Virus Infection and Death Receptor-Mediated Apoptosis

Abstract
1. Introduction of Virus-Mediated Apoptosis
2. Intrinsic and Extrinsic Apoptosis Pathways
3. Viral Induction and Viral Activators of Death Receptor-Mediated Apoptosis
3.1. Human Immunodeficiency Virus (HIV)
3.2. Hepatitis Viruses
3.3. Herpesviruses
3.4. Other RNA Viruses
3.5. Other DNA Viruses and Retroviruses
4. Viral Inhibitors of Death Receptor-Mediated Apoptosis
4.1. Herpesviruses
4.2. Hepatitis Viruses
4.3. Adenoviruses
4.4. Human Papillomaviruses
4.5. Other Viruses
5. Consequence of Death Receptor-Mediated Apoptosis during Viral Infection
6. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Danthi, P. Viruses and the diversity of cell death. Annu. Rev. Virol. 2016, 3, 533–553. [Google Scholar] [CrossRef] [PubMed]
- Teodoro, J.G.; Branton, P.E. Regulation of apoptosis by viral gene products. J. Virol. 1997, 71, 1739–1746. [Google Scholar] [PubMed]
- Galluzzi, L.; Brenner, C.; Morselli, E.; Touat, Z.; Kroemer, G. Viral control of mitochondrial apoptosis. PLoS Pathog. 2008, 4, e1000018. [Google Scholar] [CrossRef] [PubMed]
- Everett, H.; McFadden, G. Apoptosis: An innate immune response to virus infection. Trends Microbiol. 1999, 7, 160–165. [Google Scholar] [CrossRef]
- Shen, Y.; Shenk, T.E. Viruses and apoptosis. Curr. Opin. Genet. Dev. 1995, 5, 105–111. [Google Scholar] [CrossRef]
- Roulston, A.; Marcellus, R.C.; Branton, P.E. Viruses and apoptosis. Annu. Rev. Microbiol. 1999, 53, 577–628. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vitale, I.; Abrams, J.M.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; Dawson, T.M.; Dawson, V.L.; El-Deiry, W.S.; Fulda, S.; et al. Molecular definitions of cell death subroutines: Recommendations of the nomenclature committee on cell death 2012. Cell Death Differ. 2012, 19, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Ichim, G.; Tait, S.W. A fate worse than death: Apoptosis as an oncogenic process. Nat. Rev. Cancer 2016, 16, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, R.; Zauli, G. Emerging non-apoptotic functions of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)/Apo2L. J. Cell. Physiol. 2004, 201, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Chipuk, J.E.; Bouchier-Hayes, L.; Green, D.R. Mitochondrial outer membrane permeabilization during apoptosis: The innocent bystander scenario. Cell Death Differ. 2006, 13, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Tait, S.W.; Green, D.R. Mitochondria and cell death: Outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Dewson, G.; Kratina, T.; Sim, H.W.; Puthalakath, H.; Adams, J.M.; Colman, P.M.; Kluck, R.M. To trigger apoptosis, Bak exposes its BH3 domain and homodimerizes via BH3: Groove interactions. Mol. Cell 2008, 30, 369–380. [Google Scholar] [CrossRef] [PubMed]
- George, N.M.; Evans, J.J.; Luo, X. A three-helix homo-oligomerization domain containing BH3 and BH1 is responsible for the apoptotic activity of Bax. Genes Dev. 2007, 21, 1937–1948. [Google Scholar] [CrossRef] [PubMed]
- Papoff, G.; Hausler, P.; Eramo, A.; Pagano, M.G.; Di Leve, G.; Signore, A.; Ruberti, G. Identification and characterization of a ligand-independent oligomerization domain in the extracellular region of the CD95 death receptor. J. Biol. Chem. 1999, 274, 38241–38250. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.M.; Frederiksen, J.K.; Zacharias, D.A.; Chan, F.K.; Johnson, M.; Lynch, D.; Tsien, R.Y.; Lenardo, M.J. Fas preassociation required for apoptosis signaling and dominant inhibition by pathogenic mutations. Science 2000, 288, 2354–2357. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Gores, G.J. Death receptor-mediated apoptosis and the liver. J. Hepatol. 2002, 37, 400–410. [Google Scholar] [CrossRef]
- Martin-Villalba, A.; Llorens-Bobadilla, E.; Wollny, D. CD95 in cancer: Tool or target? Trends Mol. Med. 2013, 19, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.E.; Hadji, A.; Murmann, A.E.; Brockway, S.; Putzbach, W.; Pattanayak, A.; Ceppi, P. The role of CD95 and CD95 ligand in cancer. Cell Death Differ. 2015, 22, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Shepard, B.D.; Badley, A.D. The biology of TRAIL and the role of TRAIL-based therapeutics in infectious diseases. Antiinfect. Agents Med. Chem. 2009, 8, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Kantari, C.; Walczak, H. Caspase-8 and bid: Caught in the act between death receptors and mitochondria. Biochim. Biophys. Acta 2011, 1813, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Irmler, M.; Thome, M.; Hahne, M.; Schneider, P.; Hofmann, K.; Steiner, V.; Bodmer, J.L.; Schroter, M.; Burns, K.; Mattmann, C.; et al. Inhibition of death receptor signals by cellular FLIP. Nature 1997, 388, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Strasser, A.; Jost, P.J.; Nagata, S. The many roles of FAS receptor signaling in the immune system. Immunity 2009, 30, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.E.; Krammer, P.H. The CD95(APO-1/Fas) DISC and beyond. Cell Death Differ. 2003, 10, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.A.; Powley, I.R.; Jukes-Jones, R.; Horn, S.; Feoktistova, M.; Fairall, L.; Schwabe, J.W.; Leverkus, M.; Cain, K.; MacFarlane, M. Co-operative and hierarchical binding of c-FLIP and Caspase-8: A unified Model defines how c-FLIP Isoforms differentially control cell fate. Mol. Cell 2016, 61, 834–849. [Google Scholar] [CrossRef] [PubMed]
- Wajant, H.; Pfizenmaier, K.; Scheurich, P. Tumor necrosis factor signaling. Cell Death Differ. 2003, 10, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, X.; Pang, X.; Ding, L.; Wood, O.; Clouse, K.; Hewlett, I.; Dayton, A.I. Identification of a potential HIV-induced source of bystander-mediated apoptosis in T cells: Upregulation of trail in primary human macrophages by HIV-1 tat. J. Biomed. Sci. 2001, 8, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Westendorp, M.O.; Frank, R.; Ochsenbauer, C.; Stricker, K.; Dhein, J.; Walczak, H.; Debatin, K.M.; Krammer, P.H. Sensitization of T cells to CD95-mediated apoptosis by HIV-1 Tat and gp120. Nature 1995, 375, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Tikhonov, I.; Ruckwardt, T.J.; Djavani, M.; Zapata, J.C.; Pauza, C.D.; Salvato, M.S. Monocytes treated with human immunodeficiency virus Tat kill uninfected CD4(+) cells by a tumor necrosis factor-related apoptosis-induced ligand-mediated mechanism. J. Virol. 2003, 77, 6700–6708. [Google Scholar] [CrossRef] [PubMed]
