1. Introduction
The rise in anthropogenic CO
2 in the atmosphere and subsequent dissolution in the oceans has changed the carbonate: bicarbonate: dissolved CO
2 equilibrium, lowering seawater pH—a trend that is predicted to continue [
1]. Change of pH is of particular significance for marine organisms that have calcium carbonate structures, such as corals and coccolithophores, because less alkaline conditions and pH-dependent shifts in equilibrium of the carbonate system will lead to higher dissolution rates of carbonate. Coccolithophores are ubiquitous and have global significance in regulating the carbon cycle in the oceans [
2]. They form massive blooms, whose wide distribution and abundance is readily detected in satellite imagery. Given this wide distribution, it is important to determine if the lower pH of a future ocean will affect the success of coccolithophores and if there will be an impact on marine food webs and biogeochemical cycles.
The effect of changing pH on the important coccolithophore,
Emiliania huxleyi, has been the focus of much research in recent years. However, results have been variable and consensus has been difficult to reach. In laboratory experiments, both negative and positive effects of increasing
pCO
2 have been described (see, for example, [
3,
4,
5]). Another important approach has been to use large volume enclosures—mesocosms—to investigate a range of conditions that might apply to the future ocean. Unlike laboratory-based experiments, which usually focus on a single organism in the experimental design, mesocosms include all components of the pelagic system from viruses to zooplankton. By maintaining the possibility of complex interactions between different components of the food web, it has been assumed that mesocosms should offer advantages over single-organism culture experiments. However, results have also been rather variable. Early experiments suggested negative effects of higher
pCO
2 on production and calcification in
E. huxleyi [
6], but other studies have indicated that the effect of increased
pCO
2 is minimal for other coccolithophore species [
7]. Time series analysis of natural populations has been another approach and a recent analysis of coccolithophore abundance in the North Sea concluded that increasing pCO
2 on decadal scales has resulted in larger coccolithophore populations [
8]. The contradictory results make it difficult to robustly predict how natural populations will respond to pH change in a future ocean.
We suggest that real understanding of the effect of pH change/higher
pCO
2 requires more detailed information than has been obtained to date, particularly in relation to phytoplankton genetic variability and virus infection. In this study, the response of a population of
E. huxleyi to increased
pCO
2 at the early stages of a phytoplankton bloom in a mesocosm experiment has been investigated. In addition, changes in diversity of the viruses that infect
E. huxleyi (EhVs) were followed during the experiment with diversity distinguished on the basis of a major capsid protein (
mcp) gene as a molecular marker. Virus diversity is known to be high [
9,
10], and viruses are important components of the pelagic system that require attention in both laboratory and mesocosm experiments. All
E. huxleyi-infecting viruses that have been characterised to date are dsDNA viruses, classified in the family
Phycodnaviridae. They are significant mortality agents of
E. huxleyi, implicated in the termination of large-scale blooms. Viruses have a proven role in structuring and maintaining host population diversity [
11,
12,
13,
14] and virus infection can have significance for the cycling of carbon and trace elements. The ‘viral shunt’ releases nutrients as well as dissolved and particulate organic matter from lysed organisms into the organic carbon pool [
15,
16]. This material, and the rate of supply by viral lysis, of substrates for heterotrophic microbial communities, has implications for species succession, biogeochemical cycles and feedback mechanisms.
Given that diversity of both
E. huxleyi and EhV assemblages can be variable, and that different
E. huxleyi and EhV assemblages may come to eventually dominate natural communities, it is important to know the impact of elevated
pCO
2.
E. huxleyi blooms are typically dominated by certain alleles/genotypes, and by asexual reproduction, with rarer alleles/genotypes tending to fluctuate [
17]. As such, the impact of elevated CO
2 on the composition of
E. huxleyi populations can easily be monitored by studying these entities. Virus infection may be an explanation for some of the variability reported from different mesocosm experiments that were designed to investigate potential effects of higher
pCO
2. In this study, the aim was to understand how
pCO
2 change might influence
E. huxleyi and EhV population structure and the diversity of host and virus. We suggest that viral infection can result in variability between replicate enclosures.
4. Discussion
A number of mesocosm experiments have been performed to investigate the potential effects of increased pCO2 and reduced pH on complex pelagic assemblages—from bacteria to zooplankton. Mesocosm enclosures have the advantage of capturing more of the intrinsic complexity of a pelagic assemblage than is possible in a laboratory experiment because of the very large volumes of water (several thousand litres) that are involved. They also, by the nature of enclosure, eliminate problems of dispersal that make the study of variability in natural environments so complex. They are fundamentally attractive to experimentalists because they offer a means to manipulate large water volumes, with their associated planktonic assemblages, in order to test the effects of environmental problems such as eutrophication or ocean acidification.
