Small Ruminant Lentiviruses: Genetic Variability, Tropism and Diagnosis

 ,
,  , and
, and
Abstract
:1. Introduction
2. Viral Genetic Variability Sources
2.1. Mutation
2.2. Recombination
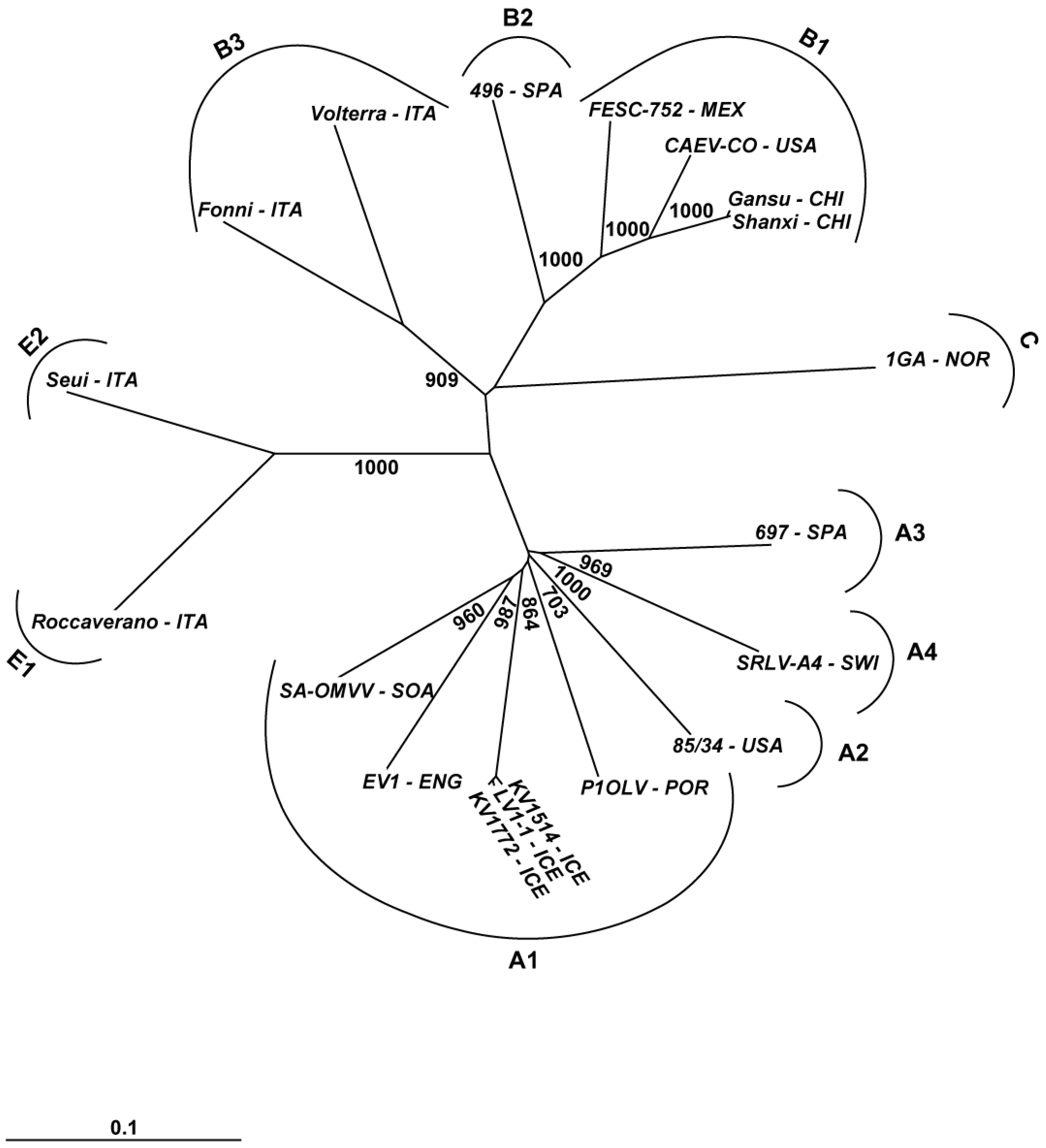
3. Phylogeny

{kind=link}
| Country | Genotype A | Genotype B | Genotype C | Genotype D | Genotype E | References | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | A2 | A3 | A4 | A5 | A6 | A7 | A8 | A9 | A10 | A11 | A12 | A13 | A14 | A15 | B1 | B2 | B3 | C | D | E1 | E2 | ||
| Brazil | G/S | G | [74,75,76] | ||||||||||||||||||||
| Canada | S | G | [77] | ||||||||||||||||||||
| China | G | [63,64] | |||||||||||||||||||||
| England | S | [70] | |||||||||||||||||||||
| Finland | S | S | [78] | ||||||||||||||||||||
| France | S | G/S | G/S | G/S | [27,79,80,81,91,92] | ||||||||||||||||||
| Greece | S | [82] | |||||||||||||||||||||
| Iceland | S | [67,68,69] | |||||||||||||||||||||
| Ireland | G | [83] | |||||||||||||||||||||
| Italy | G | G | G/S | G | G/S | G/S | S | G/S | G | G | [30,45,46,89,93,94] | ||||||||||||
| Japan | G | [84] | |||||||||||||||||||||
| Mexico | G/S | [25] | |||||||||||||||||||||
| Netherlands | S | [85] | |||||||||||||||||||||
| Norway | S | G/S | [61,62] | ||||||||||||||||||||
| Poland | G/S | G/S | G/S | G | S | [86,87] | |||||||||||||||||
| Portugal | S | [71] | |||||||||||||||||||||
| Russia | G | [88] | |||||||||||||||||||||
| Slovenia | S | G | S | G | [89] | ||||||||||||||||||
| South Africa | S | [66] | |||||||||||||||||||||
| South Korea | G | [90] | |||||||||||||||||||||
| Spain | S | S | S | G/S | [65,73] | ||||||||||||||||||
| Switzerland | S | G/S | G/S | G | G/S | G | G | S | S | [27,95] | |||||||||||||
| Turkey | S | S | S | S | S | S | [29,30] | ||||||||||||||||
| USA | S | G/S | [59,60,72,96] | ||||||||||||||||||||
4. Tropism
4.1. Host Species Restriction
4.2. Viral Genetics by Organ and Tissue
4.3 Cellular Tropism
5. In Vitro Diagnosis
5.1. ELISA Tests
5.2. Polymerase Chain Reaction (PCR)
| PCR type | DNA or cDNA source | Primers location into the viral genome | Sensitivity test | References |
|---|---|---|---|---|
| cPCR | Cell culture | gag and pol | pol-PCR was more sensitive than gag-PCR | [210] |
| snPCR | PBL | pol and LTR | LTR-PCR was more sensitive than pol-PCR | [212] |
| nPCR | PBMC | gag and pol | gag-PCR was more sensitive than pol-PCR | [213] |
| snPCR | PBMC | gag | gag-PCR was less sensitive than AGID | [214] |
| snPCR | Cell culture and PBMC | pol | High sensitivity, by using degenerate primers | [215] |
| nPCR | PBMC | gag | gag-PCR was more sensitive than AGID in seronegative animals | [216] |
| nPCR | PBL and blood | gag and LTR | gag-PCR was more sensitive than LTR-PCR | [220] |
| cPCR | Semen | env and pol | env-PCR was more sensitive than gag-PCR | [222] |
| cPCR | PBMC, milk cells and tissues | LTR | LTR-PCR had a sensitivity of 98% with regard to AGID and ELISA | [223] |
| cPCR | Cell culture | LTR, gag and env | LTR-PCR was more sensitive than gag-PCR and env-PCR | [226] |
| cPCR | PBMC | gag | gag-PCR was less sensitive than AGID | [228] |
| cPCR | Cell culture and PBMC | gag | gag-PCR was more sensitive than the ELISA and WB | [229] |
| nPCR | PBL | gag | gag-PCR increases its sensitivity when used along with hybridization | [230] |
6. Concluding Remarks
Acknowledgments
Conflict of Interest
References
- Christodoulopoulos, G. Maedi-Visna: Clinical review and short reference on the disease status in Mediterranean countries. Small Rum. Res. 2006, 62, 47–53. [Google Scholar] [CrossRef]
- Peterhans, E.; Greenland, T.; Badiola, J.; Harkiss, G.; Bertoni, G.; Amorena, B.; Eliaszewicz, M.; Juste, R.A.; Krassnig, R.; Lafont, J.P.; et al. Routes of transmission and consequences of small ruminant lentiviruses (SRLVs) infection and eradication schemes. Vet. Res. 2004, 35, 257–274. [Google Scholar] [CrossRef]
- Anderson, B.C.; Bulgin, M.S.; Adams, S.; Duelke, B. Firm udder in periparturient ewes with lymphocytic accumulations, retrovirus infection, and milk unavailable at the teat. J. Am. Vet. Med. Assoc. 1985, 186, 391–393. [Google Scholar]
- Keen, J.E.; Hungerford, L.L.; Littledike, E.T.; Wittum, T.E.; Kwang, J. Effect of ewe ovine lentivirus infection on ewe and lamb productivity. Prev. Vet. Med. 1997, 30, 155–169. [Google Scholar] [CrossRef]
- Pekelder, J.J.; Veenink, G.J.; Akkermans, J.P.; van Eldik, P.; Elving, L.; Houwers, D.J. Ovine lentivirus induced indurative lymphocytic mastitis and its effect on the growth of lambs. Vet. Rec. 1994, 134, 348–350. [Google Scholar]
- Dungu, B.; Vorster, J.; Bath, G.F.; Verwoerd, D.W. The effect of a natural maedi-visna virus infection on the productivity of South African sheep. Onderstepoort. J. Vet. Res. 2000, 67, 87–96. [Google Scholar]
- Legrottaglie, R.; Martini, M.; Barsotti, G.; Agrimi, P. The effects of ovine lentivirus infection on some productive aspects in a Sardinian sheep flock from Italy. Vet. Res. Commun. 1999, 23, 123–131. [Google Scholar] [CrossRef]
- Snowder, G.D.; Gates, N.L.; Glimp, H.A.; Gorham, J.R. Prevalence and effect of subclinical ovine progressive pneumonia virus infection on ewe wool and lamb production. J. Am. Vet. Med. Assoc. 1990, 197, 475–479. [Google Scholar]
- Arsenault, J.; Dubreuil, P.; Girard, C.; Simard, C.; Belanger, D. Maedi-visna impact on productivity in Quebec sheep flocks (Canada). Prev. Vet. Med. 2003, 59, 125–137. [Google Scholar] [CrossRef]
- Leitner, G.; Krifucks, O.; Weisblit, L.; Lavi, Y.; Bernstein, S.; Merin, U. The effect of caprine arthritis encephalitis virus infection on production in goats. Vet. J. 2010, 183, 328–331. [Google Scholar] [CrossRef]
- Nord, K.; Adnoy, T. Effects of infection by caprine arthritis-encephalitis virus on milk production of goats. J. Dairy Sci. 1997, 80, 2391–2397. [Google Scholar] [CrossRef]
- Turin, L.; Pisoni, G.; Giannino, M.L.; Antonini, M.; Rosati, S.; Ruffo, G.; Moroni, P. Correlation between milk parameters in CAEV seropositive and negative primiparous goats during an eradication program in Italian farm. Small Rum. Res. 2005, 57, 73–79. [Google Scholar] [CrossRef]
- Martínez, N.B.; Peris, R.C.; Roche, J.M.L.; Caballero, G.C. Efectos del virus de la artritis-encefalitis caprina sobre la producción y composición de la leche en cabras murciano-granadinas. PR: Pequeños Rumiantes 2002, 3, 26–30. [Google Scholar]
- Ryan, D.P.; Greenwood, P.L.; Nicholls, P.J. Effect of caprine arthritis-encephalitis virus infection on milk cell count and N-acetyl-beta-glucosaminidase activity in dairy goats. J. Dairy Res. 1993, 60, 299–306. [Google Scholar] [CrossRef]
- Martinez-Navalon, B.; Peris, C.; Gomez, E.A.; Peris, B.; Roche, M.L.; Caballero, C.; Goyena, E.; Berriatua, E. Quantitative estimation of the impact of caprine arthritis encephalitis virus infection on milk production by dairy goats. Vet. J. 2013. [Google Scholar] [CrossRef]
- Christodoulopoulos, G. Milk Production and Milk Fat Content in Commercial Karagouniko Breed Flocks Infected by Maedi Disease in Greece. In Proceedings of the Sixth International Sheep Veterinary Congress, Hersonissos Crete, Greece, 17–21 June 2005; pp. 344–345.