- Algeciras-Schimnich, A.; Vlahakis, S.R.; Villasis-Keever, A.; Gomez, T.; Heppelmann, C.J.; Bou, G.; Paya, C.V. CCR5 mediates Fas- and caspase-8 dependent apoptosis of both uninfected and HIV infected primary human CD4 T cells. AIDS 2002, 16, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- Casella, C.R.; Rapaport, E.L.; Finkel, T.H. Vpu increases susceptibility of human immunodeficiency virus type 1-infected cells to fas killing. J. Virol. 1999, 73, 92–100. [Google Scholar] [PubMed]
- Boirivant, M.; Viora, M.; Giordani, L.; Luzzati, A.L.; Pronio, A.M.; Montesani, C.; Pugliese, O. HIV-1 gp120 accelerates Fas-mediated activation-induced human lamina propria T cell apoptosis. J. Clin. Immunol. 1998, 18, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Herbein, G.; Mahlknecht, U.; Batliwalla, F.; Gregersen, P.; Pappas, T.; Butler, J.; O’Brien, W.A.; Verdin, E. Apoptosis of CD8+ T cells is mediated by macrophages through interaction of HIV gp120 with chemokine receptor CXCR4. Nature 1998, 395, 189–194. [Google Scholar] [PubMed]
- Micoli, K.J.; Pan, G.; Wu, Y.; Williams, J.P.; Cook, W.J.; McDonald, J.M. Requirement of calmodulin binding by HIV-1 gp160 for enhanced FAS-mediated apoptosis. J. Biol. Chem. 2000, 275, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Huang, C.; Wang, Y.; Lu, Z.; Zhuang, N.; Zhao, D.; He, J.; Shi, L. Hepatitis B Virus X protein sensitizes TRAIL-induced Hepatocyte apoptosis by inhibiting the E3 ubiquitin ligase A20. PLoS ONE 2015, 10, e0127329. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; You, H.; Zhao, J.; Liu, W.; Hu, L.; Luo, W.; Hu, W.; Tang, R.; Zheng, K. The enhanced expression of death receptor 5 (DR5) mediated by HBV X protein through NF-κB pathway is associated with cell apoptosis induced by (TNF-α related apoptosis inducing ligand) TRAIL in hepatoma cells. Virol. J. 2015, 12, 192. [Google Scholar] [CrossRef] [PubMed]
- Hahn, C.S.; Cho, Y.G.; Kang, B.S.; Lester, I.M.; Hahn, Y.S. The HCV core protein acts as a positive regulator of Fas-mediated apoptosis in a human lymphoblastoid T cell line. Virology 2000, 276, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Ruggieri, A.; Murdolo, M.; Rapicetta, M. Induction of FAS ligand expression in a human hepatoblastoma cell line by HCV core protein. Virus Res. 2003, 97, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Khoshnan, A.; Schneider, R.; Matsumoto, M.; Dennert, G.; Ware, C.; Lai, M.M. Hepatitis C virus core protein binds to the cytoplasmic domain of tumor necrosis factor (TNF) receptor 1 and enhances TNF-induced apoptosis. J. Virol. 1998, 72, 3691–3697. [Google Scholar] [PubMed]
- Chou, A.H.; Tsai, H.F.; Wu, Y.Y.; Hu, C.Y.; Hwang, L.H.; Hsu, P.I.; Hsu, P.N. Hepatitis C virus core protein modulates TRAIL-mediated apoptosis by enhancing Bid cleavage and activation of mitochondria apoptosis signaling pathway. J. Immunol. 2005, 174, 2160–2166. [Google Scholar] [CrossRef] [PubMed]
- Incrocci, R.; Hussain, S.; Stone, A.; Bieging, K.; Alt, L.A.; Fay, M.J.; Swanson-Mungerson, M. Epstein-barr virus latent membrane protein 2A (LMP2A)-mediated changes in Fas expression and Fas-dependent apoptosis: Role of Lyn/Syk activation. Cell. Immunol. 2015, 297, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Le Clorennec, C.; Youlyouz-Marfak, I.; Adriaenssens, E.; Coll, J.; Bornkamm, G.W.; Feuillard, J. EBV latency III immortalization program sensitizes B cells to induction of CD95-mediated apoptosis via LMP1: Role of NF-κB, STAT1, and p53. Blood 2006, 107, 2070–2078. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Fang, Y.; Sima, N.; Li, Y.; Li, W.; Li, L.; Han, L.; Liao, S.; Han, Z.; Gao, Q.; et al. Triggering of death receptor apoptotic signaling by human papillomavirus 16 E2 protein in cervical cancer cell lines is mediated by interaction with c-FLIP. Apoptosis 2011, 16, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Stoppler, H.; Stoppler, M.C.; Johnson, E.; Simbulan-Rosenthal, C.M.; Smulson, M.E.; Iyer, S.; Rosenthal, D.S.; Schlegel, R. The E7 protein of human papillomavirus type 16 sensitizes primary human keratinocytes to apoptosis. Oncogene 1998, 17, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Walsh, I.; Waterfield, M.; Xiao, G.; Fong, A.; Sun, S.C. NF-κB signaling pathway governs TRAIL gene expression and human T-cell leukemia virus-I Tax-induced T-cell death. J. Biol. Chem. 2001, 276, 40385–40388. [Google Scholar] [CrossRef] [PubMed]
- Kassis, R.; Larrous, F.; Estaquier, J.; Bourhy, H. Lyssavirus matrix protein induces apoptosis by a TRAIL-dependent mechanism involving caspase-8 activation. J. Virol. 2004, 78, 6543–6555. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.A.; Norris, P.S.; Ware, C.F. To kill or be killed: Viral evasion of apoptosis. Nat. Immunol. 2002, 3, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Davis, T.; Anderson, D.; Solam, L.; Beckmann, M.P.; Jerzy, R.; Dower, S.K.; Cosman, D.; Goodwin, R.G. A receptor for tumor necrosis factor defines an unusual family of cellular and viral proteins. Science 1990, 248, 1019–1023. [Google Scholar] [CrossRef] [PubMed]
- Reading, P.C.; Khanna, A.; Smith, G.L. Vaccinia virus CrmE encodes a soluble and cell surface tumor necrosis factor receptor that contributes to virus virulence. Virology 2002, 292, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Macen, J.L.; Garner, R.S.; Musy, P.Y.; Brooks, M.A.; Turner, P.C.; Moyer, R.W.; McFadden, G.; Bleackley, R.C. Differential inhibition of the Fas- and granule-mediated cytolysis pathways by the orthopoxvirus cytokine response modifier A/SPI-2 and SPI-1 protein. Proc. Natl. Acad. Sci. USA 1996, 93, 9108–9113. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.J.; Silke, J.; Kenshole, B.; Ruby, J. Characterization of the ectromelia virus serpin, SPI-2. J. Gen. Virol. 2000, 81, 2425–2430. [Google Scholar] [CrossRef] [PubMed]
- Dobbelstein, M.; Shenk, T. Protection against apoptosis by the vaccinia virus SPI-2 (B13R) gene product. J. Virol. 1996, 70, 6479–6485. [Google Scholar] [PubMed]
