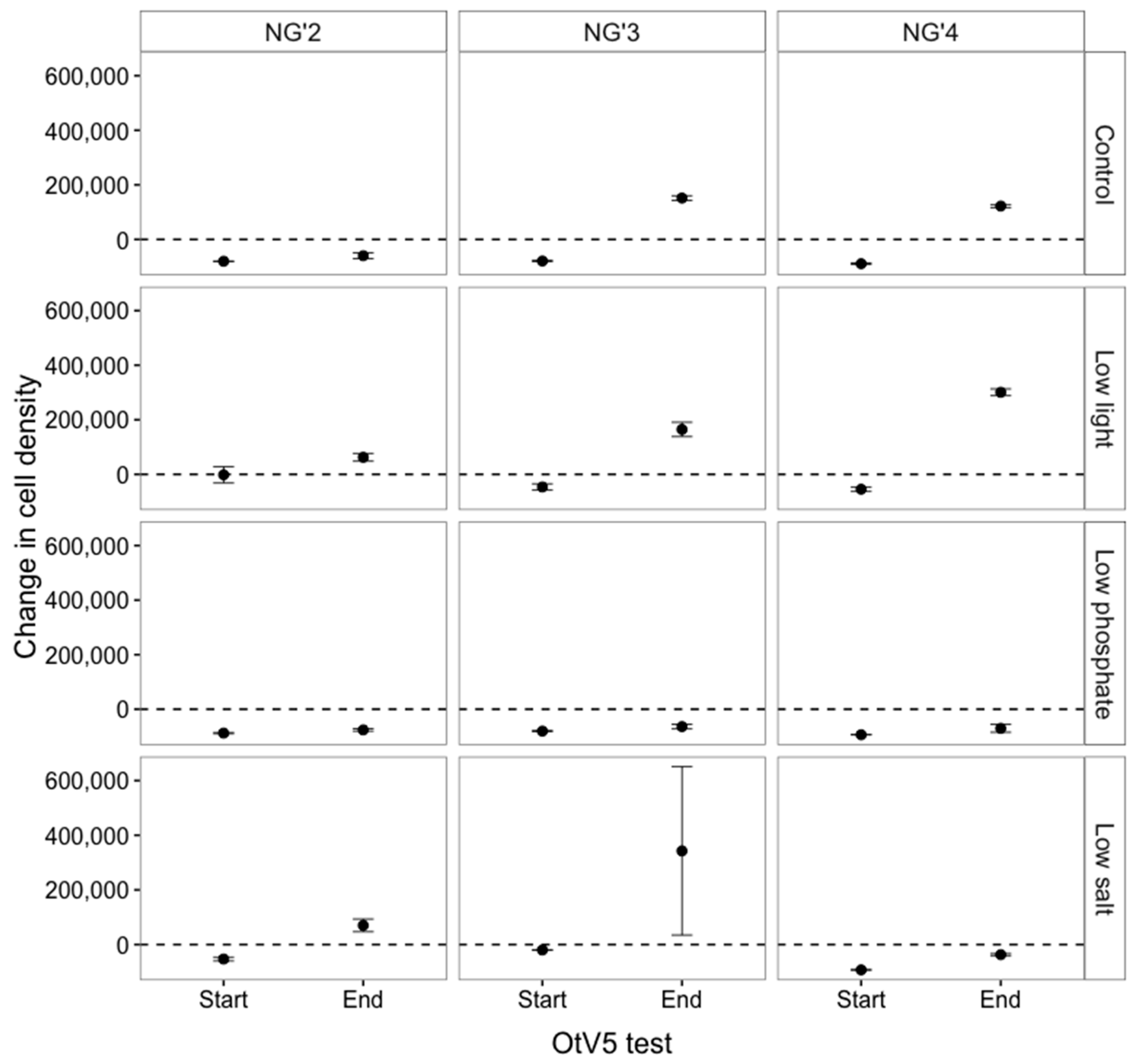
3.1.3. RPs Stopped Producing Viruses Early in the Evolution Experiment
3.2. Changes in Trait Values after Evolution
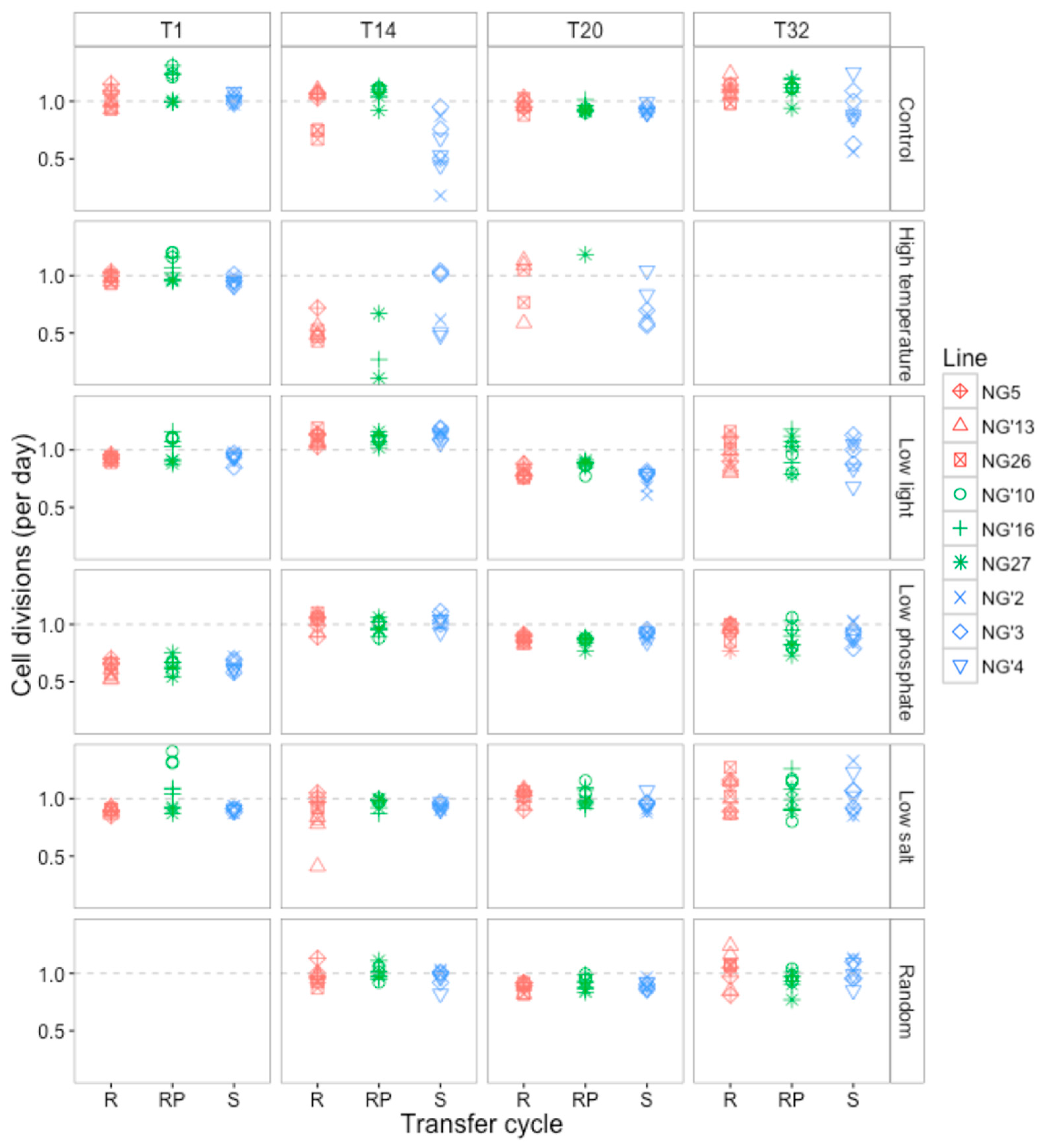
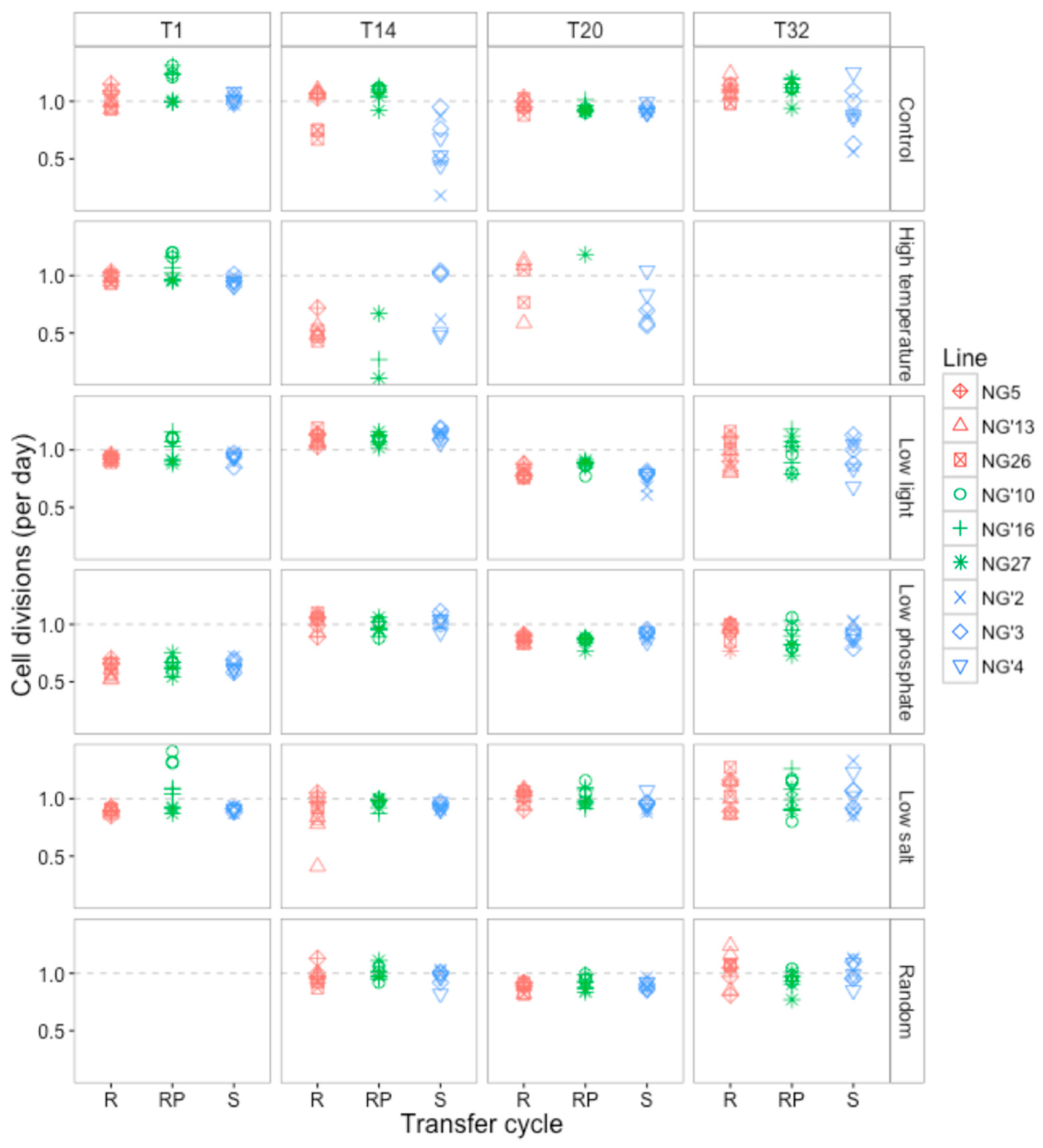
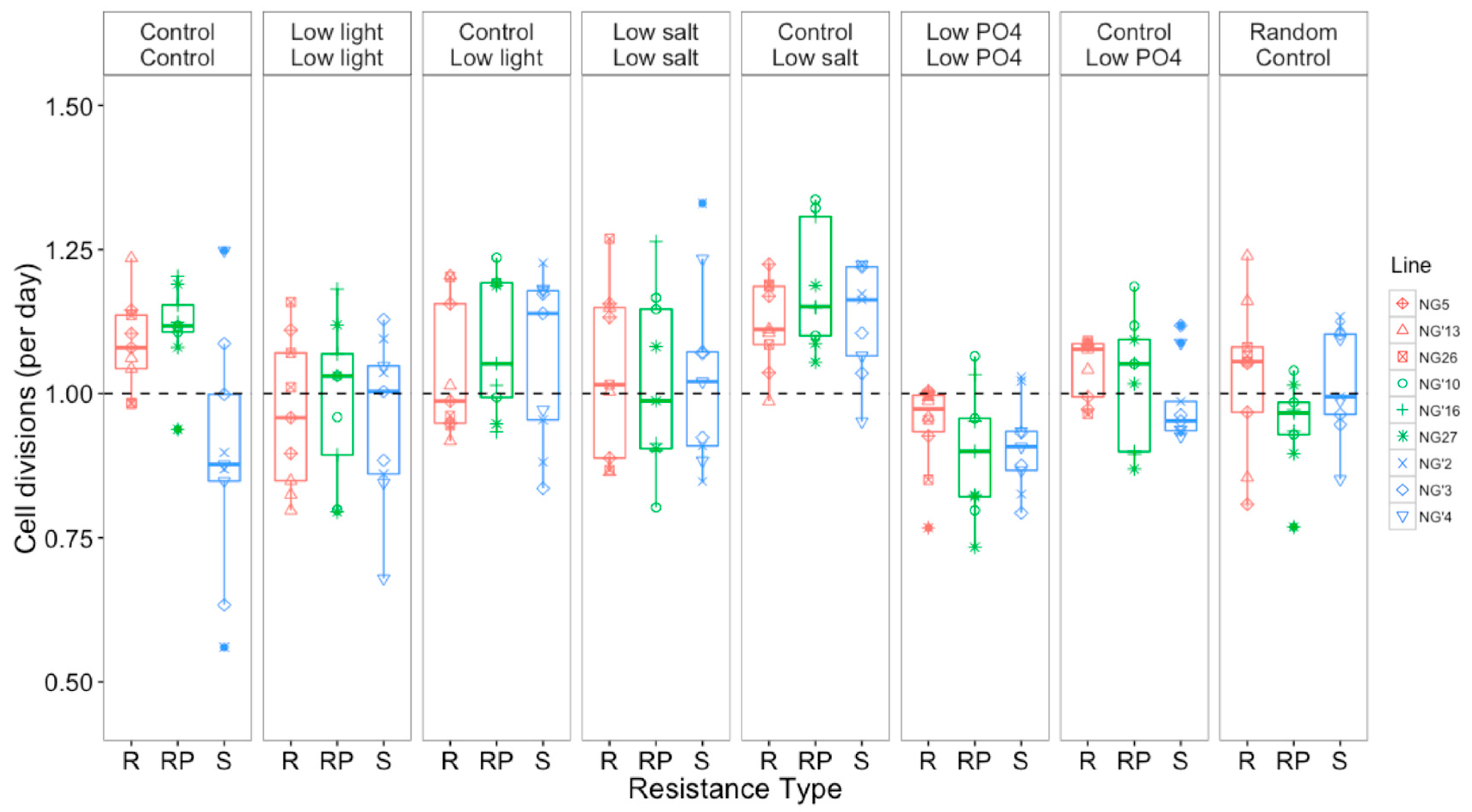
3.2.1. Changes in Cell Division Rate and Population Persistence during the Selection Experiment
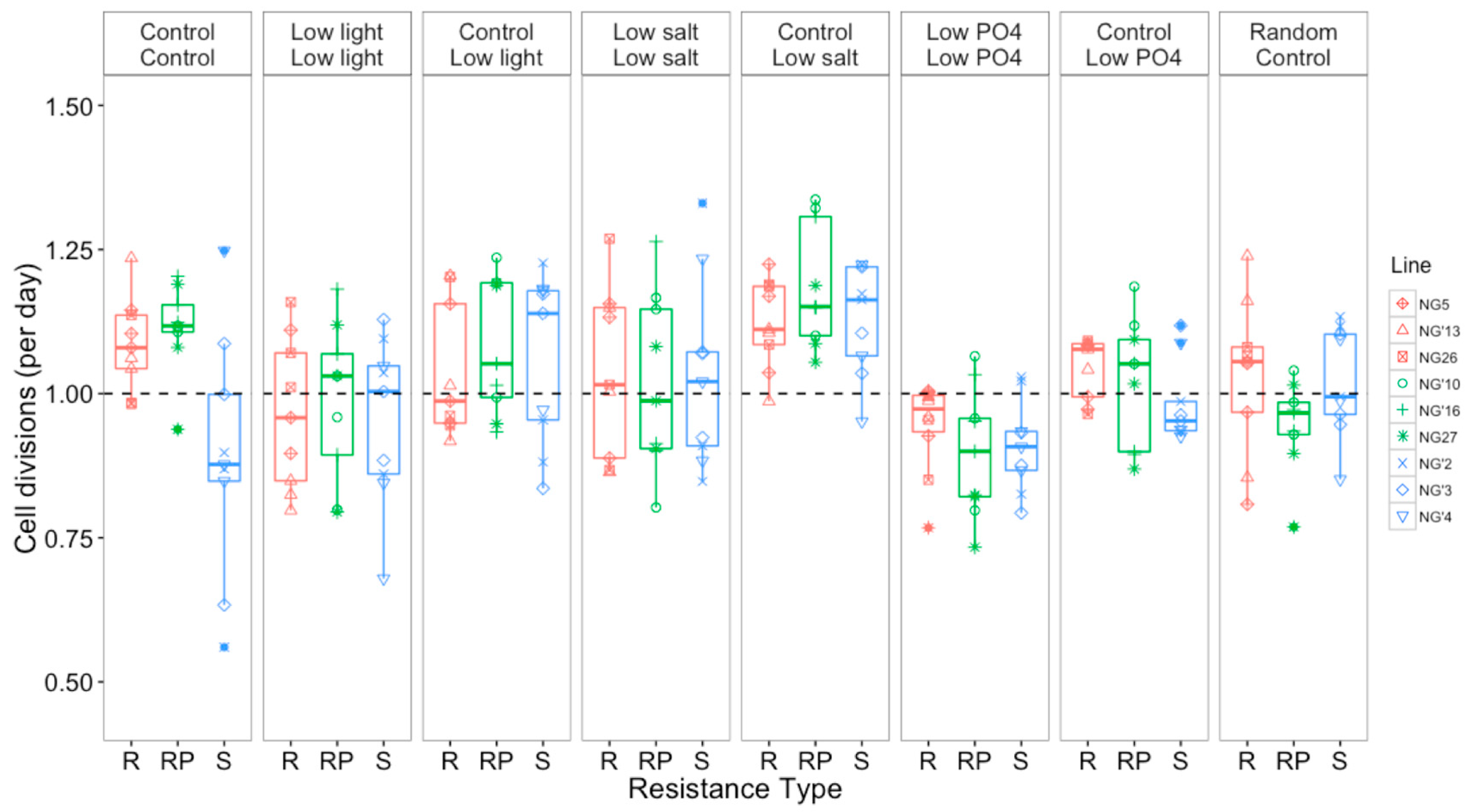
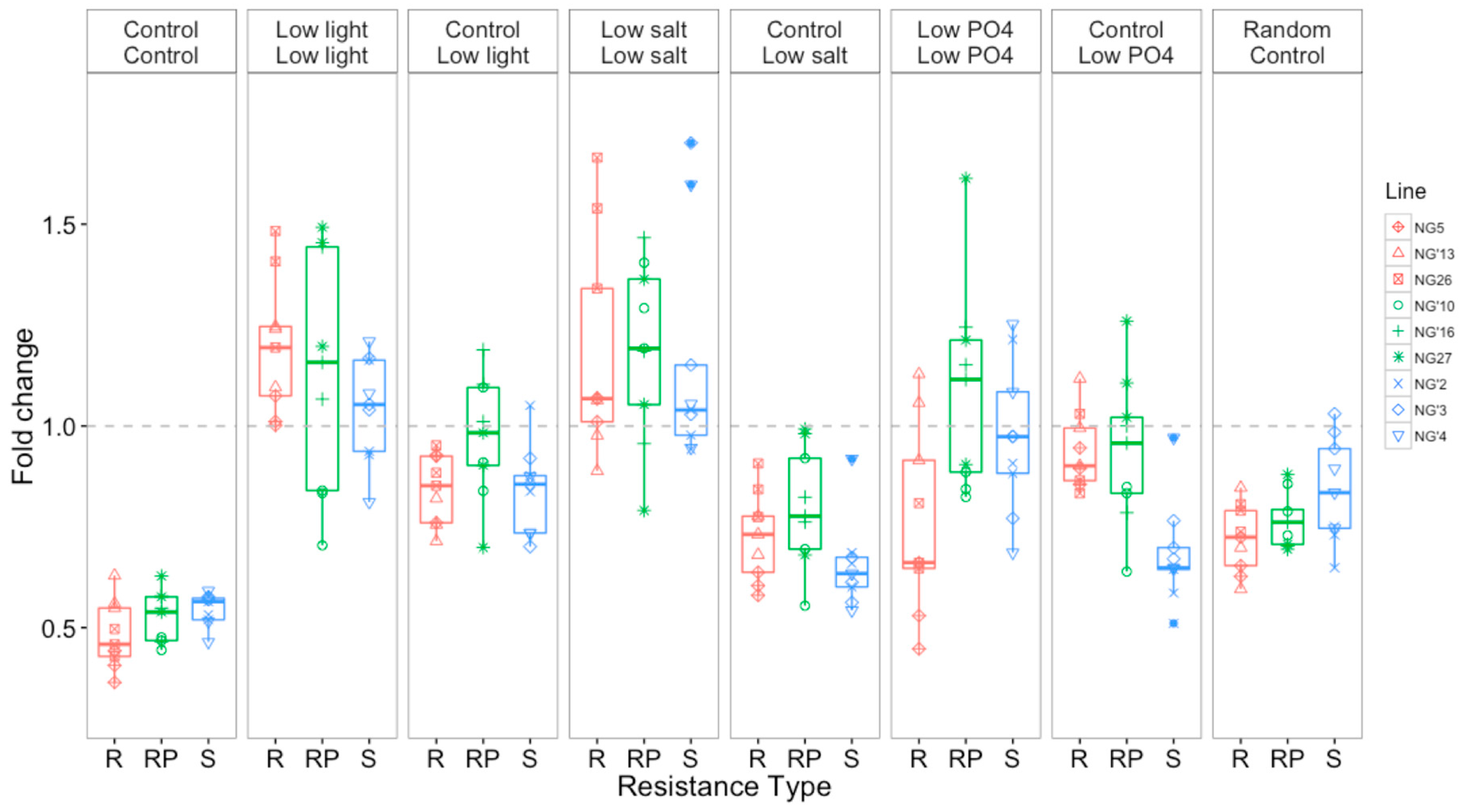
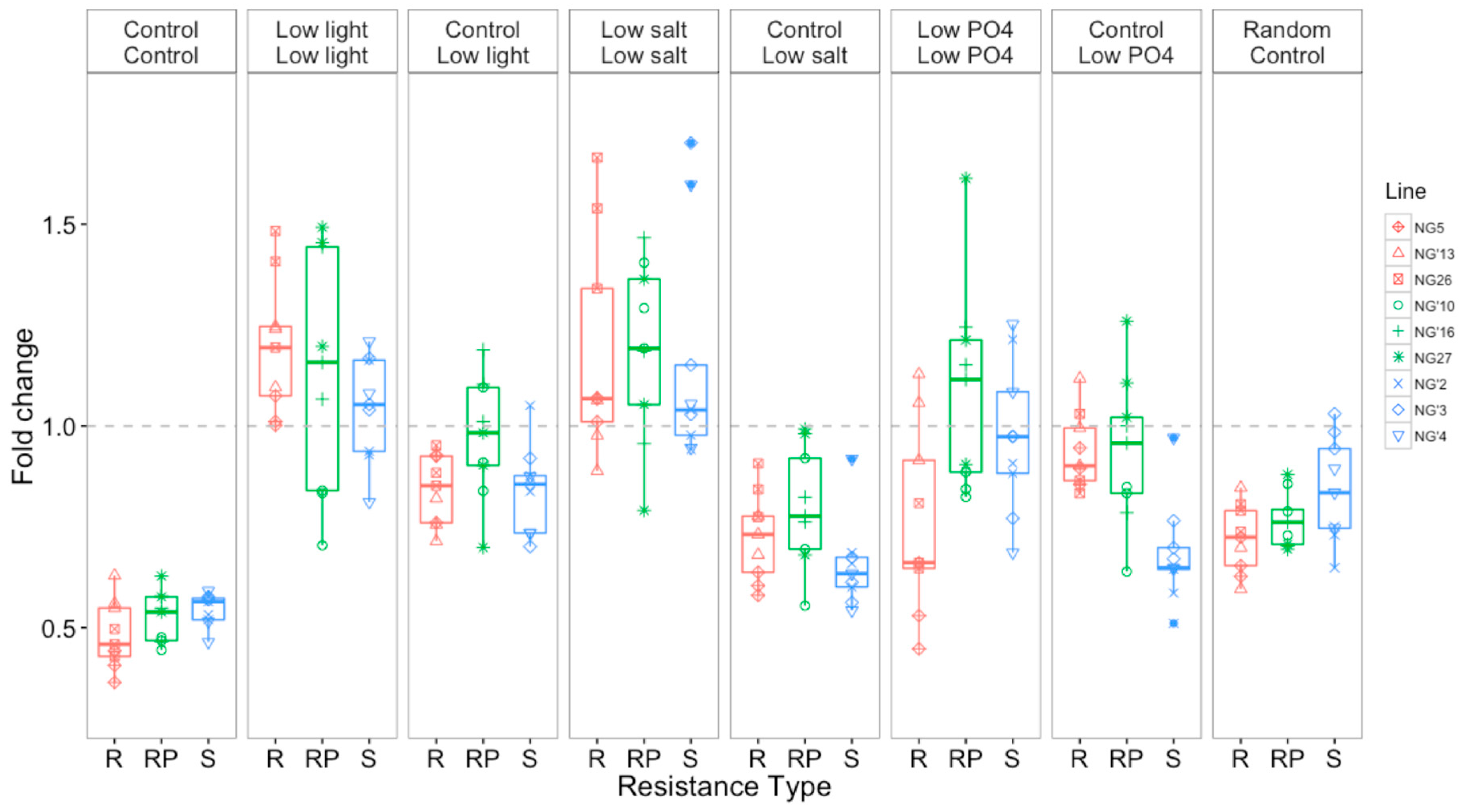
3.2.2. Growth Rates Varied with Selection Environment and Assay Environment after Evolution
3.2.3. Resistance Type Affected Cell Size and Chlorophyll Content
3.3. Selection and Assay Environments Affect Competitive Ability of O. tauri
4. Discussion
4.1. Susceptibility to OtV5 Did Not Change after Evolution
4.2. Resistance Type and Environment Affect Evolutionary Response of O. tauri to Environmental Change
4.3. Resistance Type Did Not Affect Competitive Ability Regardless of Environment
5. Conclusions
Supplementary Materials
Data
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Suttle, C.A. Marine viruses—Major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.W.; Suttle, C.A. Viruses and nutrient cycles in the sea. Bioscience 1999, 49, 781–788. [Google Scholar] [CrossRef]
- Koskella, B.; Brockhurst, M.A. Bacteria-phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiol. Rev. 2014, 38, 916–931. [Google Scholar] [CrossRef] [PubMed]
- Dennehy, J.J. What Can Phages Tell Us about Host-Pathogen Coevolution? Int. J. Evol. Biol. 2012, 2012, 396165. [Google Scholar] [CrossRef] [PubMed]
- Flint, S.J.; Enquist, L.W.; Krug, L.; Racaniella, V.; Skalka, A. Principles of Virology: Molecular Biology, Pathogenesis, Virus Ecology, 4th ed.; ASM Press: Washington, DC, USA, 2000. [Google Scholar]