- Leitner, G.; Silanikove, N.; Merin, U. Estimate of milk and curd yield loss of sheep and goats with intrammamary infection and its relation to somatic cell count. Small Rum. Res. 2008, 74, 221–225. [Google Scholar] [CrossRef]
- Broughton-Neiswanger, L.E.; White, S.N.; Knowles, D.P.; Mousel, M.R.; Lewis, G.S.; Herndon, D.R.; Herrmann-Hoesing, L.M. Non-maternal transmission is the major mode of ovine lentivirus transmission in a ewe flock: A molecular epidemiology study. Infect. Genet. Evol. 2010, 10, 998–1007. [Google Scholar] [CrossRef]
- Leginagoikoa, I.; Daltabuit-Test, M.; Alvarez, V.; Arranz, J.; Juste, R.A.; Amorena, B.; de Andres, D.; Lujan, L.L.; Badiola, J.J.; Berriatua, E. Horizontal Maedi-Visna virus (MVV) infection in adult dairy-sheep raised under varying MVV-infection pressures investigated by ELISA and PCR. Res. Vet. Sci. 2006, 80, 235–241. [Google Scholar] [CrossRef]
- Leginagoikoa, I.; Minguijon, E.; Juste, R.A.; Barandika, J.; Amorena, B.; de Andres, D.; Badiola, J.J.; Lujan, L.; Berriatua, E. Effects of housing on the incidence of visna/maedi virus infection in sheep flocks. Res. Vet. Sci. 2010, 88, 415–421. [Google Scholar] [CrossRef]
- Alvarez, V.; Arranz, J.; Daltabuit-Test, M.; Leginagoikoa, I.; Juste, R.A.; Amorena, B.; de Andres, D.; Lujan, L.L.; Badiola, J.J.; Berriatua, E. Relative contribution of colostrum from Maedi-Visna virus (MVV) infected ewes to MVV-seroprevalence in lambs. Res. Vet. Sci. 2005, 78, 237–243. [Google Scholar] [CrossRef]
- Leginagoikoa, I.; Juste, R.A.; Barandika, J.; Amorena, B.; De Andres, D.; Lujan, L.; Badiola, J.; Berriatua, E. Extensive rearing hinders Maedi-Visna Virus (MVV) infection in sheep. Vet. Res. 2006, 37, 767–778. [Google Scholar] [CrossRef]
- Sigurdsson, B.; Grimsson, H.; Palsson, P.A. Maedi, a chronic, progressive infection of sheep’s lungs. J. Infect. Dis. 1952, 90, 233–241. [Google Scholar] [CrossRef]
- Blacklaws, B.A.; Berriatua, E.; Torsteinsdottir, S.; Watt, N.J.; de Andres, D.; Klein, D.; Harkiss, G.D. Transmission of small ruminant lentiviruses. Vet. Microbiol. 2004, 101, 199–208. [Google Scholar] [CrossRef]
- Ramirez, H.; Glaria, I.; de Andres, X.; Martinez, H.A.; Hernandez, M.M.; Reina, R.; Iraizoz, E.; Crespo, H.; Berriatua, E.; Vazquez, J.; et al. Recombinant small ruminant lentivirus subtype B1 in goats and sheep of imported breeds in Mexico. Vet. J. 2011, 190, 169–172. [Google Scholar] [CrossRef]
- Straub, O.C. Maedi-Visna virus infection in sheep. History and present knowledge. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 1–5. [Google Scholar] [CrossRef]
- Shah, C.; Boni, J.; Huder, J.B.; Vogt, H.R.; Muhlherr, J.; Zanoni, R.; Miserez, R.; Lutz, H.; Schupbach, J. Phylogenetic analysis and reclassification of caprine and ovine lentiviruses based on 104 new isolates: Evidence for regular sheep-to-goat transmission and worldwide propagation through livestock trade. Virology 2004, 319, 12–26. [Google Scholar] [CrossRef]
- Gifford, R.J. Viral evolution in deep time: Lentiviruses and mammals. Trends Genet. 2012, 28, 89–100. [Google Scholar] [CrossRef]
- Muz, D.; Oguzoglu, T.C.; Rosati, S.; Reina, R.; Bertolotti, L.; Burgu, I. First molecular characterization of visna/maedi viruses from naturally infected sheep in Turkey. Arch. Virol. 2012. [Google Scholar] [CrossRef]
- Bertolotti, L.; Mazzei, M.; Puggioni, G.; Carrozza, M.L.; Dei Giudici, S.; Muz, D.; Juganaru, M.; Patta, C.; Tolari, F.; Rosati, S. Characterization of new small ruminant lentivirus subtype B3 suggests animal trade within the Mediterranean Basin. J. Gen. Virol. 2011, 92, 1923–1929. [Google Scholar] [CrossRef]
- Foley, B.T. An Overview of the Molecular Phylogeny of Lentiviruses. In HIV Sequence Compedium; Kuiken, C., Foley, B., Freed, E., Hahn, B., Korber, B., Marx, P.A., McCutchan, F., Mellors, J.W., Mullins, J.I., Sodroski, J., Wolinksy, S., Eds.; Theoretical Biology and Biophysics Group, Los Alamos National Laboratory: Los Alamos, NM, USA, 2000; pp. 35–43. [Google Scholar]
- Eigen, M. Selforganization of matter and the evolution of biological macromolecules. Naturwissenschaften 1971, 58, 465–523. [Google Scholar] [CrossRef]
- Ojosnegros, S.; Perales, C.; Mas, A.; Domingo, E. Quasispecies as a matter of fact: Viruses and beyond. Virus Res. 2011, 162, 203–215. [Google Scholar] [CrossRef]
- Korber, B.; Gaschen, B.; Yusim, K.; Thakallapally, R.; Kesmir, C.; Detours, V. Evolutionary and immunological implications of contemporary HIV-1 variation. Br. Med. Bull. 2001, 58, 19–42. [Google Scholar] [CrossRef]
- Pasick, J. Maedi-visna virus and caprine arthritis-encephalitis virus: Distinct species or quasispecies and its implications for laboratory diagnosis. Can. J. Vet. Res. 1998, 62, 241–244. [Google Scholar]
- Smyth, R.P.; Davenport, M.P.; Mak, J. The origin of genetic diversity in HIV-1. Virus Res. 2012, 169, 415–429. [Google Scholar] [CrossRef]
- Roberts, J.D.; Bebenek, K.; Kunkel, T.A. The accuracy of reverse transcriptase from HIV-1. Science 1988, 242, 1171–1173. [Google Scholar]
- Vartanian, J.P.; Meyerhans, A.; Asjo, B.; Wain-Hobson, S. Selection, recombination, and G----A hypermutation of human immunodeficiency virus type 1 genomes. J. Virol. 1991, 65, 1779–1788. [Google Scholar]
- Bishop, K.N.; Holmes, R.K.; Sheehy, A.M.; Davidson, N.O.; Cho, S.J.; Malim, M.H. Cytidine deamination of retroviral DNA by diverse APOBEC proteins. Curr. Biol. 2004, 14, 1392–1396. [Google Scholar] [CrossRef]
- Dang, Y.; Wang, X.; Esselman, W.J.; Zheng, Y.H. Identification of APOBEC3DE as another antiretroviral factor from the human APOBEC family. J. Virol. 2006, 80, 10522–10533. [Google Scholar] [CrossRef]
- Terai, C.; Carson, D.A. Pyrimidine nucleotide and nucleic acid synthesis in human monocytes and macrophages. Exp. Cell. Res. 1991, 193, 375–381. [Google Scholar] [CrossRef]
- Turelli, P.; Guiguen, F.; Mornex, J.F.; Vigne, R.; Querat, G. dUTPase-minus caprine arthritis-encephalitis virus is attenuated for pathogenesis and accumulates G-to-A substitutions. J. Virol. 1997, 71, 4522–4530. [Google Scholar]
- Wain-Hobson, S.; Sonigo, P.; Guyader, M.; Gazit, A.; Henry, M. Erratic G→A hypermutation within a complete caprine arthritis-encephalitis virus (CAEV) provirus. Virology 1995, 209, 297–303. [Google Scholar] [CrossRef]
- Juganaru, M.; Reina, R.; Bertolotti, L.; Stella, M.C.; Profiti, M.; Armentano, M.; Bollo, E.; Amorena, B.; Rosati, S. In vitro properties of small ruminant lentivirus genotype E. Virology 2011, 410, 88–95. [Google Scholar] [CrossRef]
- Reina, R.; Grego, E.; Bertolotti, L.; De Meneghi, D.; Rosati, S. Genome analysis of small-ruminant lentivirus genotype E: A caprine lentivirus with natural deletions of the dUTPase subunit, vpr-like accessory gene, and 70-base-pair repeat of the U3 region. J. Virol. 2009, 83, 1152–1155. [Google Scholar] [CrossRef]
- Reina, R.; Bertolotti, L.; Dei Giudici, S.; Puggioni, G.; Ponti, N.; Profiti, M.; Patta, C.; Rosati, S. Small ruminant lentivirus genotype E is widespread in Sarda goat. Vet. Microbiol. 2010, 144, 24–31. [Google Scholar] [CrossRef]
- Tormo-Mas, M.A.; Mir, I.; Shrestha, A.; Tallent, S.M.; Campoy, S.; Lasa, I.; Barbe, J.; Novick, R.P.; Christie, G.E.; Penades, J.R. Moonlighting bacteriophage proteins derepress staphylococcal pathogenicity islands. Nature 2012, 465, 779–782. [Google Scholar]
- Sheehy, A.M.; Gaddis, N.C.; Choi, J.D.; Malim, M.H. Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature 2002, 418, 646–650. [Google Scholar] [CrossRef]
- Andresdottir, V.; Skraban, R.; Matthiasdottir, S.; Lutley, R.; Agnarsdottir, G.; Thorsteinsdottir, H. Selection of antigenic variants in maedi-visna virus infection. J. Gen. Virol. 2002, 83, 2543–2551. [Google Scholar]