- Veyer, D.L.; Carrara, G.; Maluquer de Motes, C.; Smith, G.L. Vaccinia virus evasion of regulated cell death. Immunol. Lett. 2017, 186, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.A.; Ali, A.N.; Turner, P.C.; Moyer, R.W. A rabbitpox virus serpin gene controls host range by inhibiting apoptosis in restrictive cells. J. Virol. 1995, 69, 7688–7698. [Google Scholar] [PubMed]
- Schreiber, M.; Sedger, L.; McFadden, G. Distinct domains of M-T2, the myxoma virus tumor necrosis factor (TNF) receptor homolog, mediate extracellular TNF binding and intracellular apoptosis inhibition. J. Virol. 1997, 71, 2171–2181. [Google Scholar] [PubMed]
- Srinivasula, S.M.; Ahmad, M.; Fernandes-Alnemri, T.; Litwack, G.; Alnemri, E.S. Molecular ordering of the Fas-apoptotic pathway: The Fas/APO-1 protease Mch5 is a CrmA-inhibitable protease that activates multiple Ced-3/ICE-like cysteine proteases. Proc. Natl. Acad. Sci. USA 1996, 93, 14486–14491. [Google Scholar] [CrossRef] [PubMed]
- Medici, M.A.; Sciortino, M.T.; Perri, D.; Amici, C.; Avitabile, E.; Ciotti, M.; Balestrieri, E.; De Smaele, E.; Franzoso, G.; Mastino, A. Protection by herpes simplex virus glycoprotein D against Fas-mediated apoptosis - Role of nuclear factor κB. J. Biol. Chem. 2003, 278, 36059–36067. [Google Scholar] [CrossRef] [PubMed]
- Dufour, F.; Sasseville, A.M.; Chabaud, S.; Massie, B.; Siegel, R.M.; Langelier, Y. The ribonucleotide reductase R1 subunits of herpes simplex virus types 1 and 2 protect cells against TNFα- and FasL-induced apoptosis by interacting with caspase-8. Apoptosis 2011, 16, 256–271. [Google Scholar] [CrossRef] [PubMed]
- Chiou, S.H.; Yang, Y.P.; Lin, J.C.; Hsu, C.H.; Jhang, H.C.; Yang, Y.T.; Lee, C.H.; Ho, L.L.; Hsu, W.M.; Ku, H.H.; et al. The immediate early 2 protein of human cytomegalovirus (HCMV) mediates the apoptotic control in HCMV retinitis through up-regulation of the cellular FLICE-inhibitory protein expression. J. Immunol. 2006, 177, 6199–6206. [Google Scholar] [CrossRef] [PubMed]
- Goldmacher, V.S.; Bartle, L.M.; Skaletskaya, A.; Dionne, C.A.; Kedersha, N.L.; Vater, C.A.; Han, J.W.; Lutz, R.J.; Watanabe, S.; Cahir McFarland, E.D.; et al. A cytomegalovirus-encoded mitochondria-localized inhibitor of apoptosis structurally unrelated to BCL-2. Proc. Natl. Acad. Sci. USA 1999, 96, 12536–12541. [Google Scholar] [CrossRef] [PubMed]
- Skaletskaya, A.; Bartle, L.M.; Chittenden, T.; McCormick, A.L.; Mocarski, E.S.; Goldmacher, V.S. A cytomegalovirus-encoded inhibitor of apoptosis that suppresses caspase-8 activation. Proc. Natl. Acad. Sci. USA 2001, 98, 7829–7834. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.L.; Skaletskaya, A.; Barry, P.A.; Mocarski, E.S.; Goldmacher, V.S. Differential function and expression of the viral inhibitor of caspase 8-induced apoptosis (vICA) and the viral mitochondria-localized inhibitor of apoptosis (vMIA) cell death suppressors conserved in primate and rodent cytomegaloviruses. Virology 2003, 316, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Mack, C.; Sickmann, A.; Lembo, D.; Brune, W. Inhibition of proinflammatory and innate immune signaling pathways by a cytomegalovirus RIP1-interacting protein. Proc. Natl. Acad. Sci. USA 2008, 105, 3094–3099. [Google Scholar] [CrossRef] [PubMed]
- Belanger, C.; Gravel, A.; Tomoiu, A.; Janelle, M.E.; Gosselin, J.; Tremblay, M.J.; Flamand, L. Human herpesvirus 8 viral FLICE-inhibitory protein inhibits Fas-mediated apoptosis through binding and prevention of procaspase-8 maturation. J. Hum. Virol. 2001, 4, 62–73. [Google Scholar] [PubMed]
- Thome, M.; Schneider, P.; Hofmann, K.; Fickenscher, H.; Meinl, E.; Neipel, F.; Mattmann, C.; Burns, K.; Bodmer, J.L.; Schroter, M.; et al. Viral FLICE-inhibitory proteins (FLIPs) prevent apoptosis induced by death receptors. Nature 1997, 386, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Bertin, J.; Armstrong, R.C.; Ottilie, S.; Martin, D.A.; Wang, Y.; Banks, S.; Wang, G.H.; Senkevich, T.G.; Alnemri, E.S.; Moss, B.; et al. Death effector domain-containing herpesvirus and poxvirus proteins inhibit both Fas- and TNFR1-induced apoptosis. Proc. Natl. Acad. Sci. USA 1997, 94, 1172–1176. [Google Scholar] [CrossRef] [PubMed]
- Thurau, M.; Everett, H.; Tapernoux, M.; Tschopp, J.; Thome, M. The TRAF3-binding site of human molluscipox virus FLIP molecule MC159 is critical for its capacity to inhibit Fas-induced apoptosis. Cell Death Differ. 2006, 13, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Nanbo, A.; Yoshiyama, H.; Takada, K. Epstein-Barr virus-encoded poly(A)- RNA confers resistance to apoptosis mediated through Fas by blocking the PKR pathway in human epithelial intestine 407 cells. J. Virol. 2005, 79, 12280–12285. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, M. Epstein-Barr virus BHRF1 protein protects intestine 407 epithelial cells from apoptosis induced by tumor necrosis factor α and anti-Fas antibody. J. Virol. 1997, 71, 3319–3322. [Google Scholar] [PubMed]
- Foghsgaard, L.; Jaattela, M. The ability of BHRF1 to inhibit apoptosis is dependent on stimulus and cell type. J. Virol. 1997, 71, 7509–7517. [Google Scholar] [PubMed]
- Morrison, T.E.; Mauser, A.; Klingelhutz, A.; Kenney, S.C. Epstein-Barr virus immediate-early protein BZLF1 inhibits tumor necrosis factor α-induced signaling and apoptosis by downregulating tumor necrosis factor receptor 1. J. Virol. 2004, 78, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Snow, A.L.; Lambert, S.L.; Natkunam, Y.; Esquivel, C.O.; Krams, S.M.; Martinez, O.M. EBV can protect latently infected B cell lymphomas from death receptor-induced apoptosis. J. Immunol. 2006, 177, 3283–3293. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Yang, S.; Wang, S.; Yang, X.M.; Tang, Q.L.; Wang, S.H. Latent membrane protein 1 mediates the resistance of nasopharyngeal carcinoma cells to TRAIL-induced apoptosis by activation of the PI3K/Akt signaling pathway. Oncol. Rep. 2011, 26, 1573–1579. [Google Scholar] [PubMed]
- Wang, G.H.; Garvey, T.L.; Cohen, J.I. The murine gammaherpesvirus-68 M11 protein inhibits Fas- and TNF-induced apoptosis. J. Gen. Virol. 1999, 80 Pt 10, 2737–2740. [Google Scholar] [CrossRef] [PubMed]