- Doney, S.C.; Ruckelshaus, M.; Emmett Duffy, J.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate Change Impacts on Marine Ecosystems. Ann. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef] [PubMed]
- Nagasaki, K.; Yamaguchi, M. Effect of temperature on the algicidal activity and the stability of HaV (Heterosigma akashiwo virus). Aquat. Microb. Ecol. 1998, 15, 211–216. [Google Scholar] [CrossRef]
- Wells, L.E.; Deming, J.W. Effects of temperature, salinity and clay particles on inactivation and decay of cold-active marine Bacteriophage 9A. Aquat. Microb. Ecol. 2006, 45, 31–39. [Google Scholar] [CrossRef]
- Bellec, L.; Grimsley, N.; Derelle, E.; Moreau, H.; Desdevises, Y. Abundance, spatial distribution and genetic diversity of Ostreococcus tauri viruses in two different environments. Environ. Microbiol. Rep. 2010, 2, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Bratbak, G.; Egge, J.K.; Heldal, M. Viral mortality of the marine alga Emiliania huxleyi (Haptophyceae) and termination of algal blooms. Mar. Ecol. Prog. Ser. 1993, 93, 39–48. [Google Scholar] [CrossRef]
- Bratbak, G.; Jacobsen, A.; Heldal, M.; Nagasaki, K.; Thingstad, F. Virus production in Phaeocystis pouchetii and its relation to host cell growth and nutrition. Aquat. Microb. Ecol. 1998, 16, 1–9. [Google Scholar] [CrossRef]
- Wilson, W.H.; Carr, N.G.; Mann, N.H. The effect of phosphate status on the kinetics of cyanophage infection in the oceanic cyanobacterium Synechococcus sp. WH7803. J. Phycol. 1996, 32, 506–516. [Google Scholar] [CrossRef]
- Maat, D.; Crawfurd, K.; Timmermans, K.; Brussard, C. Elevated CO2 and phosphate limitation favor Micromonas pusilla through stimulated growth and reduced viral impact. Appl. Environ. Microbiol. 2014, 80, 3119–3127. [Google Scholar] [CrossRef] [PubMed]
- Jacquet, S.; Bratbak, G. Effects of ultraviolet radiation on marine virus-phytoplankton interactions. FEMS Microbiol. Ecol. 2003, 44, 279–289. [Google Scholar] [CrossRef]