- Andresdottir, V. Evidence for recombination in the envelope gene of maedi-visna virus. Virus Genes 2003, 27, 5–9. [Google Scholar] [CrossRef]
- Jetzt, A.E.; Yu, H.; Klarmann, G.J.; Ron, Y.; Preston, B.D.; Dougherty, J.P. High rate of recombination throughout the human immunodeficiency virus type 1 genome. J. Virol. 2000, 74, 1234–1240. [Google Scholar]
- Jolly, P.E.; Narayan, O. Evidence for interference, coinfections, and intertypic virus enhancement of infection by ovine-caprine lentiviruses. J. Virol. 1989, 63, 4682–4688. [Google Scholar]
- Hu, W.S.; Temin, H.M. Retroviral recombination and reverse transcription. Science 1990, 250, 1227–1233. [Google Scholar]
- Hu, W.S.; Temin, H.M. Genetic consequences of packaging two RNA genomes in one retroviral particle: Pseudodiploidy and high rate of genetic recombination. Proc. Natl. Acad. Sci. USA 1990, 87, 1556–1560. [Google Scholar] [CrossRef]
- Pisoni, G.; Bertoni, G.; Puricelli, M.; Maccalli, M.; Moroni, P. Demonstration of coinfection with and recombination by caprine arthritis-encephalitis virus and maedi-visna virus in naturally infected goats. J. Virol. 2007, 81, 4948–4955. [Google Scholar] [CrossRef]
- Zanoni, R.G. Phylogenetic analysis of small ruminant lentiviruses. J. Gen. Virol. 1998, 79, 1951–1961. [Google Scholar]
- Peeters, M.; Esu-Williams, E.; Vergne, L.; Montavon, C.; Mulanga-Kabeya, C.; Harry, T.; Ibironke, A.; Lesage, D.; Patrel, D.; Delaporte, E. Predominance of subtype A and G HIV type 1 in Nigeria, with geographical differences in their distribution. AIDS Res. Hum. Retroviruses 2000, 16, 315–325. [Google Scholar] [CrossRef]
- Adams, D.S.; Oliver, R.E.; Ameghino, E.; DeMartini, J.C.; Verwoerd, D.W.; Houwers, D.J.; Waghela, S.; Gorham, J.R.; Hyllseth, B.; Dawson, M.; et al. Global survey of serological evidence of caprine arthritis-encephalitis virus infection. Vet. Rec. 1984, 115, 493–495. [Google Scholar]
- Narayan, O.; Clements, J.E.; Strandberg, J.D.; Cork, L.C.; Griffin, D.E. Biological characterization of the virus causing leukoencephalitis and arthritis in goats. J. Gen. Virol. 1980, 50, 69–79. [Google Scholar] [CrossRef]
- Saltarelli, M.; Querat, G.; Konings, D.A.; Vigne, R.; Clements, J.E. Nucleotide sequence and transcriptional analysis of molecular clones of CAEV which generate infectious virus. Virology 1990, 179, 347–364. [Google Scholar] [CrossRef]
- Gjerset, B.; Jonassen, C.M.; Rimstad, E. Natural transmission and comparative analysis of small ruminant lentiviruses in the Norwegian sheep and goat populations. Virus Res. 2007, 125, 153–161. [Google Scholar] [CrossRef]
- Gjerset, B.; Storset, A.K.; Rimstad, E. Genetic diversity of small-ruminant lentiviruses: Characterization of Norwegian isolates of Caprine arthritis encephalitis virus. J. Gen. Virol. 2006, 87, 573–580. [Google Scholar] [CrossRef]
- Qu, J.; Guo, W.; Zhao, L.; Sheng, R.; Xiang, W. The gene cloning and sequence analysis of the whole genome of Caprine Arthritis Encephalitis virus (CAEV) GANSU strain. Chin. J. Virol. 2005, 21, 389–392. [Google Scholar]
- Huang, J.; Sun, Y.; Liu, Y.; Xiao, H.; Zhuang, S. Development of a loop-mediated isothermal amplification method for rapid detection of caprine arthritis-encephalitis virus proviral DNA. Arch. Virol. 2012, 157, 1463–1469. [Google Scholar] [CrossRef]
- Glaria, I.; Reina, R.; Crespo, H.; de Andres, X.; Ramirez, H.; Biescas, E.; Perez, M.M.; Badiola, J.; Lujan, L.; Amorena, B.; et al. Phylogenetic analysis of SRLV sequences from an arthritic sheep outbreak demonstrates the introduction of CAEV-like viruses among Spanish sheep. Vet. Microbiol. 2009, 138, 156–162. [Google Scholar] [CrossRef]
- Querat, G.; Audoly, G.; Sonigo, P.; Vigne, R. Nucleotide sequence analysis of SA-OMVV, a visna-related ovine lentivirus: Phylogenetic history of lentiviruses. Virology 1990, 175, 434–447. [Google Scholar] [CrossRef]
- Andresson, O.S.; Elser, J.E.; Tobin, G.J.; Greenwood, J.D.; Gonda, M.A.; Georgsson, G.; Andresdottir, V.; Benediktsdottir, E.; Carlsdottir, H.M.; Mantyla, E.O. Nucleotide sequence and biological properties of a pathogenic proviral molecular clone of neurovirulent visna virus. Virology 1993, 193, 89–105. [Google Scholar] [CrossRef]
- Sonigo, P.; Alizon, M.; Staskus, K.; Klatzmann, D.; Cole, S.; Danos, O.; Retzel, E.; Tiollais, P.; Haase, A.; Wain-Hobson, S. Nucleotide sequence of the visna lentivirus: Relationship to the AIDS virus. Cell 1985, 42, 369–382. [Google Scholar] [CrossRef]
- Staskus, K.A.; Retzel, E.F.; Lewis, E.D.; Silsby, J.L.; St Cyr, S.; Rank, J.M.; Wietgrefe, S.W.; Haase, A.T.; Cook, R.; Fast, D.; et al. Isolation of replication-competent molecular clones of visna virus. Virology 1991, 181, 228–240. [Google Scholar] [CrossRef]
- Sargan, D.R.; Bennet, I.D.; Cousens, C.; Roy, D.J.; Blacklaws, B.A.; Dalziel, R.G.; Watt, N.J.; McConnell, I. Nucleotide sequence of EV1, a British isolate of maedi-visna virus. J. Gen. Virol. 1991, 72, 1893–1903. [Google Scholar] [CrossRef]
- Fevereiro, M.T.; Barros, S.S. Caracterização biológica e molecular de um lentivírus de ovino isolado em Portugal. RPCV 2004, 99, 27–39. [Google Scholar]
- Karr, B.M.; Chebloune, Y.; Leung, K.; Narayan, O. Genetic characterization of two phenotypically distinct North American ovine lentiviruses and their possible origin from caprine arthritis-encephalitis virus. Virology 1996, 225, 1–10. [Google Scholar] [CrossRef]
- Glaria, I.; Reina, R.; Ramirez, H.; de Andres, X.; Crespo, H.; Jauregui, P.; Salazar, E.; Lujan, L.; Perez, M.M.; Benavides, J.; et al. Visna/Maedi virus genetic characterization and serological diagnosis of infection in sheep from a neurological outbreak. Vet. Microbiol. 2012, 155, 137–146. [Google Scholar] [CrossRef]
- Castro, R.S.; Greenland, T.; Leite, R.C.; Gouveia, A.; Mornex, J.F.; Cordier, G. Conserved sequence motifs involving the tat reading frame of Brazilian caprine lentiviruses indicate affiliations to both caprine arthritis-encephalitis virus and visna-maedi virus. J. Gen. Virol. 1999, 80, 1583–1589. [Google Scholar]
- Feitosaa, L.A.L.V.; de Silva Teixeira, M.F.; Rizaldo, P.R.; Silva da Cunhac, R.M.; Santos Limad, J.P.M.; Andriolib, A.; Medeiros, D.T.V.; Pessoa de Meloa, V.S.; Nunes Pinheiroa, D.C.S. Phylogenetic analysis of small ruminant lentiviruses from Northern Brazil. Small Rum. Res. 2010, 94, 205–209. [Google Scholar] [CrossRef]
- Ravazzolo, A.P.; Reischak, D.; Peterhans, E.; Zanoni, R. Phylogenetic analysis of small ruminant lentiviruses from Southern Brazil. Virus Res. 2001, 79, 117–123. [Google Scholar] [CrossRef]
- L'Homme, Y.; Ouardani, M.; Levesque, V.; Bertoni, G.; Simard, C.; Pisoni, G. Molecular characterization and phylogenetic analysis of small ruminant lentiviruses isolated from Canadian sheep and goats. Virol. J. 2011, 8. [Google Scholar] [CrossRef]
- Laamanen, I.; Jakava-Viljanen, M.; Sihvonen, L. Genetic characterization of maedi-visna virus (MVV) detected in Finland. Vet. Microbiol. 2007, 122, 357–365. [Google Scholar] [CrossRef]
- Leroux, C.; Chastang, J.; Greenland, T.; Mornex, J.F. Genomic heterogeneity of small ruminant lentiviruses: Existence of heterogeneous populations in sheep and of the same lentiviral genotypes in sheep and goats. Arch. Virol. 1997, 142, 1125–1137. [Google Scholar] [CrossRef]
- Germain, K.; Valas, S. Distribution and heterogeneity of small ruminant lentivirus envelope subtypes in naturally infected French sheep. Virus Res. 2006, 120, 156–162. [Google Scholar] [CrossRef]
- Valas, S.; Benoit, C.; Guionaud, C.; Perrin, G.; Mamoun, R.Z. North American and French caprine arthritis-encephalitis viruses emerge from ovine maedi-visna viruses. Virology 1997, 237, 307–318. [Google Scholar] [CrossRef]
- Angelopoulou, K.; Karanikolaou, K.; Papanastasopoulou, M.; Koumpati-Artopiou, M.; Vlemmas, I.; Papadopoulos, O.; Koptopoulos, G. First partial characterisation of small ruminant lentiviruses from Greece. Vet. Microbiol. 2005, 109, 1–9. [Google Scholar] [CrossRef]