- Marusawa, H.; Hijikata, M.; Chiba, T.; Shimotohno, K. Hepatitis C virus core protein inhibits Fas- and tumor necrosis factor α-mediated apoptosis via NF-κB activation. J. Virol. 1999, 73, 4713–4720. [Google Scholar] [PubMed]
- Ray, R.B.; Meyer, K.; Steele, R.; Shrivastava, A.; Aggarwal, B.B.; Ray, R. Inhibition of tumor necrosis factor (TNF-α)-mediated apoptosis by hepatitis C virus core protein. J. Biol. Chem. 1998, 273, 2256–2259. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ray, R. Evasion of TNF-α-mediated apoptosis by hepatitis C virus. Methods Mol. Biol. 2014, 1155, 125–132. [Google Scholar] [PubMed]
- Chen, Z.H.; Zhu, Y.Z.; Ren, Y.L.; Tong, Y.M.; Hua, X.; Zhu, F.H.; Huang, L.B.; Liu, Y.; Luo, Y.; Lu, W.; et al. Hepatitis C virus Protects human B lymphocytes from Fas-mediated apoptosis via E2-CD81 engagement. PLoS ONE 2011, 6, e18933. [Google Scholar] [CrossRef] [PubMed]
- Majumder, M.; Ghosh, A.K.; Steele, R.; Zhou, X.Y.; Phillips, N.J.; Ray, R.; Ray, R.B. Hepatitis C virus NS5A protein impairs TNF-mediated hepatic apoptosis, but not by an anti-FAS antibody, in transgenic mice. Virology 2002, 294, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Lin, Y.T.; Yan, X.L.; Ding, Y.L.; Wu, Y.L.; Chen, W.N.; Lin, X. Hepatitis B virus core protein inhibits Fas-mediated apoptosis of hepatoma cells via regulation of mFas/FasL and sFas expression. FASEB J. 2015, 29, 1113–1123. [Google Scholar] [CrossRef] [PubMed]
- Shisler, J.; Yang, C.; Walter, B.; Ware, C.F.; Gooding, L.R. The adenovirus E3-10.4K/14.5K complex mediates loss of cell surface Fas (CD95) and resistance to Fas-induced apoptosis. J. Virol. 1997, 71, 8299–8306. [Google Scholar] [PubMed]
- Benedict, C.A.; Norris, P.S.; Prigozy, T.I.; Bodmer, J.L.; Mahr, J.A.; Garnett, C.T.; Martinon, F.; Tschopp, J.; Gooding, L.R.; Ware, C.F. Three adenovirus E3 proteins cooperate to evade apoptosis by tumor necrosis factor-related apoptosis-inducing ligand receptor-1 and -2. J. Biol. Chem. 2001, 276, 3270–3278. [Google Scholar] [CrossRef] [PubMed]
- McNees, A.L.; Garnett, C.T.; Gooding, L.R. The adenovirus E3 RID complex protects some cultured human T and B lymphocytes from Fas-induced apoptosis. J. Virol. 2002, 76, 9716–9723. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Brachert, W.; Tchikov, V.; Merkel, O.; Jakob, M.; Hallas, C.; Kruse, M.L.; Groitl, P.; Lehn, A.; Hildt, E.; Held-Feindt, J.; et al. Inhibition of TNF receptor 1 internalization by adenovirus 14.7K as a novel immune escape mechanism. J. Clin. Investig. 2006, 116, 2901–2913. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, K.; Alonso, A. The human papillomavirus type 16 E5 protein impairs TRAIL- and FasL-mediated apoptosis in HaCaT cells by different mechanisms. J. Virol. 2002, 76, 12162–12172. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Martinez, A.; Madrid-Marina, V.; Gariglio, P. Modulation of apoptosis by early human papillomavirus proteins in cervical cancer. Biochim. Biophys. Acta 2010, 1805, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Garnett, T.O.; Filippova, M.; Duerksen-Hughes, P.J. Accelerated degradation of FADD and procaspase 8 in cells expressing human papilloma virus 16 E6 impairs TRAIL-mediated apoptosis. Cell Death Differ. 2006, 13, 1915–1926. [Google Scholar] [CrossRef] [PubMed]
- Filippova, M.; Song, H.; Connolly, J.L.; Dermody, T.S.; Duerksen-Hughes, P.J. The human papillomavirus 16 E6 protein binds to tumor necrosis factor (TNF) R1 and protects cells from TNF-induced apoptosis. J. Biol. Chem. 2002, 277, 21730–21739. [Google Scholar] [CrossRef] [PubMed]
- Duerksen-Hughes, P.J.; Yang, J.; Schwartz, S.B. HPV 16 E6 blocks TNF-mediated apoptosis in mouse fibroblast LM cells. Virology 1999, 264, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Filippova, M.; Filippov, V.A.; Kagoda, M.; Garnett, T.; Fodor, N.; Duerksen-Hughes, P.J. Complexes of human papillomavirus type 16 E6 proteins form pseudo-death-inducing signaling complex structures during tumor necrosis factor-mediated apoptosis. J. Virol. 2009, 83, 210–227. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.A.; Zacny, V.; Belinsky, G.S.; Classon, M.; Jones, D.L.; Schlegel, R.; Munger, K. The HPV E7 oncoprotein inhibits tumor necrosis factor α-mediated apoptosis in normal human fibroblasts. Oncogene 2001, 20, 3629–3640. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Fu, F.; Zhuo, J.; Wang, W.; Nishitani, J.; An, D.S.; Chen, I.S.; Liu, X. Human papillomavirus type 16 E6 and E7 oncoproteins upregulate c-IAP2 gene expression and confer resistance to apoptosis. Oncogene 2005, 24, 5069–5078. [Google Scholar] [CrossRef] [PubMed]
- Noyori, O.; Nakayama, E.; Maruyama, J.; Yoshida, R.; Takada, A. Suppression of Fas-mediated apoptosis via steric shielding by filovirus glycoproteins. Biochem. Biophys. Res. Commun. 2013, 441, 994–998. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.; Jeong, J.G.; Kim, S. Stable expression of human immunodeficiency virus type 1 Nef confers resistance against Fas-mediated apoptosis. AIDS Res. Hum. Retrov. 2001, 17, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Gibellini, D.; Re, M.C.; Ponti, C.; Maldini, C.; Celeghini, C.; Cappellini, A.; La Placa, M.; Zauli, G. HIV-1 Tat protects CD4+ Jurkat T lymphoblastoid cells from apoptosis mediated by TNF-related apoptosis-inducing ligand. Cell. Immunol. 2001, 207, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Krueger, A.; Fas, S.C.; Giaisi, M.; Bleumink, M.; Merling, A.; Stumpf, C.; Baumann, S.; Holtkotte, D.; Bosch, V.; Krammer, P.H.; et al. HTLV-1 Tax protects against CD95-mediated apoptosis by induction of the cellular FLICE-inhibitory protein (c-FLIP). Blood 2006, 107, 3933–3939. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Fujisawa, J.; Reth, M.; Yonehara, S. Human T-cell leukemia virus type-I oncoprotein Tax inhibits Fas-mediated apoptosis by inducing cellular FLIP through activation of NF-κB. Genes Cells 2006, 11, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Zehender, G.; Varchetta, S.; de Maddalena, C.; Colasante, C.; Riva, A.; Meroni, L.; Moroni, M.; Galli, M. Resistance to Fas-mediated apoptosis of human T-cell lines expressing human T-lymphotropic virus type-2 (HTLV-2) Tax protein. Virology 2001, 281, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Aries, S.P.; Schaaf, B.; Muller, C.; Dennin, R.H.; Dalhoff, K. Fas (CD95) expression on CD4+ T cells from HIV-infected patients increases with disease progression. J. Mol. Med. 1995, 73, 591–593. [Google Scholar] [CrossRef] [PubMed]