- Jacquet, S.; Heldal, M.; Iglesias-Rodriguez, D.; Larsen, A.; Wilson, W.; Bratbak, G. Flow cytometric analysis of an Emiliana huxleyi bloom terminated by viral infection. Aquat. Microb. Ecol. 2002, 27, 111–124. [Google Scholar] [CrossRef]
- Thyrhaug, R.; Larsen, A.; Brussaard, C.P.D.; Mcfadden, P. Cell Cycle Dependent Virus Production in Marine Phytoplankton 1. Cell Cycle 2002, 343, 338–343. [Google Scholar]
- Larsen, J.B.; Larsen, A.; Thyrhaug, R.; Bratbak, G.; Sandaa, R.-A. Response of marine viral populations to a nutrient induced phytoplankton bloom at different pCO2 levels. Biogeosci. Discuss. 2007, 4, 3961–3985. [Google Scholar] [CrossRef]
- Chen, S.; Gao, K.; Beardall, J. Viral attack exacerbates the susceptibility of a bloom-forming alga to ocean acidification. Glob. Chang. Biol. 2014, 21, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Corinaldesi, C.; Dell’Anno, A.; Fuhrman, J.A.; Middelburg, J.J.; Noble, R.T.; Suttle, C.A. Marine viruses and global climate change. FEMS Microbiol. Rev. 2011, 35, 993–1034. [Google Scholar] [CrossRef] [PubMed]
- Bohannan, B.J.M.; Kerr, B.; Jessup, C.M.; Hughes, J.B.; Sandvik, G. Trade-offs and coexistence in microbial microcosms. Antonie van Leeuwenhoek 2002, 81, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Lenski, R.E. Experimental Studies of Pleiotropy and Epistasis in Escherichia coli. I. Variation in Competitive Fitness Among Mutants Resistant to Virus T4. Evolution 1988, 42, 425–432. [Google Scholar] [CrossRef]
- Lennon, J.T.; Khatana, S.A.M.; Marston, M.F.; Martiny, J.B.H. Is there a cost of virus resistance in marine cyanobacteria? ISME J. 2007, 1, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Frickel, J.; Sieber, M.; Becks, L. Eco-evolutionary dynamics in a coevolving host-virus system. Ecol. Lett. 2016, 19, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Seed, K.D.; Faruque, S.M.; Mekalanos, J.J.; Calderwood, S.B.; Qadri, F.; Camilli, A. Phase Variable O Antigen Biosynthetic Genes Control Expression of the Major Protective Antigen and Bacteriophage Receptor in Vibrio cholerae O1. PLoS Pathog. 2012, 8, e1002917. [Google Scholar] [CrossRef] [PubMed]