- Rolland, M.; Mooney, J.; Valas, S.; Perrin, G.; Mamoun, R.Z. Characterisation of an Irish caprine lentivirus strain—SRLV phylogeny revisited. Virus Res. 2002, 85, 29–39. [Google Scholar] [CrossRef]
- Konishi, M.; Tsuduku, S.; Haritani, M.; Murakami, K.; Tsuboi, T.; Kobayashi, C.; Yoshikawa, K.; Kimura, K.M.; Sentsui, H. An epidemic of caprine arthritis encephalitis in Japan: Isolation of the virus. J. Vet. Med. Sci. 2004, 66, 911–917. [Google Scholar] [CrossRef]
- Zanoni, R.G.; Nauta, I.M.; Pauli, U.; Peterhans, E. Expression in Escherichia coli and sequencing of the coding region for the capsid protein of Dutch maedi-visna virus strain ZZV 1050: Application of recombinant protein in enzyme-linked immunosorbent assay for the detection of caprine and ovine lentiviruses. J. Clin. Microbiol. 1991, 29, 1290–1294. [Google Scholar]
- Kuzmak, J.; Rola, M.; Gallay, K.; Chebloune, Y. Molecular characterization of lentiviruses from goats from Poland based on gag gene sequence analysis. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 211–223. [Google Scholar] [CrossRef]
- Olech, M.; Rachid, A.; Croise, B.; Kuzmak, J.; Valas, S. Genetic and antigenic characterization of small ruminant lentiviruses circulating in Poland. Virus Res. 2011, 163, 528–536. [Google Scholar]
- Baryshnikova, E.I.; Malogolovkin, A.S.; Kolbasova, O.L.; Tsybanov, S. Comparative characteristics of the biological properties of small ruminant lentiviruses. Vopr. Virusol. 2011, 56, 42–45. [Google Scholar]
- Kuhar, U.; Barlic-Maganja, D.; Grom, J. Phylogenetic analysis of small ruminant lentiviruses detected in Slovenia. Vet. Microbiol. 2012. [Google Scholar] [CrossRef]
- Park, J.E.; Son, S.Y.; Shin, H.J. Sequence comparison on gag gene of caprine arthritis encephalitis virus from Korea. Virus Genes 2010, 41, 99–101. [Google Scholar] [CrossRef]
- Leroux, C.; Vuillermoz, S.; Mornex, J.F.; Greenland, T. Genomic heterogeneity in the pol region of ovine lentiviruses obtained from bronchoalveolar cells of infected sheep from France. J. Gen. Virol. 1995, 76, 1533–1537. [Google Scholar] [CrossRef]
- Leroux, C.; Greenland, T.; Mornex, J.F. Molecular characterization of field isolates of lentiviruses of small ruminants. AIDS Res. Hum. Retroviruses 1996, 12, 427–429. [Google Scholar] [CrossRef]
- Pisoni, G.; Bertoni, G.; Manarolla, G.; Vogt, H.R.; Scaccabarozzi, L.; Locatelli, C.; Moroni, P. Genetic analysis of small ruminant lentiviruses following lactogenic transmission. Virology 2010, 407, 91–99. [Google Scholar] [CrossRef]
- Grego, E.; Bertolotti, L.; Quasso, A.; Profiti, M.; Lacerenza, D.; Muz, D.; Rosati, S. Genetic characterization of small ruminant lentivirus in Italian mixed flocks: Evidence for a novel genotype circulating in a local goat population. J. Gen. Virol. 2007, 88, 3423–3427. [Google Scholar] [CrossRef]
- Shah, C.; Huder, J.B.; Boni, J.; Schonmann, M.; Muhlherr, J.; Lutz, H.; Schupbach, J. Direct evidence for natural transmission of small-ruminant lentiviruses of subtype A4 from goats to sheep and vice versa. J. Virol. 2004, 78, 7518–7522. [Google Scholar] [CrossRef]
- Chebloune, Y.; Sheffer, D.; Karr, B.M.; Stephens, E.; Narayan, O. Restrictive type of replication of ovine/caprine lentiviruses in ovine fibroblast cell cultures. Virology 1996, 222, 21–30. [Google Scholar] [CrossRef]
- Banks, K.L.; Adams, D.S.; McGuire, T.C.; Carlson, J. Experimental infection of sheep by caprine arthritis-encephalitis virus and goats by progressive pneumonia virus. Am. J. Vet. Res. 1983, 44, 2307–2311. [Google Scholar]
- Dickson, J.; Ellis, T. Experimental caprine retrovirus infection in sheep. Vet. Rec. 1989, 125. [Google Scholar] [CrossRef]
- Oliver, R.; Cathcart, A.; McNiven, R.; Poole, W.; Robati, G. Transmission of caprine arthritis encephalitis virus to sheep. NZ Vet. J. 1984, 32, 199–200. [Google Scholar]
- Oliver, R.; Cathcart, A.; McNiven, R.; Poole, W.; Robati, G. Infection of lambs with caprine arthritis encephalitis virus by feeding milk from infected goats. Vet. Rec. 1985, 116, 83. [Google Scholar]
- Oliver, R.E.; McNiven, R.A.; Julian, A.F.; Poole, W.S. Experimental infection of sheep and goats with caprine arthritis-encephalitis virus. NZ Vet. J. 1982, 30, 158–159. [Google Scholar] [CrossRef]
- Smith, V.W.; Dickson, J.; Coackley, W.; Carman, H. Response of merino sheep to inoculation with a caprine retrovirus. Vet. Rec. 1985, 117, 61–63. [Google Scholar]
- Pisoni, G.; Quasso, A.; Moroni, P. Phylogenetic analysis of small-ruminant lentivirus subtype B1 in mixed flocks: Evidence for natural transmission from goats to sheep. Virology 2005, 339, 147–152. [Google Scholar] [CrossRef]
- Cheevers, W.P.; Knowles, D.P., Jr.; Norton, L.K. Neutralization-resistant antigenic variants of caprine arthritis-encephalitis lentivirus associated with progressive arthritis. J. Infect. Dis. 1991, 164, 679–685. [Google Scholar] [CrossRef]
- Hotzel, I.; Cheevers, W. Differential receptor usage of small ruminant lentiviruses in ovine and caprine cells: Host range but not cytopathic phenotype is determined by receptor usage. Virology 2002, 301, 21–31. [Google Scholar] [CrossRef]
- Bruett, L.; Clements, J.E. Functional murine leukemia virus vectors pseudotyped with the visna virus envelope show expanded visna virus cell tropism. J. Virol. 2001, 75, 11464–11473. [Google Scholar] [CrossRef]
- Hotzel, I.; Cheevers, W.P. Host range of small-ruminant lentivirus cytopathic variants determined with a selectable caprine arthritis-encephalitis virus pseudotype system. J. Virol. 2001, 75, 7384–7391. [Google Scholar] [CrossRef]
- Valas, S.; Benoit, C.; Baudry, C.; Perrin, G.; Mamoun, R.Z. Variability and immunogenicity of caprine arthritis-encephalitis virus surface glycoprotein. J. Virol. 2000, 74, 6178–6185. [Google Scholar] [CrossRef]
- Patel, J.R.; Heldens, J.G.; Bakonyi, T.; Rusvai, M. Important mammalian veterinary viral immunodiseases and their control. Vaccine 2012, 30, 1767–1781. [Google Scholar] [CrossRef]
- Bieniasz, P.D. An overview of intracellular interactions between immunodeficiency viruses and their hosts. AIDS 2012, 26, 1243–1254. [Google Scholar] [CrossRef]
- Jauregui, P.; Crespo, H.; Glaria, I.; Lujan, L.; Contreras, A.; Rosati, S.; de Andres, D.; Amorena, B.; Towers, G.J.; Reina, R. Ovine TRIM5alpha can restrict visna/maedi virus. J. Virol. 2012, 86, 9504–9509. [Google Scholar] [CrossRef]
- LaRue, R.S.; Lengyel, J.; Jonsson, S.R.; Andresdottir, V.; Harris, R.S. Lentiviral Vif degrades the APOBEC3Z3/APOBEC3H protein of its mammalian host and is capable of cross-species activity. J. Virol. 2010, 84, 8193–8201. [Google Scholar] [CrossRef]
- Korber, B.T.; Kunstman, K.J.; Patterson, B.K.; Furtado, M.; Mcevilly, M.M.; Levy, R.; Wolinsky, S.M. Genetic differences between blood- and brain-derived viral sequences from Human Immunodeficiency Virus Type-1 infected patients: Evidence of conserved elements in the V3 region of the Envelope protein of brain-derived sequences. J. Virol. 1994, 68, 7467–7481. [Google Scholar]
- Keys, B.; Karis, J.; Fadeel, B.; Valentin, A.; Norkrans, G.; Haqberg, L.; Chiodi, F. V3 sequences of paired HIV-1 isolates from blood and cerebrospinal fluid cluster according to host and show variation related to the clinical stage of disease. Virology 1993, 196, 475–483. [Google Scholar] [CrossRef]
- Liu, P.; Hudson, L.C.; Tompkins, M.B.; Vahlenkamp, T.W.; Meeker, R.B. Compartmentalization and evolution of feline immunodeficiency virus between the central nervous system and periphery following intracerebroventricular or systemic inoculation. J. Neurovirol. 2006, 12, 307–321. [Google Scholar]