- Sloand, E.M.; Young, N.S.; Kumar, P.; Weichold, F.F.; Sato, T.; Maciejewski, J.P. Role of Fas ligand and receptor in the mechanism of T-cell depletion in acquired immunodeficiency syndrome: Effect on CD4+ lymphocyte depletion and human immunodeficiency virus replication. Blood 1997, 89, 1357–1363. [Google Scholar] [PubMed]
- Baumler, C.B.; Bohler, T.; Herr, I.; Benner, A.; Krammer, P.H.; Debatin, K.M. Activation of the CD95 (APO-1/Fas) system in T cells from human immunodeficiency virus type-1-infected children. Blood 1996, 88, 1741–1746. [Google Scholar] [PubMed]
- McCloskey, T.W.; Oyaizu, N.; Kaplan, M.; Pahwa, S. Expression of the Fas antigen in patients infected with human immunodeficiency virus. Cytometry 1995, 22, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Gehri, R.; Hahn, S.; Rothen, M.; Steuerwald, M.; Nuesch, R.; Erb, P. The Fas receptor in HIV infection: Expression on peripheral blood lymphocytes and role in the depletion of T cells. AIDS 1996, 10, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Boudet, F.; Lecoeur, H.; Gougeon, M.L. Apoptosis associated with ex vivo down-regulation of BCL-2 and up-regulation of Fas in potential cytotoxic CD8+ T lymphocytes during HIV infection. J. Immunol. 1996, 156, 2282–2293. [Google Scholar] [PubMed]
- Katsikis, P.D.; Wunderlich, E.S.; Smith, C.A.; Herzenberg, L.A.; Herzenberg, L.A. Fas antigen stimulation induces marked apoptosis of T lymphocytes in human immunodeficiency virus-infected individuals. J. Exp. Med. 1995, 181, 2029–2036. [Google Scholar] [CrossRef] [PubMed]
- Badley, A.D.; McElhinny, J.A.; Leibson, P.J.; Lynch, D.H.; Alderson, M.R.; Paya, C.V. Upregulation of Fas ligand expression by human immunodeficiency virus in human macrophages mediates apoptosis of uninfected T lymphocytes. J. Virol. 1996, 70, 199–206. [Google Scholar] [PubMed]
- Herbeuval, J.P.; Grivel, J.C.; Boasso, A.; Hardy, A.W.; Chougnet, C.; Dolan, M.J.; Yagita, H.; Lifson, J.D.; Shearer, G.M. CD4+ T-cell death induced by infectious and noninfectious HIV-1: Role of type 1 interferon-dependent, TRAIL/DR5-mediated apoptosis. Blood 2005, 106, 3524–3531. [Google Scholar] [CrossRef] [PubMed]
- Herbeuval, J.P.; Hardy, A.W.; Boasso, A.; Anderson, S.A.; Dolan, M.J.; Dy, M.; Shearer, G.M. Regulation of TNF-related apoptosis-inducing ligand on primary CD4+ T cells by HIV-1: Role of type I IFN-producing plasmacytoid dendritic cells. Proc. Natl. Acad. Sci. USA 2005, 102, 13974–13979. [Google Scholar] [CrossRef] [PubMed]
- Miura, Y.; Koyanagi, Y.; Mizusawa, H. TNF-related apoptosis-inducing ligand (TRAIL) induces neuronal apoptosis in HIV-encephalopathy. J. Med. Dent. Sci. 2003, 50, 17–25. [Google Scholar] [PubMed]
- Miura, Y.; Misawa, N.; Maeda, N.; Inagaki, Y.; Tanaka, Y.; Ito, M.; Kayagaki, N.; Yamamoto, N.; Yagita, H.; Mizusawa, H.; et al. Critical contribution of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) to apoptosis of human CD4+ T cells in HIV-1-infected hu-PBL-NOD-SCID mice. J. Exp. Med. 2001, 193, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Dabrowska, A.; Jenner, R.G.; Aldovini, A. Human and simian immunodeficiency virus-mediated upregulation of the apoptotic factor TRAIL occurs in antigen-presenting cells from AIDS-susceptible but not from AIDS-resistant species. J. Virol. 2007, 81, 7584–7597. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Erdmann, N.; Peng, H.; Herek, S.; Davis, J.S.; Luo, X.; Ikezu, T.; Zheng, J. TRAIL-mediated apoptosis in HIV-1-infected macrophages is dependent on the inhibition of Akt-1 phosphorylation. J. Immunol. 2006, 177, 2304–2313. [Google Scholar] [CrossRef] [PubMed]
- Van Grevenynghe, J.; Cubas, R.A.; Noto, A.; DaFonseca, S.; He, Z.; Peretz, Y.; Filali-Mouhim, A.; Dupuy, F.P.; Procopio, F.A.; Chomont, N.; et al. Loss of memory B cells during chronic HIV infection is driven by Foxo3a- and TRAIL-mediated apoptosis. J. Clin. Investig. 2011, 121, 3877–3888. [Google Scholar] [CrossRef] [PubMed]
- Mundt, B.; Kuhnel, F.; Zender, L.; Paul, Y.; Tillmann, H.; Trautwein, C.; Manns, M.P.; Kubicka, S. Involvement of TRAIL and its receptors in viral hepatitis. FASEB J. 2003, 17, 94–96. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, F.; Pontisso, P.; Pettenazzo, E.; Benvegnu, L.; Vario, A.; Chemello, L.; Alberti, A.; Valente, M. Liver cell apoptosis in chronic hepatitis C correlates with histological but not biochemical activity or serum HCV-RNA levels. Hepatology 2000, 31, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Mita, E.; Hayashi, N.; Iio, S.; Takehara, T.; Hijioka, T.; Kasahara, A.; Fusamoto, H.; Kamada, T. Role of Fas ligand in apoptosis induced by hepatitis C virus infection. Biochem. Biophys. Res. Commun. 1994, 204, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Taya, N.; Torimoto, Y.; Shindo, M.; Hirai, K.; Hasebe, C.; Kohgo, Y. Fas-mediated apoptosis of peripheral blood mononuclear cells in patients with hepatitis C. Br. J. Haematol. 2000, 110, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Lan, L.; Gorke, S.; Rau, S.J.; Zeisel, M.B.; Hildt, E.; Himmelsbach, K.; Carvajal-Yepes, M.; Huber, R.; Wakita, T.; Schmitt-Graeff, A.; et al. Hepatitis C virus infection sensitizes human hepatocytes to TRAIL-induced apoptosis in a caspase 9-dependent manner. J. Immunol. 2008, 181, 4926–4935. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Yan, H.; Hu, J.; Zhang, S.; Peng, P.; Liu, Q.; Guo, D. Hepatitis C virus sensitizes host cells to TRAIL-induced apoptosis by up-regulating DR4 and DR5 via a MEK1-dependent pathway. PLoS ONE 2012, 7, e37700. [Google Scholar] [CrossRef] [PubMed]
- Silberstein, E.; Ulitzky, L.; Lima, L.A.; Cehan, N.; Teixeira-Carvalho, A.; Roingeard, P.; Taylor, D.R. HCV-mediated Apoptosis of hepatocytes in culture and viral pathogenesis. PLoS ONE 2016, 11, e0155708. [Google Scholar] [CrossRef] [PubMed]