- León, M.; Bastías, R. Virulence reduction in bacteriophage resistant bacteria. Front. Microbiol. 2015, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Clerissi, C.; Desdevises, Y.; Grimsley, N. Prasinoviruses of the marine green alga Ostreococcus tauri are mainly species specific. J. Virol. 2012, 86, 4611–4619. [Google Scholar] [CrossRef] [PubMed]
- Marston, M.F.; Pierciey, F.J.; Shepard, A.; Gearin, G.; Qi, J.; Yandava, C.; Schuster, S.C.; Henn, M.R.; Martiny, J.B.H. Rapid diversification of coevolving marine Synechococcus and a virus. Proc. Natl. Acad. Sci. USA 2012, 109, 4544–4549. [Google Scholar] [CrossRef] [PubMed]
- Avrani, S.; Wurtzel, O.; Sharon, I.; Sorek, R.; Lindell, D. Genomic island variability facilitates Prochlorococcus-virus coexistence. Nature 2011, 474, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.R.; Agrawal, A.A.; Quick, R.T.; Dobias, D.T.; Schneider, D.; Lenski, R.E. Parallel changes in host resistance to viral infection during 45,000 generations of relaxed selection. Evolution 2010, 64, 3024–3034. [Google Scholar] [CrossRef] [PubMed]
- Avrani, S.; Lindell, D. Convergent evolution toward an improved growth rate and a reduced resistance range in Prochlorococcus strains resistant to phage. Proc. Natl. Acad. Sci. USA 2015, 112, E2191–E2200. [Google Scholar] [CrossRef] [PubMed]
- Avrani, S.; Schwartz, D.A.; Lindell, D. Virus-host swinging party in the oceans: Incorporating biological complexity into paradigms of antagonistic coexistence. Mob. Genet. Elem. 2012, 2, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Field, C.B. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Bellec, L.; Grimsley, N.; Moreau, H.; Desdevises, Y. Phylogenetic analysis of new Prasinoviruses (Phycodnaviridae) that infect the green unicellular algae Ostreococcus, Bathycoccus and Micromonas. Environ. Microbiol. Rep. 2009, 1, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Derelle, E.; Ferraz, C.; Escande, M.-L.; Eychenié, S.; Cooke, R.; Piganeau, G.; Desdevises, Y.; Bellec, L.; Moreau, H.; Grimsley, N. Life-cycle and genome of OtV5, a large DNA virus of the pelagic marine unicellular green alga Ostreococcus tauri. PLoS ONE 2008, 3, e2250. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Grimsley, N.; Escande, M.-L.; Subirana, L.; Derelle, E.; Moreau, H. Acquisition and maintenance of resistance to viruses in eukaryotic phytoplankton populations. Environ. Microbiol. 2011, 13, 1412–1420. [Google Scholar] [CrossRef] [PubMed]
- Yau, S.; Hemon, C.; Derelle, E.; Moreau, H.; Piganeau, G.; Grimsley, N. A Viral Immunity Chromosome in the Marine Picoeukaryote, Ostreococcus tauri. PLoS Pathog. 2016, 12, e1005965. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.D.; Selvin, R.C.; Claus, W.; Guillard, R.R.L. Media for the culture of oceanic ultraphytoplankton. J. Phycol. 1987, 23, 633–638. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models using lme4. J. Stat. Softw. 2014, 67, 51. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest: Tests in Linear Mixed Effects Models. Cran, R Package. 2015. Available online: http://CRAN.R-project.org/package=lmerTest (accessed on 15 January 2017).
- Schwarzländer, M.; Fricker, M.; Müller, C.; Marty, L.; Brach, T.; Novak, J.; Sweetlove, L.; Hell, R.; Meyer, A. Confocal imaging of glutathione redox potential in living plant cells. J. Microsc. 2008, 231, 299–316. [Google Scholar] [CrossRef] [PubMed]