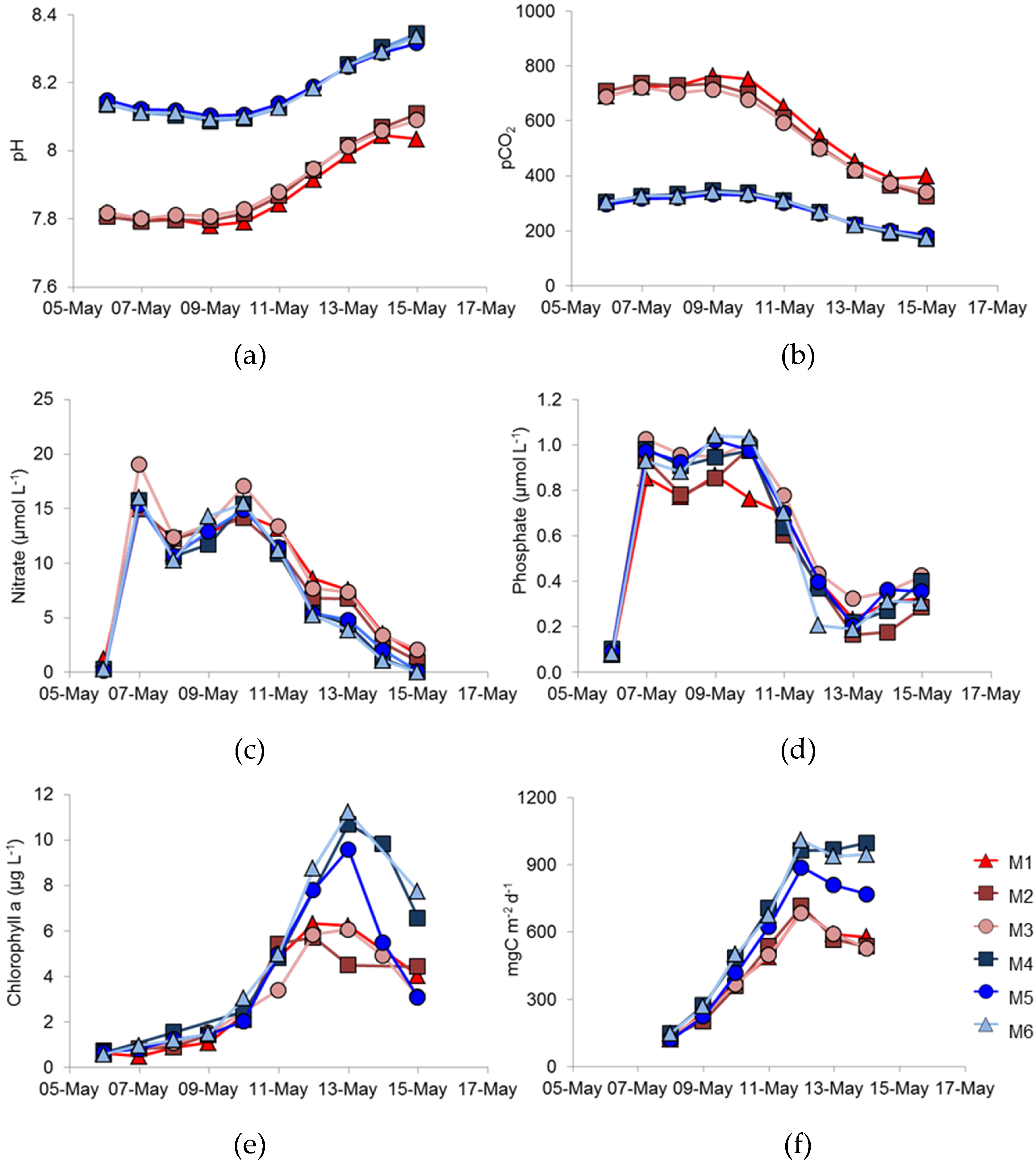
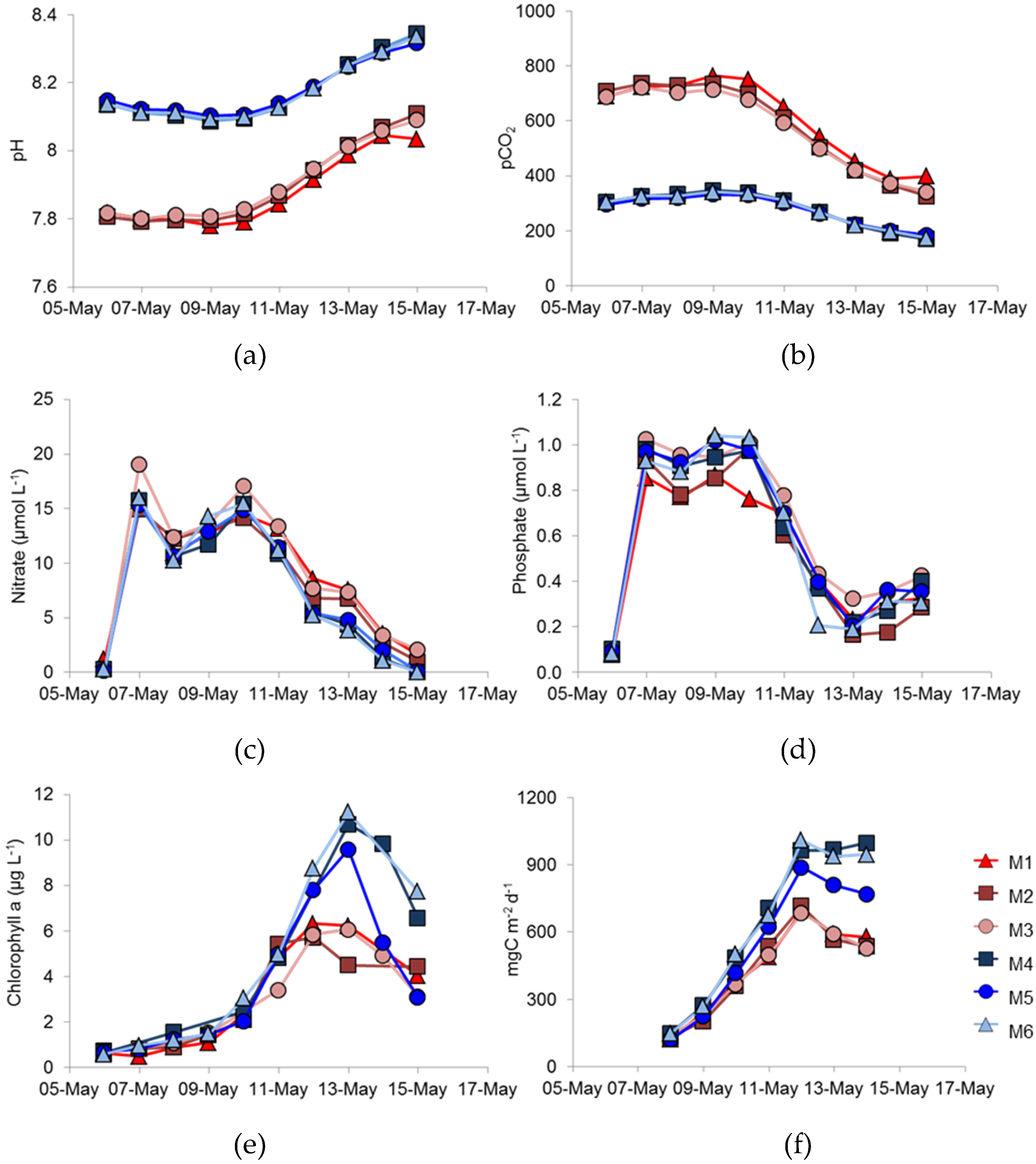
In this study, we aimed to investigate how phytoplankton might respond in a future high-
pCO
2 ocean by comparing natural phytoplankton assemblage development in enclosures at ambient
pCO
2, (initial condition ca. 300 µatm.) and enriched
pCO
2 conditions (initially ca. 700 µatm.)—
Figure 1a,b; see also Hopkins et al. [
18]. During the course of the experiment, utilisation of CO
2 by phytoplankton reduced
pCO
2 and the
pCO
2/pH values were continually changing. An obvious difference between the treatments was that less phytoplankton biomass, as indicted by chlorophyll
a concentration, developed in the high- compared to the ambient-
pCO
2 conditions (
Figure 1e). Not only did less biomass develop, but primary production (
Figure 1f) was also lower under high-, rather than under ambient-
pCO
2, suggesting that increased
pCO
2 might have a deleterious effect on the total phytoplankton biomass. In contrast, other experiments have suggested that dissolved inorganic carbon uptake would be enhanced under elevated
pCO
2 conditions [
28]. Different phytoplankton types are likely to respond differently to
pCO
2 and Riebesell et al. suggested that diatoms showed enhanced uptake, whereas coccolithophores did not [
28]. There was no suggestion in that study, though, that coccolithophore growth might be reduced under elevated
pCO
2 conditions.