- Janssen, H.L.; Higuchi, H.; Abdulkarim, A.; Gores, G.J. Hepatitis B virus enhances tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) cytotoxicity by increasing TRAIL-R1/death receptor 4 expression. J. Hepatol. 2003, 39, 414–420. [Google Scholar] [CrossRef]
- Fischer, R.; Baumert, T.; Blum, H.E. Hepatitis C virus infection and apoptosis. World J. Gastroenterol. 2007, 13, 4865–4872. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Watanabe, M.; Ihara, T.; Kamiya, H.; Sakurai, M. Fas antigen and BCL-2 expression on lymphocytes cultured with cytomegalovirus and varicella-zoster virus antigen. Cell. Immunol. 1995, 160, 173–177. [Google Scholar] [CrossRef]
- Brazeau, E.; Mahalingam, R.; Gilden, D.; Wellish, M.; Kaufer, B.B.; Osterrieder, N.; Pugazhenthi, S. Varicella-zoster virus-induced apoptosis in MeWo cells is accompanied by down-regulation of BCL-2 expression. J. Neurovirol. 2010, 16, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Ando, K.; Tanaka, K.; Ikeda, Y.; Koga, Y. Fas-mediated apoptosis of the hematopoietic progenitor cells in mice infected with murine cytomegalovirus. Blood 1997, 89, 3565–3573. [Google Scholar] [PubMed]
- Poole, E.; Lau, J.C.; Sinclair, J. Latent infection of myeloid progenitors by human cytomegalovirus protects cells from FAS-mediated apoptosis through the cellular IL-10/PEA-15 pathway. J. Gen. Virol. 2015, 96, 2355–2359. [Google Scholar] [CrossRef] [PubMed]
- Seirafian, S.; Prod’homme, V.; Sugrue, D.; Davies, J.; Fielding, C.; Tomasec, P.; Wilkinson, G.W. Human cytomegalovirus suppresses Fas expression and function. J. Gen. Virol. 2014, 95, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Philip, P.S.; Attoub, S.; Khan, G. Epstein-Barr virus-infected cells release Fas ligand in exosomal fractions and induce apoptosis in recipient cells via the extrinsic pathway. J. Gen. Virol. 2015, 96, 3646–3659. [Google Scholar] [CrossRef] [PubMed]
- Tanner, J.E.; Alfieri, C. Epstein-barr virus induces Fas (CD95) in T cells and Fas ligand in B cells leading to T-cell apoptosis. Blood 1999, 94, 3439–3447. [Google Scholar] [PubMed]
- Wada, N.; Matsumura, M.; Ohba, Y.; Kobayashi, N.; Takizawa, T.; Nakanishi, Y. Transcription stimulation of the Fas-encoding gene by nuclear factor for interleukin-6 expression upon influenza virus infection. J. Biol. Chem. 1995, 270, 18007–18012. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, T.; Matsukawa, S.; Higuchi, Y.; Nakamura, S.; Nakanishi, Y.; Fukuda, R. Induction of programmed cell death (apoptosis) by influenza virus infection in tissue culture cells. J. Gen. Virol. 1993, 74, 2347–2355. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, T.; Fukuda, R.; Miyawaki, T.; Ohashi, K.; Nakanishi, Y. Activation of the apoptotic Fas antigen-encoding gene upon influenza virus infection involving spontaneously produced β-interferon. Virology 1995, 209, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, I.; Takizawa, T.; Ohba, Y.; Nakanishi, Y. Co-expression of Fas and Fas-ligand on the surface of influenza virus-infected cells. Cell Death Differ. 1998, 5, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, E.; Nakazawa, M.; Yoshinari, M.; Minami, M. Role of tumor necrosis factor-related apoptosis-inducing ligand in immune response to influenza virus infection in mice. J. Virol. 2005, 79, 7658–7663. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, D.R.; Milligan, L.; Stark, J.M. Induction of CD95 (Fas) and apoptosis in respiratory epithelial cell cultures following respiratory syncytial virus infection. Virology 1999, 257, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Kotelkin, A.; Prikhod’ko, E.A.; Cohen, J.I.; Collins, P.L.; Bukreyev, A. Respiratory syncytial virus infection sensitizes cells to apoptosis mediated by tumor necrosis factor-related apoptosis-inducing ligand. J. Virol. 2003, 77, 9156–9172. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.; Meintzer, S.M.; Gibson, S.; Widmann, C.; Garrington, T.P.; Johnson, G.L.; Tyler, K.L. Reovirus-induced apoptosis is mediated by TRAIL. J. Virol. 2000, 74, 8135–8139. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.; Meintzer, S.M.; Moffitt, L.A.; Tyler, K.L. Two distinct phases of virus-induced nuclear factor κB regulation enhance tumor necrosis factor-related apoptosis-inducing ligand-mediated apoptosis in virus-infected cells. J. Biol. Chem. 2003, 278, 18092–18100. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.; Meintzer, S.M.; Spalding, A.C.; Johnson, G.L.; Tyler, K.L. Caspase 8-dependent sensitization of cancer cells to TRAIL-induced apoptosis following reovirus-infection. Oncogene 2001, 20, 6910–6919. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Wang, H.X.; Mao, X.; Fang, H.; Wang, H.; Li, Y.; Sun, Y.; Meng, C.; Tan, L.; Song, C.; et al. RIP1 is a central signaling protein in regulation of TNF-α/TRAIL mediated apoptosis and necroptosis during Newcastle disease virus infection. Oncotarget 2017, 8, 43201–43217. [Google Scholar] [PubMed]
- Ghosh, S.; Dutta, K.; Basu, A. Chandipura virus induces neuronal death through Fas-mediated extrinsic apoptotic pathway. J. Virol. 2013, 87, 12398–12406. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Xu, J.; Huang, J. FasL/Fas pathway is involved in dengue virus induced apoptosis of the vascular endothelial cells. J. Med. Virol. 2010, 82, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.; Leser, J.S.; Quick, E.D.; Dionne, K.R.; Beckham, J.D.; Tyler, K.L. Death receptor-mediated apoptotic signaling is activated in the brain following infection with West Nile virus in the absence of a peripheral immune response. J. Virol. 2014, 88, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- Olmo, I.G.; Carvalho, T.G.; Costa, V.V.; Alves-Silva, J.; Ferrari, C.Z.; Izidoro-Toledo, T.C.; da Silva, J.F.; Teixeira, A.L.; Souza, D.G.; Marques, J.T.; et al. Zika virus Promotes neuronal cell death in a non-cell autonomous manner by triggering the release of neurotoxic factors. Front. Immunol. 2017, 8, 1016. [Google Scholar] [CrossRef] [PubMed]
- Souza, B.S.; Sampaio, G.L.; Pereira, C.S.; Campos, G.S.; Sardi, S.I.; Freitas, L.A.; Figueira, C.P.; Paredes, B.D.; Nonaka, C.K.; Azevedo, C.M.; et al. Zika virus infection induces mitosis abnormalities and apoptotic cell death of human neural progenitor cells. Sci. Rep. 2016, 6, 39775. [Google Scholar] [CrossRef] [PubMed]