- Corellou, F.; Schwartz, C.; Motta, J.-P.; Djouani-Tahri, E.B.; Sanchez, F.; Bouget, F.-Y. Clocks in the green lineage: Comparative functional analysis of the circadian architecture of the picoeukaryote Ostreococcus. Plant Cell 2009, 21, 3436–3449. [Google Scholar] [CrossRef] [PubMed]
- Van Ooijen, G.; Knox, K.; Kis, K.; Bouget, F.-Y.; Millar, A.J. Genomic Transformation of the Picoeukaryote Ostreococcus tauri. J. Vis. Exp. 2012, 65, e4074. [Google Scholar] [CrossRef] [PubMed]
- Heath, S.E.; Collins, S. Mode of resistance to viral lysis affects host growth across multiple environments in the marine picoeukaryote Ostreococcus tauri. Environ. Microbiol. 2016, 18, 4628–4639. [Google Scholar] [CrossRef] [PubMed]
- Schaum, E.; Collins, S. Plasticity predicts evolution in a marine alga. Proc. R. Soc. B 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Bell, G. Selection: The Mechanism of Evolution, 2nd ed.; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Lenski, R.E. Experimental Studies of Pleiotropy and Epistasis in Escherichia coli. II. Compensation for Maldaptive Effects Associated with Resistance to Virus T4. Evolution 1988, 42, 433–440. [Google Scholar] [CrossRef]
- Björkman, J.; Nagaev, I.; Berg, O.G.; Hughes, D.; Andersson, D.I. Effects of environment on compensatory mutations to ameliorate costs of antibiotic resistance. Science 2000, 287, 1479–1482. [Google Scholar] [PubMed]
- Bellec, L.; Clerissi, C.; Edern, R.; Foulon, E.; Simon, N.; Grimsley, N.; Desdevises, Y. Cophylogenetic interactions between marine viruses and eukaryotic picophytoplankton. BMC Evol. Biol. 2014, 14, 59. [Google Scholar] [CrossRef] [PubMed]
- Luria, S.; Delbrück, M. Mutations of Bacteria from Virus Sensitivity to Virus Resistance. Genetics 1943, 28, 491–511. [Google Scholar] [PubMed]
- Karafistan, A.; Martin, J.M.; Rixen, M.; Beckers, J.M. Space and time distributions of phosphate in the Mediterranean Sea. Deep. Sea Res. I 2002, 49, 67–82. [Google Scholar] [CrossRef]