The dominant species of coccolithophore within the mesocosms was
E. huxleyi, with coccolithophores being reported as contributing 6% and 12% to the total flagellate biomass for M1 and M6, respectively [
18]. Manipulating the development of
E. huxleyi blooms within mesocosm enclosures is well established at the mesocosm facility at Raunefjorden, and is well documented. Addition of nitrate and phosphate, with the omission of silicate, usually results in a bloom of
E. huxleyi at this site, particularly during May/June [
11,
12,
14] and an increase in
E. huxleyi abundance happened in the present study. Previous mesocosm experiments of this nature have described a dominance of coccolithophores; however, differences in methodology most likely resulted in the dominance of large picoeukaryotes [
18]. In the present study, a single nutrient enrichment was undertaken at the beginning of the study, whereas daily enrichments are often used e.g., Jacquet et al. [
29]. Nevertheless, the maximum number of
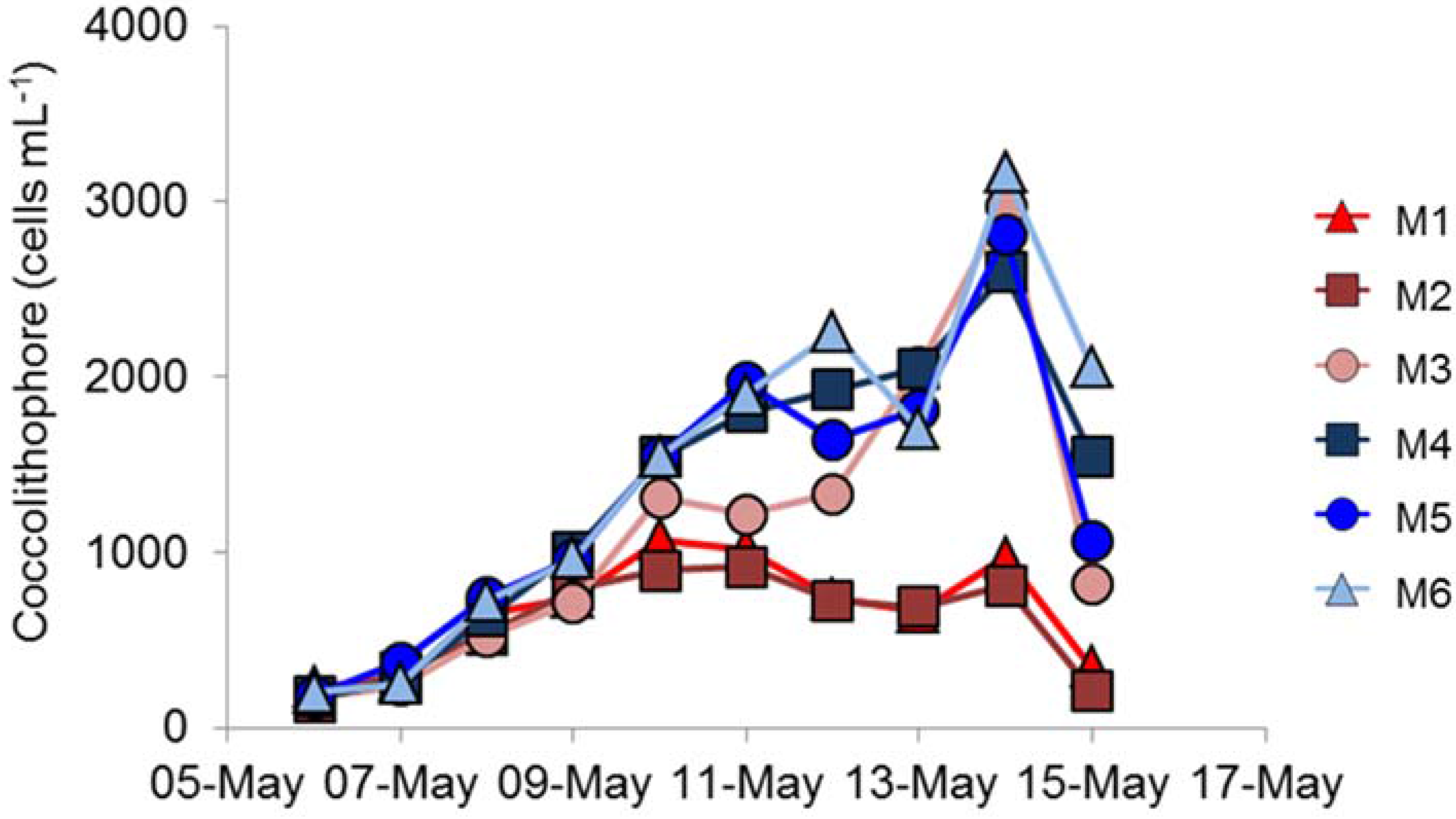
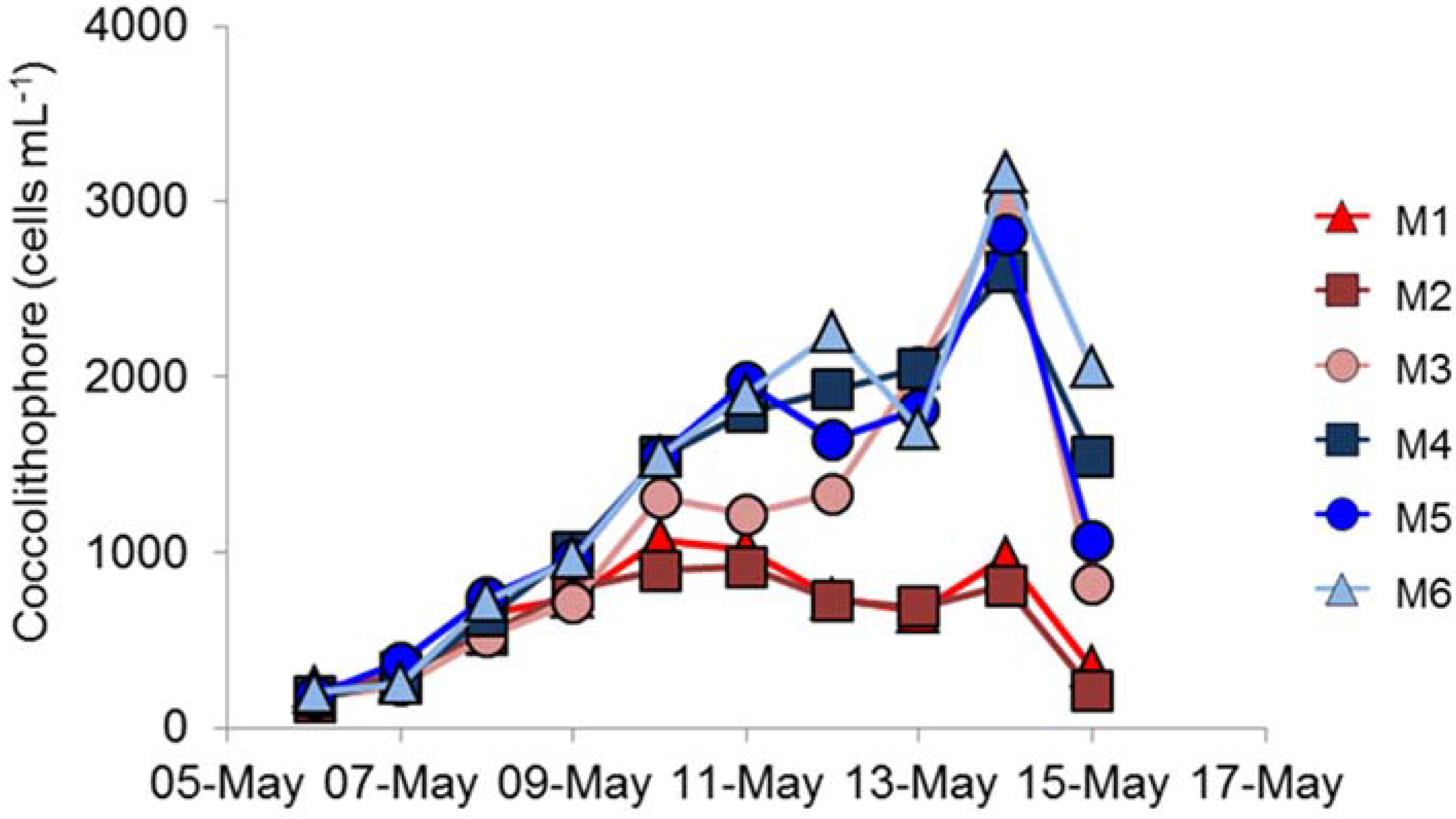
E. huxleyi cells that developed was significantly lower in the high- compared to ambient-
pCO
2 conditions (
Figure 2), suggesting that
E. huxleyi might be particularly susceptible to variations in
pCO
2.
Replication between enclosures was rather variable. The peak chlorophyll concentrations were very similar in the three ambient
pCO
2 replicates (
Figure 1e), but the bloom decayed more rapidly in M5 than in M4 and M6; there were similar differences in primary production (
Figure 1f). However, the largest difference between replicates was in coccolithophore numbers (
Figure 2), which showed significant differences between the three high-
pCO
2 treatment mesocosms with such differences between replicates in other key phytoplankton groups not reported by Hopkins et al. [
18]. Enclosure M3 had coccolithophore cell numbers at the peak of the bloom that were very similar to the three ambient