- Bradfute, S.B.; Swanson, P.E.; Smith, M.A.; Watanabe, E.; McDunn, J.E.; Hotchkiss, R.S.; Bavari, S. Mechanisms and consequences of ebolavirus-induced lymphocyte apoptosis. J. Immunol. 2010, 184, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Impagliazzo, A.; Milder, F.; Kuipers, H.; Wagner, M.V.; Zhu, X.; Hoffman, R.M.; van Meersbergen, R.; Huizingh, J.; Wanningen, P.; Verspuij, J.; et al. A stable trimeric influenza hemagglutinin stem as a broadly protective immunogen. Science 2015, 349, 1301–1306. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.S.; Hensley, L.E.; Geisbert, J.B.; Jahrling, P.B.; Geisbert, T.W. Depletion of peripheral blood T lymphocytes and NK cells during the course of ebola hemorrhagic Fever in cynomolgus macaques. Viral Immunol. 2004, 17, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Sarid, R.; Ben-Moshe, T.; Kazimirsky, G.; Weisberg, S.; Appel, E.; Kobiler, D.; Lustig, S.; Brodie, C. vFLIP protects PC-12 cells from apoptosis induced by Sindbis virus: Implications for the role of TNF-α. Cell Death Differ. 2001, 8, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.A.; Butrovich, K.D.; Lurain, N.S.; Corbeil, J.; Rooney, I.; Schneider, P.; Tschopp, J.; Ware, C.F. Cutting edge: A novel viral TNF receptor superfamily member in virulent strains of human cytomegalovirus. J. Immunol. 1999, 162, 6967–6970. [Google Scholar] [PubMed]
- Sedy, J.R.; Spear, P.G.; Ware, C.F. Cross-regulation between herpesviruses and the TNF superfamily members. Nat. Rev. Immunol. 2008, 8, 861–873. [Google Scholar] [CrossRef] [PubMed]
- Upton, J.W.; Kaiser, W.J.; Mocarski, E.S. Virus inhibition of RIP3-dependent necrosis. Cell Host Microbe 2010, 7, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Garnett, T.O.; Duerksen-Hughes, P.J. Modulation of apoptosis by human papillomavirus (HPV) oncoproteins. Arch. Virol. 2006, 151, 2321–2335. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Almasan, A. Genomic organization and transcriptional regulation of human Apo2/TRAIL gene. Biochem. Biophys. Res. Commun. 2000, 278, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Hida, S.; Takayanagi, H.; Yokochi, T.; Kayagaki, N.; Takeda, K.; Yagita, H.; Okumura, K.; Tanaka, N.; Taniguchi, T.; et al. Antiviral response by natural killer cells through TRAIL gene induction by IFN-α/β. Eur. J. Immunol. 2001, 31, 3138–3146. [Google Scholar] [CrossRef]
- Smyth, M.J.; Takeda, K.; Hayakawa, Y.; Peschon, J.J.; van den Brink, M.R.; Yagita, H. Nature’s TRAIL—On a path to cancer immunotherapy. Immunity 2003, 18, 1–6. [Google Scholar] [CrossRef]
- Stegmann, K.A.; Bjorkstrom, N.K.; Veber, H.; Ciesek, S.; Riese, P.; Wiegand, J.; Hadem, J.; Suneetha, P.V.; Jaroszewicz, J.; Wang, C.; et al. Interferon-α-induced TRAIL on natural killer cells is associated with control of hepatitis C virus infection. Gastroenterology 2010, 138, 1885–1897. [Google Scholar] [CrossRef] [PubMed]
- Miot, C.; Beaumont, E.; Duluc, D.; Le Guillou-Guillemette, H.; Preisser, L.; Garo, E.; Blanchard, S.; Hubert Fouchard, I.; Creminon, C.; Lamourette, P.; et al. IL-26 is overexpressed in chronically HCV-infected patients and enhances TRAIL-mediated cytotoxicity and interferon production by human NK cells. Gut 2015, 64, 1466–1475. [Google Scholar] [CrossRef] [PubMed]
- Peppa, D.; Gill, U.S.; Reynolds, G.; Easom, N.J.; Pallett, L.J.; Schurich, A.; Micco, L.; Nebbia, G.; Singh, H.D.; Adams, D.H.; et al. Up-regulation of a death receptor renders antiviral T cells susceptible to NK cell-mediated deletion. J. Exp. Med. 2013, 210, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Schuster, I.S.; Wikstrom, M.E.; Brizard, G.; Coudert, J.D.; Estcourt, M.J.; Manzur, M.; O’Reilly, L.A.; Smyth, M.J.; Trapani, J.A.; Hill, G.R.; et al. TRAIL+ NK cells control CD4+ T cell responses during chronic viral infection to limit autoimmunity. Immunity 2014, 41, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Nemcovicova, I.; Benedict, C.A.; Zajonc, D.M. Structure of human cytomegalovirus UL141 binding to TRAIL-R2 reveals novel, non-canonical death receptor interactions. PLoS Pathog. 2013, 9, e1003224. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.; Tomasec, P.; Aicheler, R.; Loewendorf, A.; Nemcovicova, I.; Wang, E.C.; Stanton, R.J.; Macauley, M.; Norris, P.; Willen, L.; et al. Human cytomegalovirus glycoprotein UL141 targets the TRAIL death receptors to thwart host innate antiviral defenses. Cell Host Microbe 2013, 13, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Loewendorf, A.; Wang, Q.; McDonald, B.; Redwood, A.; Benedict, C.A. Inhibition of the TRAIL death receptor by CMV reveals its importance in NK cell-mediated antiviral defense. PLoS Pathog. 2014, 10, e1004268. [Google Scholar] [CrossRef] [PubMed]
- Achard, C.; Guillerme, J.B.; Bruni, D.; Boisgerault, N.; Combredet, C.; Tangy, F.; Jouvenet, N.; Gregoire, M.; Fonteneau, J.F. Oncolytic measles virus induces tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-mediated cytotoxicity by human myeloid and plasmacytoid dendritic cells. Oncoimmunology 2017, 6, e1261240. [Google Scholar] [CrossRef] [PubMed]
- Chaperot, L.; Blum, A.; Manches, O.; Lui, G.; Angel, J.; Molens, J.P.; Plumas, J. Virus or TLR agonists induce TRAIL-mediated cytotoxic activity of plasmacytoid dendritic cells. J. Immunol. 2006, 176, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Stary, G.; Klein, I.; Kohlhofer, S.; Koszik, F.; Scherzer, T.; Mullauer, L.; Quendler, H.; Kohrgruber, N.; Stingl, G. Plasmacytoid dendritic cells express TRAIL and induce CD4+ T-cell apoptosis in HIV-1 viremic patients. Blood 2009, 114, 3854–3863. [Google Scholar] [CrossRef] [PubMed]
- Zajac, A.J.; Quinn, D.G.; Cohen, P.L.; Frelinger, J.A. Fas-dependent CD4+ cytotoxic T-cell-mediated pathogenesis during virus infection. Proc. Natl. Acad. Sci. USA 1996, 93, 14730–14735. [Google Scholar] [CrossRef] [PubMed]
- Medana, I.M.; Gallimore, A.; Oxenius, A.; Martinic, M.M.; Wekerle, H.; Neumann, H. MHC class I-restricted killing of neurons by virus-specific CD8+ T lymphocytes is effected through the Fas/FasL, but not the perforin pathway. Eur. J. Immunol. 2000, 30, 3623–3633. [Google Scholar] [CrossRef]