- Pisoni, G.; Moroni, P.; Turin, L.; Bertoni, G. Compartmentalization of small ruminant lentivirus between blood and colostrum in infected goats. Virology 2007, 369, 119–130. [Google Scholar] [CrossRef]
- Ramirez, H.; Reina, R.; Bertolotti, L.; Cenoz, A.; Hernandez, M.M.; San Roman, B.; Glaria, I.; de Andres, X.; Crespo, H.; Jauregui, P.; et al. Study of compartmentalization in the visna clinical form of small ruminant lentivirus infection in sheep. BMC Vet. Res. 2012, 8, e8. [Google Scholar] [CrossRef]
- Chen, M.F.; Westmoreland, S.; Ryzhova, E.V.; Martín-García, J.; Soldan, S.S.; Lackner, A.; González-Scarano, F. Simian immunodeficiency virus envelope compartmentalizes in brain regions independent of neuropathology. J. Neurovirol. 2006, 12, 73–89. [Google Scholar] [CrossRef]
- Zárate, S.; Kosakovsky, P.S.L.; Shapshak, P.; Frost, S.D.W. Comparative study of methods for detecting sequence compartmentalization in human immunodeficiency virus type 1. J. Virol. 2007, 81, 6643–6651. [Google Scholar] [CrossRef]
- Abbate, I.; Cappiello, G.; Longo, R.; Ursitti, A.; Spano, A.; Calcaterra, S.; Dianzani, F.; Antinori, A.; Capobianchi, M.R. Cell membrane proteins and quasispecies compartmentalization of CSF and plasma HIV-1 from AIDS patients with neurological disorders. Infect. Genet. Evol. 2005, 5, 247–253. [Google Scholar] [CrossRef]
- Burkala, E.; He, J.; West, J.; Wood, C.; Petito, C. Compartmentalization of HIV-1 in the central nervous system: Role of the choroid plexus. AIDS 2005, 19, 675–684. [Google Scholar] [CrossRef]
- Philpott, S.; Burger, H.; Tsoukas, C.; Foley, B.; Anastos, K.; Kitchen, C.; Weiser, B. Human immunodeficiency virus type 1 genomic RNA sequences in the female genital tract and blood: Compartmentalization and intrapatient recombination. J. Virol. 2005, 79, 353–363. [Google Scholar]
- Saag, M.S.; Hahn, B.H.; Gibbons, J.; Li, Y. Extensive variation of human immunodeficiency virus type-1 in vivo. Nature 1988, 334, 440–444. [Google Scholar] [CrossRef]
- Shankarappa, R.; Margolick, J.B.; Gange, S.J.; Rodrigo, A.G.; Upchurch, D.; Farzadegan, H.; Gupta, P.; Rinaldo, C.R.; Learn, G.H.; He, X.; et al. Consistent viral evolutionary changes associated with the progression of human immunodeficiency virus type 1 infection. J. Virol. 1999, 73, 10489–10502. [Google Scholar]
- Si-Mohamed, A.; Kazatchkine, M.D.; Heard, I.; Goujon, C.; Prazuck, T.; Aymard, G.; Cessot, G.; Kuo, Y.H.; Bernard, M.C.; Diquet, B.; et al. Selection of drug-resistant variants in the female genital tract of human immunodeficiency virus type 1-infected women receiving antiretroviral therapy. J. Infect. Dis. 2000, 182, 112–122. [Google Scholar] [CrossRef]
- Smit, T.; Brew, B.; Tourtellotte, W.; Morgello, S.; Gelman, B.; Saksena., N. Independent evolution of human immunodeficiency virus (HIV) drug resistance mutations in diverse areas of the brain in HIV-infected patients, with and without dementia, on antiretroviral treatment. J. Virol. 2004, 78, 10133–10148. [Google Scholar] [CrossRef]
- Pepin, M.; Vitu, C.; Russo, P.; Mornex, J.F.; Peterhans, E. Maedi-visna virus infection in sheep: A review. Vet. Res. 1998, 29, 341–367. [Google Scholar]
- Barros, S.C.; Andresdottir, V.; Fevereiro, M. Cellular specificity and replication rate of Maedi Visna virus in vitro can be controlled by LTR sequences. Arch. Virol. 2005, 150, 201–213. [Google Scholar] [CrossRef]
- Oskarsson, T.; Hreggvidsdottir, H.S.; Agnarsdottir, G.; Matthiasdottir, S.; Ogmundsdottir, M.H.; Jonsson, S.R.; Georgsson, G.; Ingvarsson, S.; Andresson, O.S.; Andresdottir, V. Duplicated sequence motif in the long terminal repeat of maedi-visna virus extends cell tropism and is associated with neurovirulence. J. Virol. 2007, 81, 4052–4057. [Google Scholar] [CrossRef]
- Murphy, B.; McElliott, V.; Vapniarsky, N.; Oliver, A.; Rowe, J. Tissue tropism and promoter sequence variation in caprine arthritis encephalitis virus infected goats. Virus Res. 2010, 151, 177–184. [Google Scholar] [CrossRef]
- Bertoni, G.; Hertig, C.; Zahno, M.L.; Vogt, H.R.; Dufour, S.; Cordano, P.; Peterhans, E.; Cheevers, W.P.; Sonigo, P.; Pancino, G. B-cell epitopes of the envelope glycoprotein of caprine arthritis-encephalitis virus and antibody response in infected goats. J. Gen. Virol. 2000, 81, 2929–2940. [Google Scholar]
- Hotzel, I.; Cheevers, W.P. Sequence similarity between the envelope surface unit (SU) glycoproteins of primate and small ruminant lentiviruses. Virus Res. 2000, 69, 47–54. [Google Scholar] [CrossRef]
- Mwaengo, D.M.; Grant, R.F.; DeMartini, J.C.; Carlson, J.O. Envelope glycoprotein nucleotide sequence and genetic characterization of North American ovine lentiviruses. Virology 1997, 238, 135–144. [Google Scholar] [CrossRef]
- Blacklaws, B.; Bird, P.; McConnell, I. Early events in infection of lymphoid tissue by a lentivirus, maedi-visna. Trends Microbiol. 1995, 3, 434–440. [Google Scholar] [CrossRef]
- Leroux, C.; Mornex, J.F. Retroviral infections in sheep and the associated diseases. Small Rum. Res. 2008, 76, 68–76. [Google Scholar] [CrossRef]
- Palsson, P.A. Maedi and visna in sheep. Front. Biol. 1976, 44, 17–43. [Google Scholar]
- Torsteinsdottir, S.; Agnarsdottir, G.; Matthiasdottir, S.; Rafnar, B.; Andresdottir, V.; Andresson, O.S.; Staskus, K.; Petursson, G.; Palsson, P.A.; Georgsson, G. In vivo and in vitro infection with two different molecular clones of visna virus. Virology 1997, 229, 370–380. [Google Scholar] [CrossRef]
- Andresdottir, V.; Tang, X.; Agnarsdottir, G.; Andresson, O.S.; Georgsson, G.; Skraban, R.; Torsteinsdottir, S.; Rafnar, B.; Benediktsdottir, E.; Matthiasdottir, S.; et al. Biological and genetic differences between lung- and brain-derived isolates of maedi-visna virus. Virus Genes 1998, 16, 281–293. [Google Scholar] [CrossRef]
- Narayan, O.; Wolinsky, J.S.; Clements, J.E.; Strandberg, J.D.; Griffin, D.E.; Cork, L.C. Slow virus replication: The role of macrophages in the persistence and expression of visna viruses of sheep and goats. J. Gen. Virol. 1982, 59, 345–356. [Google Scholar] [CrossRef]
- Ryan, S.; Tiley, L.; McConnell, I.; Blacklaws, B. Infection of dendritic cells by the Maedi-Visna lentivirus. J. Virol. 2000, 74, 10096–10103. [Google Scholar] [CrossRef]
- Gendelman, H.E.; Narayan, O.; Kennedy-Stoskopf, S.; Kennedy, P.G.; Ghotbi, Z.; Clements, J.E.; Stanley, J.; Pezeshkpour, G. Tropism of sheep lentiviruses for monocytes: Susceptibility to infection and virus gene expression increase during maturation of monocytes to macrophages. J. Virol. 1986, 58, 67–74. [Google Scholar]
- Guiguen, F.; Asso, J. Inhibition of CAEV expression in goats experimentally and naturally infected. Dev. Biol. Stand. 1990, 72, 235–239. [Google Scholar]
- Narayan, O.; Kennedy-Stoskopf, S.; Sheffer, D.; Griffin, D.E.; Clements, J.E. Activation of caprine arthritis-encephalitis virus expression during maturation of monocytes to macrophages. Infect. Immun. 1983, 41, 67–73. [Google Scholar]
- Peluso, R.; Haase, A.T.; Stowring, L.; Edwards, M.; Ventura, P. A Trojan horse mechanism for the spread of visna virus in monocytes. Nucleic Acids Res. 1988, 16, 4116–4173. [Google Scholar]
- Barros, S.C.; Ramos, F.; Duarte, M.; Fagulha, T.; Cruz, B.; Fevereiro, M. Genomic characterization of a slow/low maedi visna virus. Virus Genes 2004, 29, 199–210. [Google Scholar] [CrossRef]
- Querat, G.; Barban, V.; Sauze, N.; Filippi, P.; Vigne, R.; Russo, P.; Vitu, C. Highly lytic and persistent lentiviruses naturally present in sheep with progressive pneumonia are genetically distinct. J. Virol. 1984, 52, 672–679. [Google Scholar]
- Singh, D.K.; Chebloune, Y.; Mselli-Lakhal, L.; Karr, B.M.; Narayan, O. Ovine lentivirus-infected macrophages mediate productive infection in cell types that are not susceptible to infection with cell-free virus. J. Gen. Virol. 1999, 80, 1437–1444. [Google Scholar]