pCO
2 mesocosms, unlike the lower numbers in M1 and M2. Although cell numbers were different in M3, no major differences in
E. huxleyi genotype or phenotype were detected between treatments or over the course of the experiment, suggesting a stable community within all enclosures over the duration of the experiment. Stable
E. huxleyi populations have been found in previous mesocosm experiments [
12,
14] and, indeed, in naturally occurring
E. huxleyi blooms [
9,
17]. In this study, we have no evidence to support the hypothesis that higher
pCO
2 conditions might benefit certain
E. huxleyi genotypes; we could detect no restructuring of the
E. huxleyi population in the different
pCO
2 treatments.
Contradictory results are constant features of experiments to investigate the effect of
pCO
2/pH change on coccolithophores. In laboratory culture experiments, Riebesell et al. [
30], Zondervan et al. [
31] and Richier et al. [
32] all reported increased production by
E. huxleyi under elevated
pCO
2 conditions, but Sciandra et al. [
33] and Langer et al. [
4] found decreased production. In two different CO
2-manipulated mesocosm experiments in the Raunefjorden, Engel et al. [
6] and Paulino et al. [
34] found little difference in
E. huxleyi cell concentrations over the course of their experiments. However, Engel et al. [
6] calculated that the net specific growth rate of
E. huxleyi was reduced at 710 µatm compared with 410 μatm. In a long-term batch culture experiment conducted over one year, Lohbeck et al. [
5] found that
E. huxleyi cultures maintained at ambient
pCO
2 (400 μatm) went through 530 generations over the one year experimental period, but the same strain cultured at 1100 μatm achieved only 500 generations, and, at 2200 μatm, growth was even lower, with only 430 generations.
E. huxleyi would appear to be more sensitive to
pCO
2 change than other phytoplankton.