- Jessup, C.M.; Bohannan, B.J.M. The shape of an ecological trade-off varies with environment. Ecol. Lett. 2008, 11, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Bohannan, B.J.M.; Travisano, M.; Lenski, R.E. Epistatic Interactions Can Lower the Cost of Resistance to Multiple Consumers. Evolution 1999, 53, 292–295. [Google Scholar] [CrossRef]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Raven, J.A. Phytoplankton in a changing world: Cell size and elemental stoichiometry. J. Plankton Res. 2010, 32, 119–137. [Google Scholar] [CrossRef]
- Peter, K.H.; Sommer, U. Interactive effect of warming, nitrogen and phosphorus limitation on phytoplankton cell size. Ecol. Evol. 2015, 5, 1011–1024. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, D.; Ciotti, B.J.; Montagnes, D.J.S. Protists decrease in size linearly with temperature: Ca. 2.5% °C−1. Proc. Biol. Sci. 2003, 270, 2605–2611. [Google Scholar] [CrossRef] [PubMed]
- Morán, X.A.G.; López-Urrutia, Á.; Calvo-Díaz, A.; Li, W.K.W. Increasing importance of small phytoplankton in a warmer ocean. Glob. Chang. Biol. 2010, 16, 1137–1144. [Google Scholar] [CrossRef]
- Geider, R.; Platt, T.; Raven, J. Size dependence of growth and photosynthesis in diatoms: A synthesis. Mar. Ecol. Ser. 1986, 30, 93–104. [Google Scholar] [CrossRef]
- Šupraha, L.; Gerecht, A.C.; Probert, I.; Henderiks, J. Eco-physiological adaptation shapes the response of calcifying algae to nutrient limitation. Sci. Rep. 2015, 5, 16499. [Google Scholar] [CrossRef] [PubMed]
- Ryther, J.; Menzel, D. Light adaptation by marine phytoplankton. Limnol. Oceanogr. 1959, 4, 492–497. [Google Scholar] [CrossRef]
- Wozniak, B.; Hapter, R.; Dera, J. Light curves of marine plankton photosynthesis in the Baltic. Oceanologia 1989, 29, 61–78. [Google Scholar]
- Renk, H.; Ochocki, S. Photosynthetic rate and light curves of phytoplankton in the southern Baltic. Oceanologia 1998, 40, 331–344. [Google Scholar]