- Sihvonen, L.; Veijalainen, P. Kinetics of maedi virus production in sheep choroid plexus cells. Vet. Microbiol. 1981, 6, 1–8. [Google Scholar] [CrossRef]
- Sigurdardottir, B.; Thormar, H. Isolation of a viral agent from the lungs of sheep affected with maedi. J. Infect. Dis. 1964, 114, 55–60. [Google Scholar] [CrossRef]
- Belov, L.; Whalley, J.M. Virus-specific polypeptides of caprine arthritis-encephalitis virus recognized by monoclonal antibodies to virion proteins p24 and p14. J. Gen. Virol. 1988, 69, 1097–1103. [Google Scholar] [CrossRef]
- Kirkland, P.D.; Batty, E.M. Caprine arthritis-encephalitis virus: An efficient method for the large scale production of serological antigens. J. Virol. Methods 1987, 16, 323–326. [Google Scholar] [CrossRef]
- Dahlberg, J.E.; Gaskin, J.M.; Perk, K. Morphological and immunological comparison of caprine arthritis encephalitis and ovine progressive pneumonia viruses. J. Virol. 1981, 39, 914–919. [Google Scholar]
- Lechat, E.; Milhau, N.; Brun, P.; Bellaton, C.; Greenland, T.; Mornex, J.F.; Le Jan, C. Goat endothelial cells may be infected in vitro by transmigration of caprine arthritis-encephalitis virus-infected leucocytes. Vet. Immunol. Immunopathol. 2005, 104, 257–263. [Google Scholar] [CrossRef]
- Milhau, N.; Bellaton, C.; Balleydier, S.; Gaonach, M.; Le Jan, C. In vitro infection of aortic endothelial cells by caprine arthritis encephalitis virus enhances in vitro transmigration of peripheral blood leukocytes and modulates their phenotypic expression. Vet. Res. 2003, 34, 273–284. [Google Scholar] [CrossRef]
- Mselli-Lakhal, L.; Guiguen, F.; Fornazero, C.; Favier, C.; Durand, J.; Grezel, D.; Moussa, A.; Mornex, J.F.; Chebloune, Y. Immortalized goat milk epithelial cell lines replicate CAEV at high level. Vet. Res. 2001, 32, 429–440. [Google Scholar] [CrossRef]
- Bolea, R.; Monleon, E.; Carrasco, L.; Vargas, A.; de Andres, D.; Amorena, B.; Badiola, J.J.; Lujan, L. Maedi-visna virus infection of ovine mammary epithelial cells. Vet. Res. 2006, 37, 133–144. [Google Scholar] [CrossRef]
- Lamara, A.; Fieni, F.; Mselli-Lakhal, L.; Tainturier, D.; Chebloune, Y. Efficient replication of caprine arthritis-encephalitis virus in goat granulosa cells. Virus Res. 2001, 79, 165–172. [Google Scholar] [CrossRef]
- Lamara, A.; Fieni, F.; Mselli-Lakhal, L.; Tainturier, D.; Chebloune, Y. Epithelial cells from goat oviduct are highly permissive for productive infection with caprine arthritis-encephalitis virus (CAEV). Virus Res. 2002, 87, 69–77. [Google Scholar] [CrossRef]
- Baszler, T.V.; Harwood, W.G.; Lester, K.L.; Davis, W.C.; Knowles, D.P. Characterization of caprine microglial cells and in vitro infection with caprine arthritis-encephalitis lentivirus. Lab. Invest. 1994, 70, 933–943. [Google Scholar]
- Adebayo, I.A.; Olaleye, O.D.; Awoniyi, T.A. Affinity (tropism) of caprine arthritis encephalitis virus for brain cells. Afr. J. Med. Med. Sci. 2008, 39, 89–93. [Google Scholar]
- Angelopoulou, K.; Brellou, G.D.; Vlemmas, I. Detection of maedi-visna virus in the kidneys of naturally infected sheep. J. Comp. Pathol. 2006, 134, 329–335. [Google Scholar] [CrossRef]
- Brellou, G.D.; Angelopoulou, K.; Poutahidis, T.; Vlemmas, I. Detection of maedi-visna virus in the liver and heart of naturally infected sheep. J. Comp. Pathol. 2007, 136, 27–35. [Google Scholar] [CrossRef]
- Capucchio, M.T.; Sanna, E.; Sanna, M.P.; Farigu, S.; Minelli, R.; Guarda, F. Maedi-visna virus detection in ovine third eyelids. J. Comp. Pathol. 2003, 129, 37–43. [Google Scholar] [CrossRef]
- Carrozza, M.L.; Mazzei, M.; Bandecchi, P.; Arispici, M.; Tolari, F. In situ PCR-associated immunohistochemistry identifies cell types harbouring the Maedi-Visna virus genome in tissue sections of sheep infected naturally. J. Virol. Methods 2003, 107, 121–127. [Google Scholar] [CrossRef]
- Sanchez, A.B.; Rodriguez, D.; Garzon, A.; Amorena, B.; Esteban, M.; Rodriguez, J.R. Visna/maedi virus Env protein expressed by a vaccinia virus recombinant induces cell-to-cell fusion in cells of different origins in the apparent absence of Env cleavage: Role of glycosylation and of proteoglycans. Arch. Virol. 2002, 147, 2377–2392. [Google Scholar] [CrossRef]
- Lyall, J.W.; Solanky, N.; Tiley, L.S. Restricted species tropism of maedi-visna virus strain EV-1 is not due to limited receptor distribution. J. Gen. Virol. 2000, 81, 2919–2927. [Google Scholar]
- Mselli-Lakhal, L.; Favier, C.; Leung, K.; Guiguen, F.; Grezel, D.; Miossec, P.; Mornex, J.F.; Narayan, O.; Querat, G.; Chebloune, Y. Lack of functional receptors is the only barrier that prevents caprine arthritis-encephalitis virus from infecting human cells. J. Virol. 2000, 74, 8343–8348. [Google Scholar] [CrossRef]
- Bruett, L.; Barber, S.A.; Clements, J.E. Characterization of a membrane-associated protein implicated in visna virus binding and infection. Virology 2000, 271, 132–141. [Google Scholar] [CrossRef]
- Dalziel, R.G.; Hopkins, J.; Watt, N.J.; Dutia, B.M.; Clarke, H.A.; McConnell, I. Identification of a putative cellular receptor for the lentivirus visna virus. J. Gen. Virol. 1991, 72, 1905–1911. [Google Scholar] [CrossRef]
- Hovden, A.O.; Sommerfelt, M.A. The influence of CD4 and CXCR4 on maedi-visna virus-induced syncytium formation. APMIS 2002, 110, 697–708. [Google Scholar] [CrossRef]
- Crane, S.E.; Buzy, J.; Clements, J.E. Identification of cell membrane proteins that bind visna virus. J. Virol. 1991, 65, 6137–6143. [Google Scholar]
- Hotzel, I.; Cheevers, W.P. A maedi-visna virus strain K1514 receptor gene is located in sheep chromosome 3p and the syntenic region of human chromosome 2. J. Gen. Virol. 2002, 83, 1759–1764. [Google Scholar]
- Crespo, H.; Reina, R.; Glaria, I.; Ramírez, H.; de Andrés, X.; Jáuregui, P.; Luján, L.; Martinez-Pomares, L.; Amorena, B.; de Andrés, D.F. Identification of the ovine mannose receptor and its possible role in Visna/Maedi virus infection. BMCVet. Res. 2011, 42, e28. [Google Scholar]
- Crespo, H.; Jáuregui, P.; Glaria, I.; Sanjose, L.; Polledo, L.; García-Marin, J.F.; Luján, L.; de Andrés, D.; Amorena, B.; Reina, R. Mannose receptor may be involved in small ruminant lentivirus pathogenesis. BMCVet. Res. 2012, 43, e43. [Google Scholar]
- Reina, R.; Berriatua, E.; Lujan, L.; Juste, R.; Sanchez, A.; de Andrés, D.; Amorena, B. Prevention strategies against small ruminant lentiviruses: An update. Vet. J. 2009, 182, 31–37. [Google Scholar] [CrossRef]
- Biescas, E.; Preziuso, S.; Bulgin, M.; DeMartini, J.C. Ovine lentivirus-associated leucomyelitis in naturally infected North American sheep. J. Comp. Pathol. 2005, 132, 107–116. [Google Scholar] [CrossRef]
- Storset, A.K.; Teig, A.; Rimstad, E. Detection of caprine arthritis-encephalitis virus RNA in macrophages by in situ hybridization using fluorescein-labelled single-stranded RNA probes. Vet. Microbiol. 1996, 52, 25–35. [Google Scholar] [CrossRef]
- Knowles, D.P., Jr.; Evermann, J.F.; Shropshire, C.; VanderSchalie, J.; Bradway, D.; Gezon, H.M.; Cheevers, W.P. Evaluation of agar gel immunodiffusion serology using caprine and ovine lentiviral antigens for detection of antibody to caprine arthritis-encephalitis virus. J. Clin. Microbiol. 1994, 32, 243–245. [Google Scholar]
- De Andres, D.; Klein, D.; Watt, N.J.; Berriatua, E.; Torsteinsdottir, S.; Blacklaws, B.A.; Harkiss, G.D. Diagnostic tests for small ruminant lentiviruses. Vet. Microbiol. 2005, 107, 49–62. [Google Scholar] [CrossRef]
- Knowles, D.P., Jr. Laboratory diagnostic tests for retrovirus infections of small ruminant. Vet. Clin. North. Am. Food Anim. Pract. 1997, 13, 1–11. [Google Scholar]
- Herrmann-Hoesing, L.M. Diagnostic assays used to control small ruminant lentiviruses. J. Vet. Diagn. Invest. 2010, 22, 843–855. [Google Scholar] [CrossRef]