In the context of the present study, the reduced primary production in the high-
pCO
2 treatment enclosures is consistent with the finding of Lohbeck et al. [
5] that higher
pCO
2/lower pH reduces primary production of an
E. huxleyi dominated phytoplankton assemblage, although, in this study, other phytoplankton groups, e.g., picoeukaryotes, cryptophytes and cyanobacteria, will have also contributed to this. However, it is not consistent with the suggestion of Rivero-Calle et al. [
8] that increasing
pCO
2 is one of the factors most responsible for the decadal increase in abundance of coccolithophores in the North Atlantic. The relationship between
pCO
2/pH change and success or, otherwise of cocccolithophores, remains confusing and requires clearer examination of mechanisms that might lead to phytoplankton changes in the future ocean.
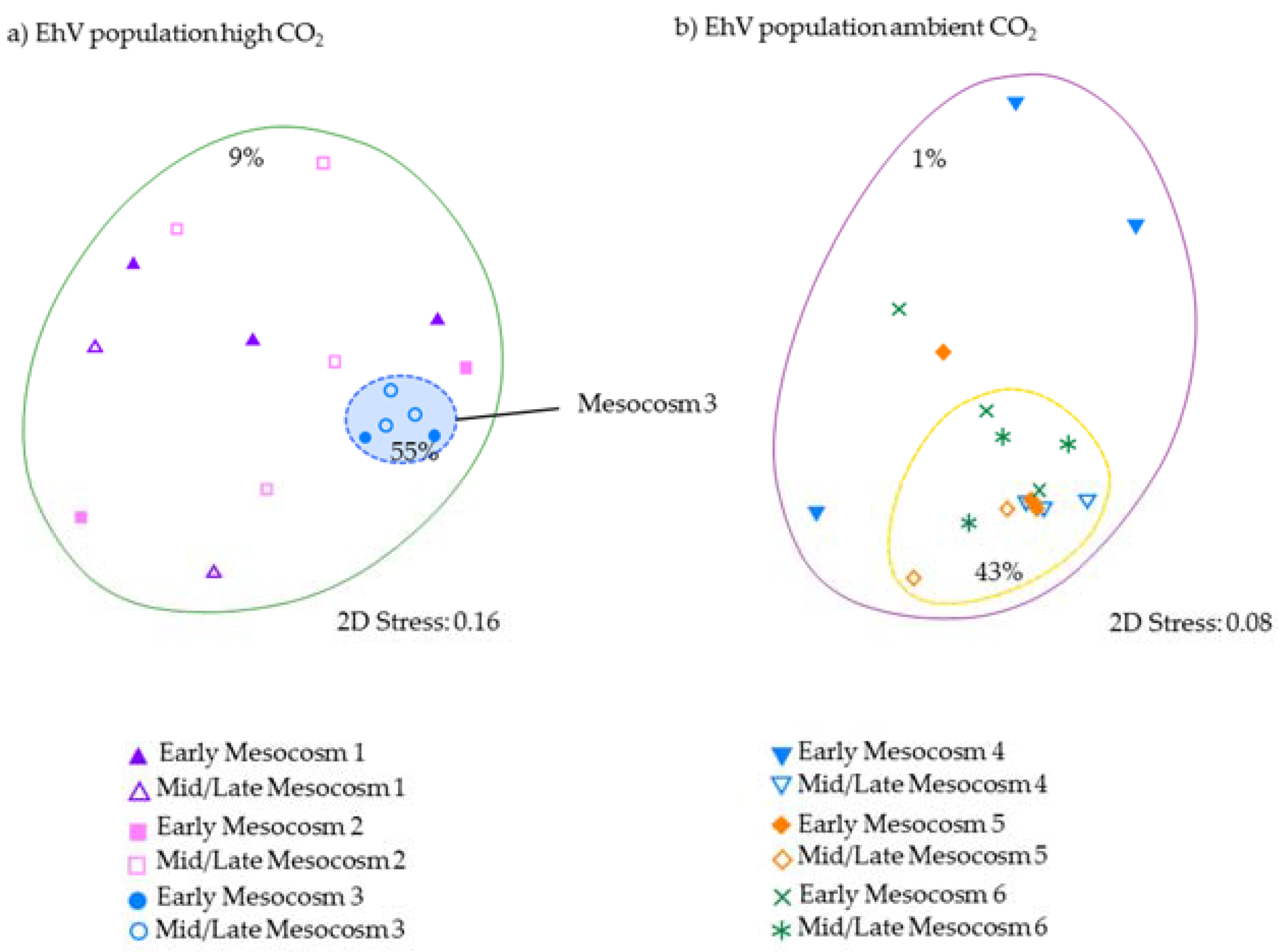
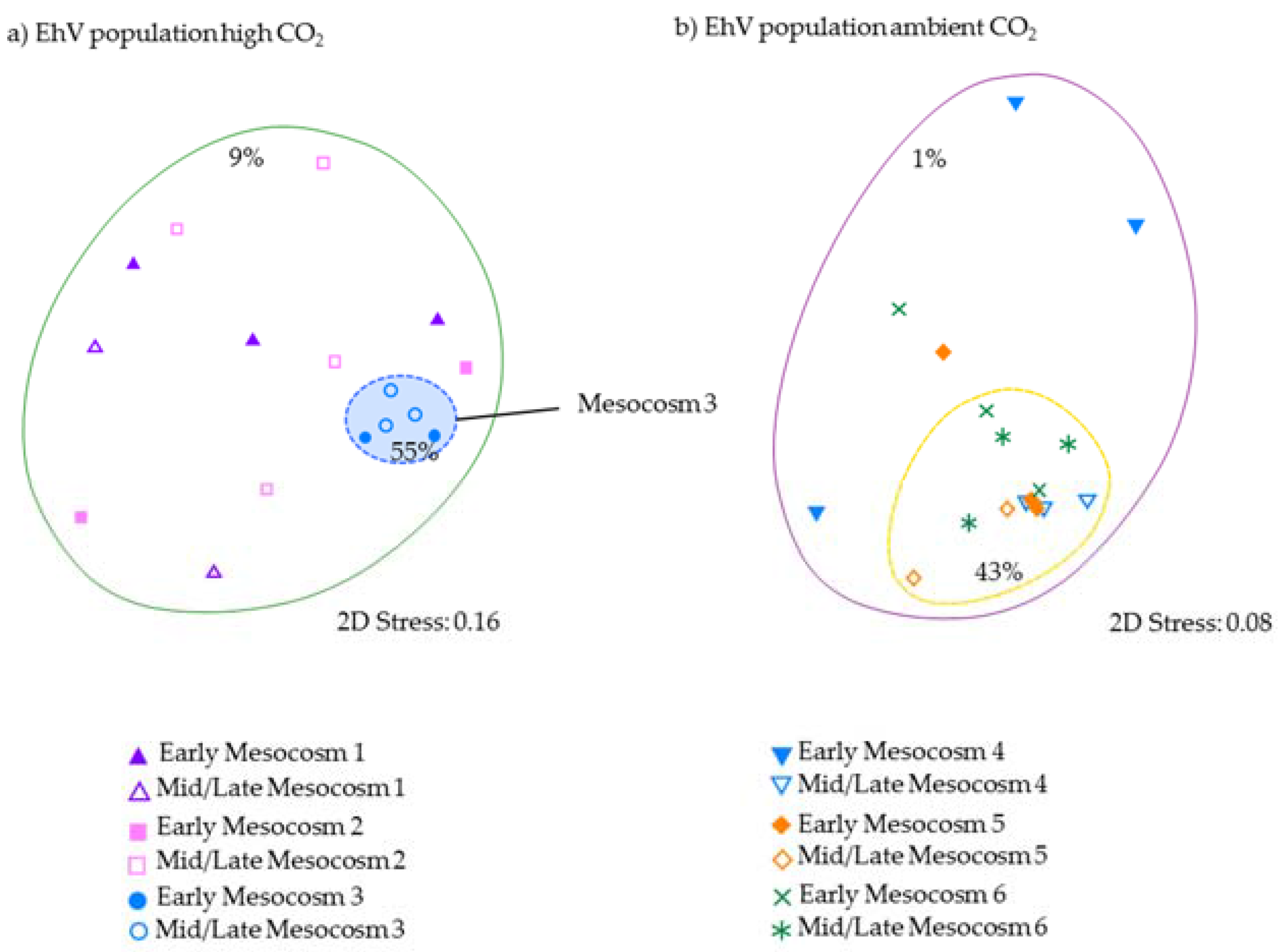
Given that the coccolithophores numbers in enclosure M3 were very different from the other two high pCO
2 treatments, and yet the
E. huxleyi genetic diversity was very similar in all three enclosures, could viral infection by EhV be a contributing factor to explain the observed variations within and between treatments? In the three ambient-
pCO
2 enclosures, the EhV population followed a pattern that has been seen in other mesocosm experiments—high variability during the early stages of phytoplankton bloom development, with a smaller number of genotypes coming to dominate as the
E. huxleyi numbers increase [
12,