- Riemann, B.; Simonsen, P.; Stensgaard, L. The carbon and chlorophyll content of phytoplankton from various nutrient regimes. J. Plankton Res. 1989, 11, 1037–1045. [Google Scholar] [CrossRef]
- McLachlan, J. The effect of salinity on growth and chlorophyll content in representative classes of unicellular marine algae. Can. J. Microbiol. 1961, 7, 399–406. [Google Scholar] [CrossRef]
- Sigaud, T.C.S.; Aidar, E. Salinity and temperature effects on the growth and chlorophyll-a content of some planktonic aigae. Bol. Inst. Oceanogr. 1993, 41, 95–103. [Google Scholar] [CrossRef]
- Brockhurst, M.A.; Rainey, P.B.; Buckling, A. The effect of spatial heterogeneity and parasites on the evolution of host diversity. Proc. Biol. Sci. 2004, 271, 107–111. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Selection Environment | Control | Treatment |
|---|---|---|
| Light (μmol m−2 s−1) | 85 | 60 |
| Phosphate (μM) | 10 | 5 |
| Salinity (ppt) | 30 | 25 |
| Temperature (°C) | 18 | 20 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heath, S.E.; Knox, K.; Vale, P.F.; Collins, S. Virus Resistance Is Not Costly in a Marine Alga Evolving under Multiple Environmental Stressors. Viruses 2017, 9, 39. https://doi.org/10.3390/v9030039
Heath SE, Knox K, Vale PF, Collins S. Virus Resistance Is Not Costly in a Marine Alga Evolving under Multiple Environmental Stressors. Viruses. 2017; 9(3):39. https://doi.org/10.3390/v9030039
Chicago/Turabian StyleHeath, Sarah E., Kirsten Knox, Pedro F. Vale, and Sinead Collins. 2017. "Virus Resistance Is Not Costly in a Marine Alga Evolving under Multiple Environmental Stressors" Viruses 9, no. 3: 39. https://doi.org/10.3390/v9030039
APA StyleHeath, S. E., Knox, K., Vale, P. F., & Collins, S. (2017). Virus Resistance Is Not Costly in a Marine Alga Evolving under Multiple Environmental Stressors. Viruses, 9(3), 39. https://doi.org/10.3390/v9030039