- Gogolewski, R.P.; Adams, D.S.; McGuire, T.C.; Banks, K.L.; Cheevers, W.P. Antigenic cross-reactivity between caprine arthritis-encephalitis, visna and progressive pneumonia viruses involves all virion-associated proteins and glycoproteins. J. Gen. Virol. 1985, 66, 1233–1240. [Google Scholar] [CrossRef]
- Cheevers, W.P.; Cordery-Cotter, R.; McGuire, T.C.; DeMartini, J.C. Neutralizing antibody responses and evolution of antigenic variants in monozygotic twin lambs infected with phenotypically distinct ovine lentiviruses. Virology 1999, 258, 382–388. [Google Scholar] [CrossRef]
- Herrmann, L.M.; Cheevers, W.P.; Marshall, K.L.; McGuire, T.C.; Hutton, M.M.; Lewis, G.S.; Knowles, D.P. Detection of serum antibodies to ovine progressive pneumonia virus in sheep by using a caprine arthritis-encephalitis virus competitive-inhibition enzyme-linked immunosorbent assay. Clin. Diagn. Lab. Immunol. 2003, 10, 862–865. [Google Scholar]
- Herrmann, L.M.; Cheevers, W.P.; McGuire, T.C.; Adams, D.S.; Hutton, M.M.; Gavin, W.G.; Knowles, D.P. Competitive-inhibition enzyme-linked immunosorbent assay for detection of serum antibodies to caprine arthritis-encephalitis virus: Diagnostic tool for successful eradication. Clin. Diagn. Lab. Immunol. 2003, 10, 267–271. [Google Scholar]
- Herrmann-Hoesing, L.M.; Broughton-Neiswanger, L.E.; Gouine, K.C.; White, S.N.; Mousel, M.R.; Lewis, G.S.; Marshall, K.L.; Knowles, D.P. Evaluation of a caprine arthritis-encephalitis virus/maedi-visna virus indirect enzyme-linked immunosorbent assay in the serological diagnosis of ovine progressive pneumonia virus in U.S. sheep. Clin. Vaccine Immunol. 2010, 17, 307–310. [Google Scholar] [CrossRef]
- Carrozza, M.L.; Mazzei, M.; Lacerenza, D.; Del Chiaro, L.; Giammarioli, M.; Marini, C.; Rutili, D.; Rosati, S.; Tolari, F. Seroconversion against SU5 derived synthetic peptides in sheep experimentally infected with different SRLV genotypes. Vet. Microbiol. 2009, 137, 369–374. [Google Scholar] [CrossRef]
- Reina, R.; Grego, E.; Profiti, M.; Glaria, I.; Robino, P.; Quasso, A.; Amorena, B.; Rosati, S. Development of specific diagnostic test for small ruminant lentivirus genotype E. Vet. Microbiol. 2009, 138, 251–257. [Google Scholar] [CrossRef]
- Brinkhof, J.; van Maanen, C. Evaluation of five enzyme-linked immunosorbent assays and an agar gel immunodiffusion test for detection of antibodies to small ruminant lentiviruses. Clin. Vaccine Immunol. 2007, 14, 1210–1214. [Google Scholar] [CrossRef]
- Toft, N.; Akerstedt, J.; Tharaldsen, J.; Hopp, P. Evaluation of three serological tests for diagnosis of Maedi-Visna virus infection using latent class analysis. Vet. Microbiol. 2007, 120, 77–86. [Google Scholar] [CrossRef]
- Ramirez, H.; Roman, B.S.; Glaria, I.; Reina, R.; Hernandez, M.M.; de Andres, X.; Crespo, H.; Hichou, B.; Cianca, S.; Goni, C.; et al. Antibody-based diagnosis of small ruminant lentivirus infection in seminal fluid. Theriogenology 2009, 72, 1085–1096. [Google Scholar] [CrossRef]
- Zanoni, R.G.; Vogt, H.R.; Pohl, B.; Bottcher, J.; Bommeli, W.; Peterhans, E. An ELISA based on whole virus for the detection of antibodies to small-ruminant lentiviruses. Zentralbl. Veterinarmed. 1994, 41, 662–669. [Google Scholar]
- Saman, E.; Van Eynde, G.; Lujan, L.; Extramiana, B.; Harkiss, G.; Tolari, F.; Gonzalez, L.; Amorena, B.; Watt, N.; Badiola, J. A new sensitive serological assay for detection of lentivirus infections in small ruminants. Clin. Diagn. Lab. Immunol. 1999, 6, 734–740. [Google Scholar]
- Lacerenza, D.; Giammarioli, M.; Grego, E.; Marini, C.; Profiti, M.; Rutili, D.; Rosati, S. Antibody response in sheep experimentally infected with different small ruminant lentivirus genotypes. Vet. Immunol. Immunopathol. 2006, 112, 264–271. [Google Scholar] [CrossRef]
- Cardinaux, L.; Zahno, M.L.; Deubelbeiss, M.; Zanoni, R.; Vogt, H.R.; Bertoni, G. Virological and phylogenetic characterization of attenuated small ruminant lentivirus isolates eluding efficient serological detection. Vet. Microbiol. 2013, 162, 572–581. [Google Scholar] [CrossRef]
- Grego, E.; Bertolotti, L.; Carrozza, M.L.; Profiti, M.; Mazzei, M.; Tolari, F.; Rosati, S. Genetic and antigenic characterization of the matrix protein of two genetically distinct ovine lentiviruses. Vet. Microbiol. 2005, 106, 179–185. [Google Scholar] [CrossRef]
- Grego, E.; Profiti, M.; Giammarioli, M.; Giannino, L.; Rutili, D.; Woodall, C.; Rosati, S. Genetic heterogeneity of small ruminant lentiviruses involves immunodominant epitope of capsid antigen and affects sensitivity of single-strain-based immunoassay. Clin. Diagn. Lab. Immunol. 2002, 9, 828–832. [Google Scholar]
- Mordasini, F.; Vogt, H.R.; Zahno, M.L.; Maeschli, A.; Nenci, C.; Zanoni, R.; Peterhans, E.; Bertoni, G. Analysis of the antibody response to an immunodominant epitope of the envelope glycoprotein of a lentivirus and its diagnostic potential. J. Clin. Microbiol. 2006, 44, 981–991. [Google Scholar] [CrossRef]
- De Andrés, X.; Ramírez, H.; Bertolotti, L.; San Román, B.; Glaria, I.; Crespo, H.; Jáuregui, P.; Minguijón, E.; Juste, R.; Leginagoikoa, I.; et al. An insight into a combination of ELISA strategies to diagnose Small Ruminant Lentivirus infections. Vet. Immunol. Immunopathol. 2013, 152, 277–288. [Google Scholar] [CrossRef]
- Brinkhof, J.M.; Houwers, D.J.; Moll, L.; Dercksen, D.; van Maanen, C. Diagnostic performance of ELISA and PCR in identifying SRLV-infected sheep and goats using serum, plasma and milk samples and in early detection of infection in dairy flocks through bulk milk testing. Vet. Microbiol. 2010, 142, 193–198. [Google Scholar] [CrossRef]
- Motha, M.X.; Ralston, J.C. Evaluation of ELISA for detection of antibodies to CAEV in milk. Vet. Microbiol. 1994, 38, 359–367. [Google Scholar] [CrossRef]
- Plaza, M.; Sánchez, A.; Corrales, J.C.; De la Fe, C.; Contreras, A. Caprine arthritis encephalitis virus diagnosed by ELISA in lactating goats using milk samples. Small Rum. Res. 2009, 81, 189–192. [Google Scholar] [CrossRef]
- Brinkhof, J.M.A.; Houwers, D.J.; van Maanen, C. Development of a sample pooling strategy for the serodiagnosis of small ruminant lentiviral infections using the ELITEST-MVV ELISA. Small Rum. Res. 2007, 70, 194–199. [Google Scholar] [CrossRef]
- Reina, R.; Glaria, I.; Cianca, S.; Crespo, H.; Andres, X.D.; Goni, C.; Lasarte, J.M.; Lujan, L.; Amorena, B.; de Andres, D.F. Use of small ruminant lentivirus-infected rams for artificial insemination. Vet. J. 2010, 189, 106–107. [Google Scholar]
- Gudmundsson, B.; Bjarnadottir, H.; Kristjansdottir, S.; Jonsson, J.J. Quantitative assays for maedi-visna virus genetic sequences and mRNA’s based on RT-PCR with real-time FRET measurements. Virology 2003, 307, 135–142. [Google Scholar] [CrossRef]
- Zhang, Z.; Watt, N.J.; Hopkins, J.; Harkiss, G.; Woodall, C.J. Quantitative analysis of maedi-visna virus DNA load in peripheral blood monocytes and alveolar macrophages. J. Virol. Methods 2000, 86, 13–20. [Google Scholar] [CrossRef]
- Brinkhof, J.M.; van Maanen, C.; Wigger, R.; Peterson, K.; Houwers, D.J. Specific detection of small ruminant lentiviral nucleic acid sequences located in the proviral long terminal repeat and leader-gag regions using real-time polymerase chain reaction. J. Virol. Methods 2008, 147, 338–344. [Google Scholar] [CrossRef]
- Carrozza, M.L.; Mazzei, M.; Bandecchi, P.; Fraisier, C.; Perez, M.; Suzan-Monti, M.; de Andres, D.; Amorena, B.; Rosati, S.; Andresdottir, V.; et al. Development and comparison of strain specific gag and pol real-time PCR assays for the detection of Visna/maedi virus. J. Virol. Methods 2010, 165, 161–167. [Google Scholar] [CrossRef]