14]. Although phytoplankton bloom development in the current study was short (<10 days), there was sufficient time for the virus populations to change because EhV populations are inherently dynamic [
9] and known to change on very short time scales [
14]. Daily changes in EhV composition can be expected since Sorensen et al. [
14] showed that EhVs can appear/disappear from the water column in a matter of hours.
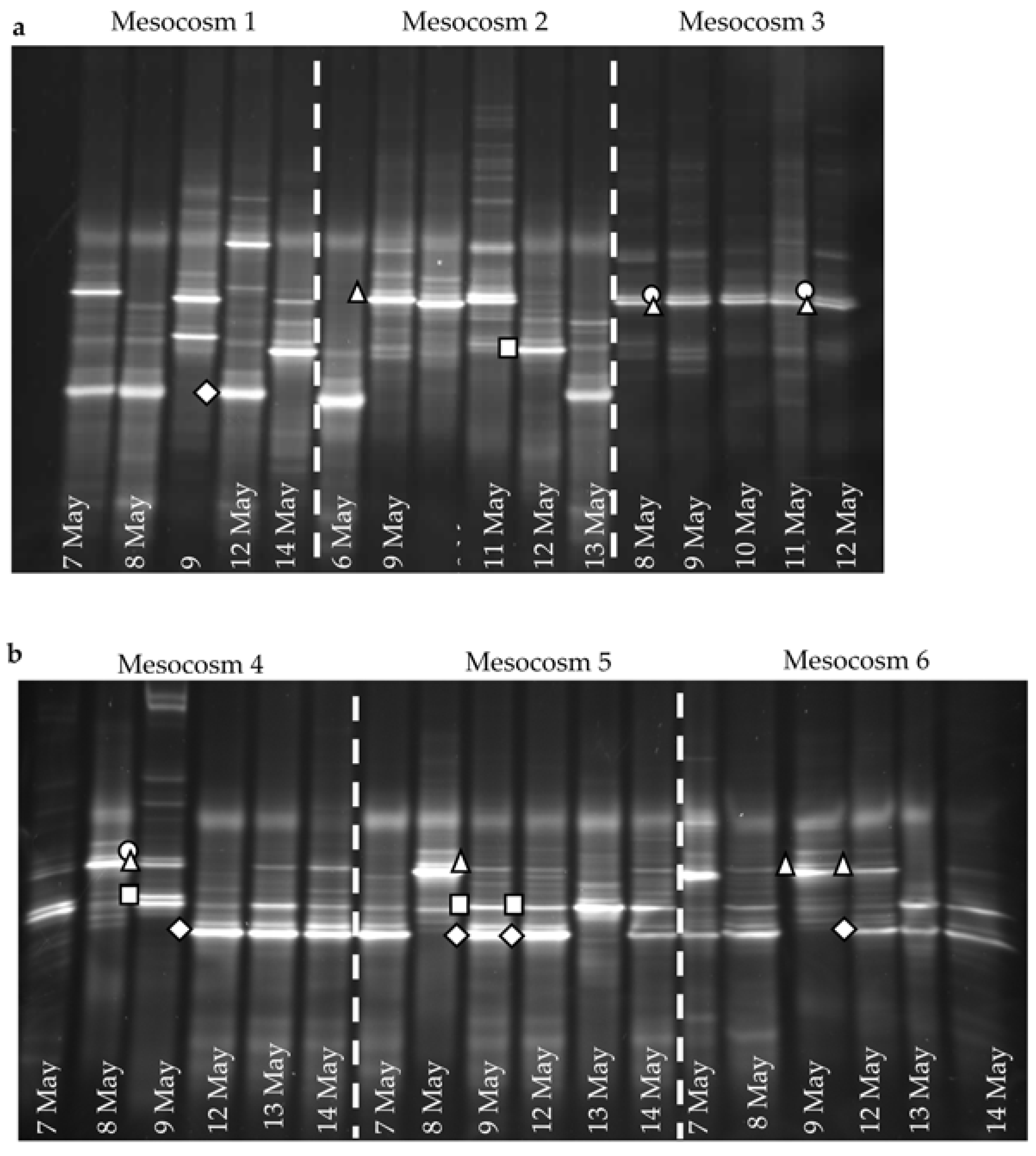
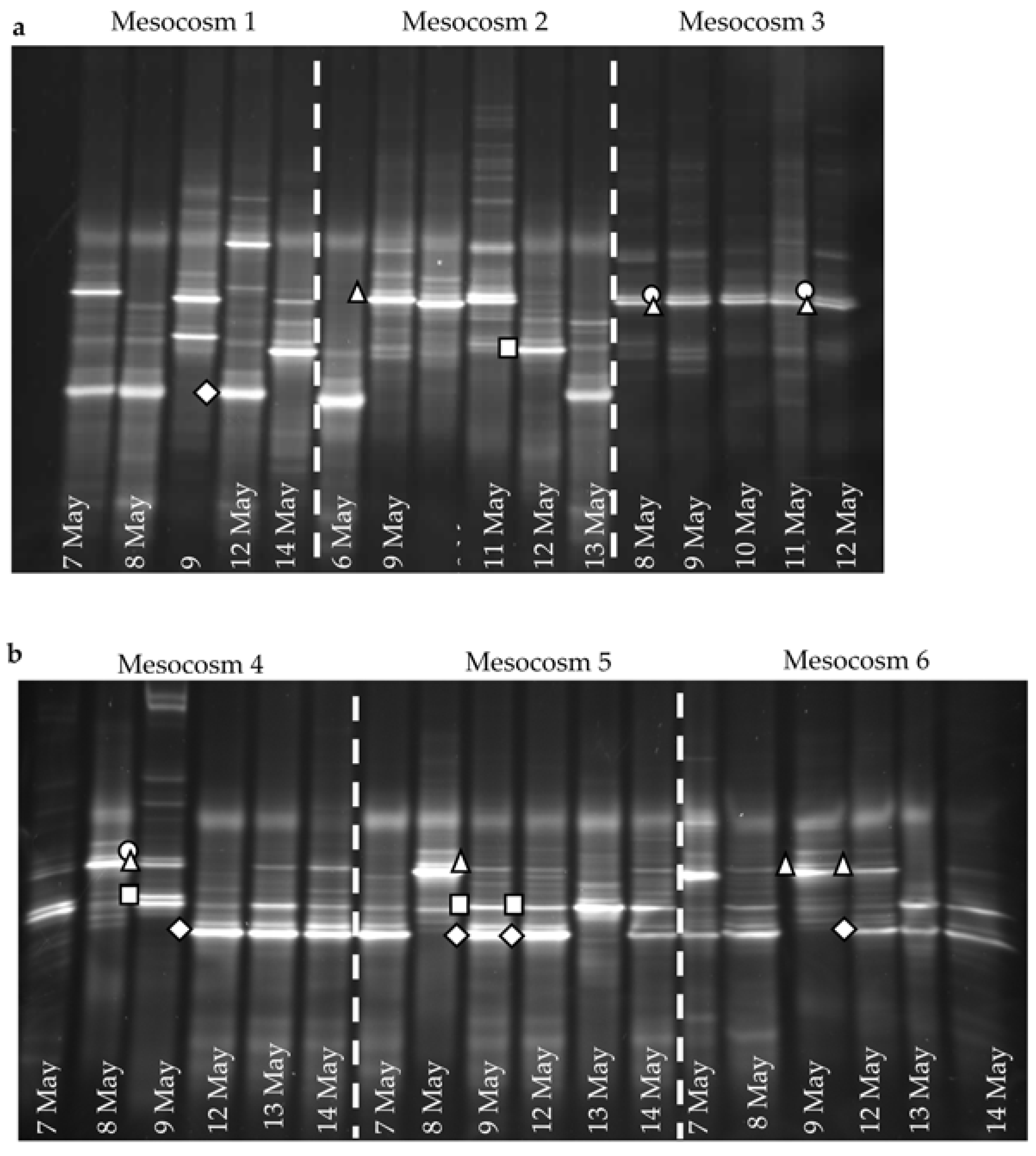
In the high-
pCO
2 treatment enclosures M1 and M2, where
E. huxleyi population densities did not exceed 1.1 × 10
3 mL
−1, the EhV population did not stabilise (
Figure 4 and
Figure 5) and EhV diversity was typical of early or non-bloom conditions [
10]; that is, it was a highly dynamic and diverse EhV population. EhVs are known to have different host ranges [
35] as well as different characteristics of infection, such as burst size and latent period. Therefore, any changes in environmental conditions that directly affect these traits could ultimately select for different genotypes, hence restructuring the EhV population.
In contrast, the third high-
pCO
2 treatment enclosure M3, where coccolithophore cell densities reached similar values to ambient enclosures, and the EhV population structure was very different. Two EhVs dominated over the course of the experiment (
Figure 4) and the population was much less changeable compared to the other high-
pCO
2 enclosures and, indeed, to the three ambient enclosures. The EhV assemblage in M3, right from the early stages of the experiment, reflected what would be expected in the later stages of a bloom. Even in the early stages of the bloom, the EhV assemblage was stable and clustered closely in the MDS plot (