- Ando, K.; Hiroishi, K.; Kaneko, T.; Moriyama, T.; Muto, Y.; Kayagaki, N.; Yagita, H.; Okumura, K.; Imawari, M. Perforin, Fas/Fas ligand, and TNF-α pathways as specific and bystander killing mechanisms of hepatitis C virus-specific human CTL. J. Immunol. 1997, 158, 5283–5291. [Google Scholar] [PubMed]
- Gremion, C.; Grabscheid, B.; Wolk, B.; Moradpour, D.; Reichen, J.; Pichler, W.; Cerny, A. Cytotoxic T lymphocytes derived from patients with chronic hepatitis C virus infection kill bystander cells via Fas-FasL interaction. J. Virol. 2004, 78, 2152–2157. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.D.; Redchenko, I.; Williams, N.A.; Morgan, A.J. CD4+ T cells inhibit growth of Epstein-Barr virus-transformed B cells through CD95-CD95 ligand-mediated apoptosis. Int. Immunol. 1998, 10, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Zhang, C.; Dematos, J.; Kuang, L.; Jung, J.U.; Liang, X. CD95 Signaling Inhibits B Cell receptor-mediated γherpesvirus replication in apoptosis-resistant B lymphoma cells. J. Virol. 2016, 90, 9782–9796. [Google Scholar] [CrossRef] [PubMed]
- Topham, D.J.; Cardin, R.C.; Christensen, J.P.; Brooks, J.W.; Belz, G.T.; Doherty, P.C. Perforin and Fas in murine gammaherpesvirus-specific CD8(+) T cell control and morbidity. J. Gen. Virol. 2001, 82, 1971–1981. [Google Scholar] [CrossRef] [PubMed]
- Brincks, E.L.; Gurung, P.; Langlois, R.A.; Hemann, E.A.; Legge, K.L.; Griffith, T.S. The magnitude of the T cell response to a clinically significant dose of influenza virus is regulated by TRAIL. J. Immunol. 2011, 187, 4581–4588. [Google Scholar] [CrossRef] [PubMed]
- Brincks, E.L.; Katewa, A.; Kucaba, T.A.; Griffith, T.S.; Legge, K.L. CD8 T cells utilize TRAIL to control influenza virus infection. J. Immunol. 2008, 181, 4918–4925. [Google Scholar] [CrossRef] [PubMed]
- Mueller, Y.M.; De Rosa, S.C.; Hutton, J.A.; Witek, J.; Roederer, M.; Altman, J.D.; Katsikis, P.D. Increased CD95/Fas-induced apoptosis of HIV-specific CD8(+) T cells. Immunity 2001, 15, 871–882. [Google Scholar] [CrossRef]
- Liu, Z.X.; Govindarajan, S.; Okamoto, S.; Dennert, G. Fas-mediated apoptosis causes elimination of virus-specific cytotoxic T cells in the virus-infected liver. J. Immunol. 2001, 166, 3035–3041. [Google Scholar] [CrossRef] [PubMed]
- Contini, P.; Ghio, M.; Merlo, A.; Brenci, S.; Filaci, G.; Indiveri, F.; Puppo, F. Soluble HLA class I/CD8 ligation triggers apoptosis in EBV-specific CD8+ cytotoxic T lymphocytes by Fas/Fas-ligand interaction. Hum. Immunol. 2000, 61, 1347–1351. [Google Scholar] [CrossRef]
- Krzyzowska, M.; Shestakov, A.; Eriksson, K.; Chiodi, F. Role of Fas/FasL in regulation of inflammation in vaginal tissue during HSV-2 infection. Cell Death Dis. 2011, 2, e132. [Google Scholar] [CrossRef] [PubMed]
- Krzyzowska, M.; Baska, P.; Orlowski, P.; Zdanowski, R.; Winnicka, A.; Eriksson, K.; Stankiewicz, W. HSV-2 regulates monocyte inflammatory response via the Fas/FasL pathway. PLoS ONE 2013, 8, e70308. [Google Scholar] [CrossRef] [PubMed]
- Krzyzowska, M.; Baska, P.; Grochowska, A.; Orlowski, P.; Nowak, Z.; Winnicka, A. Fas/FasL pathway participates in resolution of mucosal inflammatory response early during HSV-2 infection. Immunobiology 2014, 219, 64–77. [Google Scholar] [CrossRef] [PubMed]


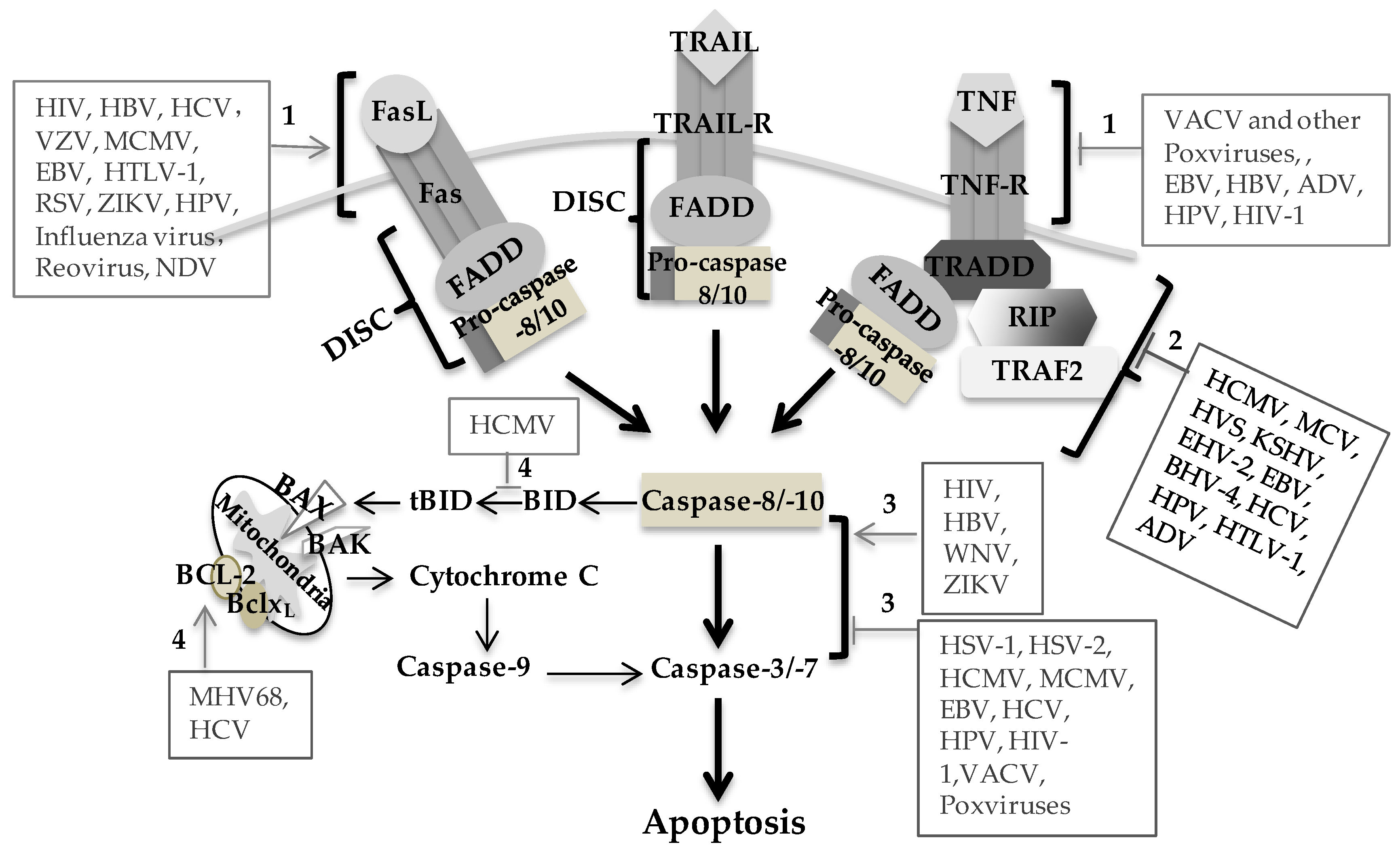{kind=link}
| Virus | Viral Activator | Mediated Signaling | References |
| HIV-1 | Tat | Fas, TRAIL-R | [27,28,29] |
| Env | Fas | [30] | |
| Vpu | Fas | [31] | |
| gp120 | Fas, TNF-R | [28,32,33] | |
| gp160 | Fas | [34] | |
| HBV | HBX | TRAIL-R | [35,36] |
| HCV | Core protein | Fas, TNF-R, TRAIL-R | [37,38,39,40] |
| EBV | LMP2A | Fas | [41] |
| LMP1 | Fas | [42] | |
| HPV | E2 | Fas, TNF-R | [43] |
| E7 | TNF-R | [44] | |
| HTLV-1 | Tax | TRAIL-R | [45] |
| Lyssavirus | Matrix protein | TRAIL-R | [46] |
| Virus | Viral Inhibitor | Mediated Signaling | References |
| Fibroma virus | TNFR2 ortholog | TNF-R | [47,48] |
| Vaccinia virus | CrmE | TNF-R | [49] |
| SPI-1 | Fas | [50] | |
| SPI-2 | Fas, TNF-R | [50,51,52,53,54] | |
| Myxoma virus | T2 | TNF-R | [55] |
| Cowpox virus | CrmA | Fas, TNF-R | [56] |
| HSV-1 | gD | Fas– | [57] |
| Ribonucleotide reductase R1 | Fas | [58] | |
| HSV-2 | Ribonucleotide reductase R1 | Fas | [58] |
| HCMV | IE2 | Fas, TRAIL-R | [59] |
| vMIA | Fas | [60] | |
| UL36 | Fas | [61] | |
| MCMV | M36 | Fas | [62] |
| M45 | TNF-R | [63] | |
| KSHV | v-FLIP | Fas, TRAIL-R | [64,65] |
| MCV | MC159 | Fas, TNF-R, TRAIL-R | [65,66,67] |
| EHV2 | E8 | Fas, TNF-R | [66] |
| HVS | v-FLIP | Fas, TRAIL-R | [65] |
| BHV-4 | v-FLIP | Fas, TRAIL-R | [65] |
| EBV | EBER | Fas | [68,69] |
| BHRF1 | Fas, TNF-R, TRAIL-R | [70,69] | |
| BZLF1 | TNF-R | [71] | |
| LMP1 | Fas, TRAIL-R | [72,73] | |
| MHV68 | M11 | Fas, TNF-R | [74] |
| HCV | Core protein | Fas, TNF-R | [75,76,77] |
| E2 | Fas | [78] | |
| NS5A | TNF-R | [79] | |
| HBV | Core protein | Fas | [80] |
| ADV | E3-10.4K/14.5K complex | Fas | [81] |
| E3-6.7K/10.4K/14.5K complex | TRAIL-R | [82] | |
| E3-RID complex | Fas, TNF-R, TRAIL-R | [83] | |
| E3-14.7K | Fas, TNF-R | [84] | |
| HPV | E5 | Fas, TRAIL-R | [85,86] |
| E6 | TNF-R, TRAIL-R | [87,88,89,90] | |
| E7 | Fas, TNF-R | [91,92] | |
| EBOV | Glycoprotein | Fas | [93] |
| MARV | Glycoprotein | Fas | [93] |
| HIV-1 | Nef | Fas | [94] |
| Tat | TRAIL-R | [95] | |
| HTLV-1 | Tax | Fas | [96,97] |
| HTLV-2 | Tax | Fas | [98] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Jiang, W.; Liu, Z.; Liu, S.; Liang, X. Virus Infection and Death Receptor-Mediated Apoptosis. Viruses 2017, 9, 316. https://doi.org/10.3390/v9110316
Zhou X, Jiang W, Liu Z, Liu S, Liang X. Virus Infection and Death Receptor-Mediated Apoptosis. Viruses. 2017; 9(11):316. https://doi.org/10.3390/v9110316
Chicago/Turabian StyleZhou, Xingchen, Wenbo Jiang, Zhongshun Liu, Shuai Liu, and Xiaozhen Liang. 2017. "Virus Infection and Death Receptor-Mediated Apoptosis" Viruses 9, no. 11: 316. https://doi.org/10.3390/v9110316
APA StyleZhou, X., Jiang, W., Liu, Z., Liu, S., & Liang, X. (2017). Virus Infection and Death Receptor-Mediated Apoptosis. Viruses, 9(11), 316. https://doi.org/10.3390/v9110316