- Herrmann-Hoesing, L.M.; White, S.N.; Lewis, G.S.; Mousel, M.R.; Knowles, D.P. Development and validation of an ovine progressive pneumonia virus quantitative PCR. Clin. Vaccine Immunol. 2007, 14, 1274–1278. [Google Scholar] [CrossRef]
- Haase, A.T.; Retzel, E.F.; Staskus, K.A. Amplification and detection of lentiviral DNA inside cells. Proc. Natl. Acad. Sci. USA 1990, 87, 4971–4975. [Google Scholar] [CrossRef]
- Zanoni, R.; Pauli, U.; Peterhans, E. Detection of caprine arthritis-encephalitis- and maedi-visna viruses using the polymerase chain reaction. Experientia 1990, 46, 316–319. [Google Scholar] [CrossRef]
- Leroux, C.; Lerondelle, C.; Chastang, J.; Mornex, J.F. RT-PCR detection of lentiviruses in milk or mammary secretions of sheep or goats from infected flocks. Vet. Res. 1997, 28, 115–121. [Google Scholar]
- Alvarez, V.; Daltabuit-Test, M.; Arranz, J.; Leginagoikoa, I.; Juste, R.A.; Amorena, B.; de Andres, D.; Lujan, L.; Badiola, J.J.; Berriatua, E. PCR detection of colostrum-associated Maedi-Visna virus (MVV) infection and relationship with ELISA-antibody status in lambs. Res. Vet. Sci. 2006, 80, 226–234. [Google Scholar] [CrossRef]
- Barlough, J.; East, N.; Rowe, J.D.; Van Hoosear, K.; DeRock, E.; Bigornia, L.; Rimstad, E. Double-nested polymerase chain reaction for detection of caprine arthritis-encephalitis virus proviral DNA in blood, milk, and tissues of infected goats. J. Virol. Methods 1994, 50, 101–113. [Google Scholar] [CrossRef]
- Celer, V., Jr.; Celer, V.; Nejedla, E.; Bertoni, G.; Peterhans, E.; Zanoni, R.G. The detection of proviral DNA by semi-nested polymerase chain reaction and phylogenetic analysis of Czech Maedi-Visna isolates based on gag gene sequences. J. Vet. Med. B Infect. Dis. Vet. Public Health 2000, 47, 203–215. [Google Scholar] [CrossRef]
- Eltahir, Y.M.; Dovas, C.I.; Papanastassopoulou, M.; Koumbati, M.; Giadinis, N.; Verghese-Nikolakaki, S.; Koptopoulos, G. Development of a semi-nested PCR using degenerate primers for the generic detection of small ruminant lentivirus proviral DNA. J. Virol. Methods 2006, 135, 240–246. [Google Scholar] [CrossRef]
- Modolo, J.R.; Stachissini, A.V.M.; Padovani, C.R.; Araujo Júnior, J.P.; Castro, R.S.; Ravazzolo, A.P.; Leite, B.L.S. PCR associated with agar gel immunodiffusion assay improve caprine arthritis-encephalitis (CAEV) control. Small Rum. Res. 2009, 81, 18–20. [Google Scholar] [CrossRef]
- Reddy, P.G.; Sapp, W.J.; Heneine, W. Detection of caprine arthritis-encephalitis virus by polymerase chain reaction. J. Clin. Microbiol. 1993, 31, 3042–3043. [Google Scholar]
- Rimstad, E.; East, N.E.; Torten, M.; Higgins, J.; DeRock, E.; Pedersen, N.C. Delayed seroconversion following naturally acquired caprine arthritis-encephalitis virus infection in goats. Am. J. Vet. Res. 1993, 54, 1858–1862. [Google Scholar]
- Zanoni, R.G.; Cordano, P.; Nauta, I.M.; Peterhans, E. PCR for the detection of lentiviruses from small ruminants. Schweiz Arch. Tierheilkd. 1996, 138, 93–98. [Google Scholar]
- Leginagoikoa, I.; Minguijon, E.; Berriatua, E.; Juste, R.A. Improvements in the detection of small ruminant lentivirus infection in the blood of sheep by PCR. J. Virol. Methods 2009, 156, 145–149. [Google Scholar] [CrossRef]
- Extramiana, A.B.; González, L.; Cortabarría, N.; García, M.; Juste, R.A. Evaluation of a PCR technique for the detection of Maedi-Visna proviral DNA in blood, milk and tissue samples of naturally infected sheep. Small Rum. Res. 2002, 44, 109–118. [Google Scholar] [CrossRef]
- Ali Al Ahmad, M.Z.; Fieni, F.; Pellerin, J.L.; Guiguen, F.; Cherel, Y.; Chatagnon, G.; Bouzar, A.B.; Chebloune, Y. Detection of viral genomes of caprine arthritis-encephalitis virus (CAEV) in semen and in genital tract tissues of male goat. Theriogenology 2008, 69, 473–480. [Google Scholar] [CrossRef]
- Travassos, C.; Benoit, C.; Valas, S.; da Silva, A.; Perrin, G. Caprine artritis-encephalitis virus in semen of naturally infected bucks. Small Rum. Res. 1999, 32, 101–106. [Google Scholar] [CrossRef]
- Brodie, S.J.; Pearson, L.D.; Snowder, G.D.; DeMartini, J.C. Host-virus interaction as defined byamplification of viral DNA and serology in lentivirus-infected sheep. Arch. Virol. 1993, 130, 413–428. [Google Scholar] [CrossRef]
- Zanoni, R.; Nauta, I.M.; Kuhnert, P.; Pauli, U.; Pohl, B.; Peterhans, E. Genomic heterogeneity of small ruminant lentiviruses detected by PCR. Vet. Microbiol. 1992, 33, 241–351. [Google Scholar]
- Rosati, S.; Kwang, J.; Keen, J.E. Genome analysis of North American small ruminant lentiviruses by polymerase chain reaction and restriction enzyme analysis. J. Vet. Diagn. Invest. 1995, 7, 437–443. [Google Scholar] [CrossRef]
- Johnson, L.K.; Meyer, A.L.; Zink, M.C. Detection of ovine lentivirus in seronegative sheep by in situ hybridization, PCR, and cocultivation with susceptible cells. Clin. Immunol. Immunopathol. 1992, 65, 254–260. [Google Scholar] [CrossRef]
- Wagter, L.H.; Jansen, A.; Bleumink-Pluym, N.M.; Lenstra, J.A.; Houwers, D.J. PCR detection of lentiviral GAG segment DNA in the white blood cells of sheep and goats. Vet. Res. Commun. 1998, 22, 355–362. [Google Scholar] [CrossRef]
- Karanikolaou, K.; Angelopoulou, K.; Papanastasopoulou, M.; Koumpati-Artopiou, M.; Papadopoulos, O.; Koptopoulos, G. Detection of small ruminant lentiviruses by PCR and serology tests in field samples of animals from Greece. Small Rum. Res. 2005, 58, 181–187. [Google Scholar] [CrossRef]
- Brodie, S.J.; Marcom, K.A.; Pearson, L.D.; Anderson, B.C.; de la Concha-Bermejillo, A.; Ellis, J.A.; DeMartini, J.C. Effects of virus load in the pathogenesis of lentivirus-induced lymphoid interstitial pneumonia. J. Infect. Dis. 1992, 166, 531–541. [Google Scholar] [CrossRef]
- Clavijo, A.; Thorsen, J. Application of polymerase chain reaction for the diagnosis of caprine arthritis-encephalitis. Small Rum. Res. 1996, 22, 69–77. [Google Scholar] [CrossRef]
- Kuzmak, J.; Kedziora, A.; Rola, M.; Kozaczynska, B.; Chebloune, Y.; Gallay, K. Evaluation of PCR and PCR/hybridation method for the detection of caprine arthritis-encephalitis virus infection in goats. Bull. Vet. Inst. Pulawy 2003, 47, 293–300. [Google Scholar]
- Herrmann-Hoesing, L.M.; Palmer, G.H.; Knowles, D.P. Evidence of proviral clearance following postpartum transmission of an ovine lentivirus. Virology 2007, 362, 226–234. [Google Scholar] [CrossRef]
- Barquero, N.; Arjona, A.; Domenech, A.; Toural, C.; de las Heras, A.; Fernandez-Garayzabal, J.F.; Ruiz-Santa Quiteria, J.A.; Gomez-Lucia, E. Diagnostic performance of PCR and ELISA on blood and milk samples and serological survey for small ruminant lentiviruses in central Spain. Vet. Rec. 2011, 168. [Google Scholar] [CrossRef]
- Brinkhof, J.M.; Moll, L.; van Maanen, C.; Houwers, D.J. Use of serology and polymerase chain reaction for the rapid eradication of small ruminant lentivirus infections from a sheep flock: A case report. Res. Vet. Sci. 2010, 88, 41–43. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ramírez, H.; Reina, R.; Amorena, B.; Andrés, D.D.; Martínez, H.A. Small Ruminant Lentiviruses: Genetic Variability, Tropism and Diagnosis. Viruses 2013, 5, 1175-1207. https://doi.org/10.3390/v5041175
Ramírez H, Reina R, Amorena B, Andrés DD, Martínez HA. Small Ruminant Lentiviruses: Genetic Variability, Tropism and Diagnosis. Viruses. 2013; 5(4):1175-1207. https://doi.org/10.3390/v5041175
Chicago/Turabian StyleRamírez, Hugo, Ramsés Reina, Beatriz Amorena, Damián De Andrés, and Humberto A. Martínez. 2013. "Small Ruminant Lentiviruses: Genetic Variability, Tropism and Diagnosis" Viruses 5, no. 4: 1175-1207. https://doi.org/10.3390/v5041175
APA StyleRamírez, H., Reina, R., Amorena, B., Andrés, D. D., & Martínez, H. A. (2013). Small Ruminant Lentiviruses: Genetic Variability, Tropism and Diagnosis. Viruses, 5(4), 1175-1207. https://doi.org/10.3390/v5041175