Figure 5) with samples taken later in the bloom.
Given that pH and
pCO
2 were so similar throughout the experiment in the three high-
pCO
2 enclosures, why did enclosure M3 have lower coccolithophore numbers and a lower and stable EhV diversity? Other studies have shown that, under non-bloom conditions, many different EhV genotypes are present and abundance fluctuates on short time scales [
14]. In addition, during the initial phase of an
E. huxleyi bloom, EhV populations remain diverse and are often highly dynamic. Sorensen et al. [
14] showed that, as a bloom developed in a mesocosm experiment and viruses numbers proliferated, one viral genotype dominated, and suggested that this dominant virus caused the termination of the bloom. However, the dominant EhV is not always the same, even when host genotypes do not vary. Martínez-Martínez et al. [
12] and Sorensen et al. [
14] found that, although
E. huxleyi populations were dominated by the same genotypes in different years at Raunefjorden (2000, 2003 and 2008), viruses changed and the dominant EhV in 2008 was different to the EhVs that dominated during the 2000 and 2003 mesocosm experiments. The reason is not known, but these authors suggest that a slight change in environmental conditions might have favoured dominance by a different virus genotype. Whilst the number of mesocosms sampled might be perceived as limited, the fact that Martinéz-Martinéz [
35] described how the same DGGE profile was generated from four replicate mesocosms; in his studies, we can assume that the changes that we are observing are genuine.
Other mesocosm experiments have studied how altered
pCO
2 influences natural virus communities. Larsen et al. [
36], using flow cytometry analysis and pulsed field gel electrophoresis (PFGE), found slightly more (but statistically insignificant) EhVs under present-day
pCO
2 mesocosms than in high-
pCO
2 treatments; this was not a consequence of lower
E. huxleyi cell densities in the high-
pCO
2 treatments. The authors speculated that elevated-
pCO
2 may affect host–virus interactions or influence viral replication, since a 26 kb genome virus was only detected in ambient conditions, and was absent from high-
pCO
2 treatments, and a 105 kb genome virus was only detected in the highest
pCO
2 treatment of 1050 ppm. Unfortunately, the taxonomic affiliation of the viruses was not verified, which limits comparison with the present study.
Some laboratory experiments have investigated the effect of higher
pCO
2 on marine phytoplankton viruses. Carreira et al. [
37] studied interaction between
E. huxleyi and the virus EhV-99B1.
E. huxleyi growth rate was not affected by the different
pCO
2 treatments, but the burst size of EhV-99B1 was lower in present-day, compared with higher and lower (pre-industrial)
pCO
2. In addition, release of EhVs was delayed in high-
pCO
2 treatments. Other virus groups that have also been tested for sensitivity to elevated
pCO
2. Chen et al. [
38] found lower burst size of the
Phaeocystis globosa virus (PgV) at high-
pCO
2, and Traving et al. [
39] found that the cyanophage S-PM2, which infects
Synechococcus sp, had reduced burst size at lower pH. However, the extracellular phase, quantified as infectivity loss rates/decay, did not change. These experiments illustrate that
pCO
2 can influence virus–host interactions, albeit to a relatively minor extent. However, extrapolation from these laboratory-scale experiments to natural virus communities would involve considerable uncertainty.
Another study considered a much longer time scale. Coolen [
40] investigated
E. huxleyi and EhV diversity in Black Sea sediments over a 7000 year period, showing that EhV diversity was highest during periods of change in hydrological and nutrient regimes. Shifts in EhV genotypic diversity typically coincided with Holocene environmental change with some viruses having limited persistence, yet others were found to persist for over a century. This study alluded to the impact that a change in CO
2/pH could have on future EhV populations.
Might the different EhV populations that dominated in each enclosure be an explanation for the differences observed? It is generally accepted that virus infection is a major reason why
E. huxleyi cells stop growing and blooms are terminated [
14]. Certainly, nutrients were still available, albeit at low concentrations when biomass peaked in each enclosure (
Figure 1c,d). If viral mortality was the major limit on bloom development, then enclosures M1 & M2 must have been infected with more aggressive EhVs than in the other treatments. The effect of
pCO
2 treatment, per se, on
E. huxleyi cells cannot be responsible for the reduced growth in the two high-
pCO
2 treatments because an identical pH/pCO
2 treatment in M3 did not reduce the size of the bloom. Therefore,
pCO
2 change must have resulted in different viral diversity, if infection is indeed the main cause of lower cell numbers and smaller
E. huxleyi bloom.
How might reduced EhV diversity in M3 have resulted in higher coccolithophore numbers developing than in the two other high
pCO
2 treatments where growth was curtailed? One explanation would be that the EhVs that lead to rapid termination of
E. huxleyi growth [
14] were not present in sufficient numbers in M3 to suppress growth of the
E. huxleyi population in this enclosure. Genetic diversity in natural populations of EhVs, especially in pre-bloom conditions [
10], coupled with the rapid rate at which individual EhVs can come to dominate [
9,
14] means that the matrix of EhVs that could be selected for is large. It is not clear why only two EhVs were dominant in enclosure M3, but it is probably significant that viral infection in this enclosure did not suppress growth of
E. huxleyi compared to M1 and M2.
In this study, it is difficult to distinguish whether
pCO
2 change is affecting the viruses specifically, or their hosts independently, and/or the interactions between them. Both the external virus population (the virus particles present in the water used to fill the mesocosms) and internal virus population (present in infected
E. huxleyi cells) are important for the ultimate progression of the
E. huxleyi bloom and EhV population. By comparing data from Schroeder et al. [
13] and Martínez Martínez [
12] it can be seen that the diversity of EhVs amplified from water samples within a mesocosm bloom can be very different from that amplified from
E. huxleyi cells, particularly at the early stage of the bloom—that is, external and internal EhV assemblages can have very different composition. Thus, studies that aim to explain the effect of elevated
pCO
2 must incorporate into their experimental design ways to distinguish between a direct effect on external virus particles, or on
E. huxleyi cells, or on EhVs that have already infected
E. huxleyi cells. Alternatively, other unknown factors that are not related to
pCO
2 cannot be dismissed and might be the cause of the different coccolithophore response in enclosure M3.
Our study demonstrates the need for further investigations on the effects of elevated
pCO
2 on the
E. huxleyi EhV system since there are ecological impacts of virus competition on biogeochemical cycles. Nissimov et al. [
41] investigated competition between the two EhVs, finding that EhV-207 had a competitive advantage over EhV-86. It would thus be of value to determine how external factors, such as elevated CO
2, would affect relative competitive ability of EhVs; would EhV-207 still outcompete EhV-86?
In this study, we have shown that elevated pCO2 can affect the structure of EhV assemblages. The data do not allow us to distinguish if this is a direct impact of pCO2 on the viruses themselves, but it is clear that caution is required in interpreting results from manipulation experiments and that deep analysis is required to truly understand how complex assemblages respond. For example, analysis of only the chlorophyll concentration or primary production data would not have revealed that there were differences in the triplicate high-pCO2 treatments: cell counts and microscopy would not have revealed the difference in E. huxleyi diversity in the replicate enclosures, and only analysis of virus genotypes could have revealed how different the viruses were in apparently identical replicate enclosures. We suggest that viruses cannot be ignored in any study of the potential effects of ocean acidification on phytoplankton productivity in the future high-CO2 ocean.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}