Genome Scale Transcriptomics of Baculovirus-Insect Interactions

Abstract
:1. Introduction
2. Host Genes/Pathways that Baculoviruses Require for Successful Infection Processes in Insect Cells
2.1. Transcriptomic Studies

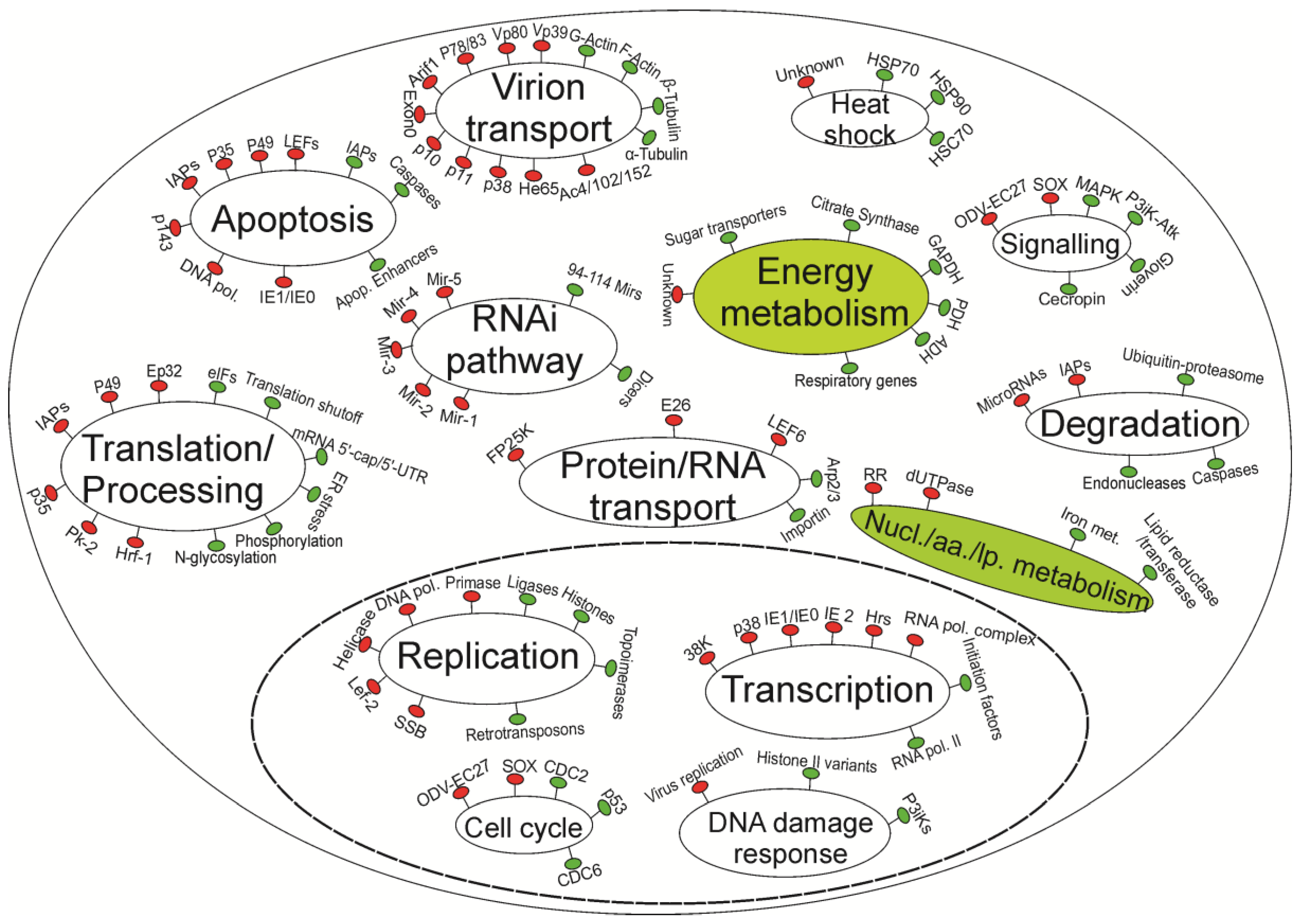
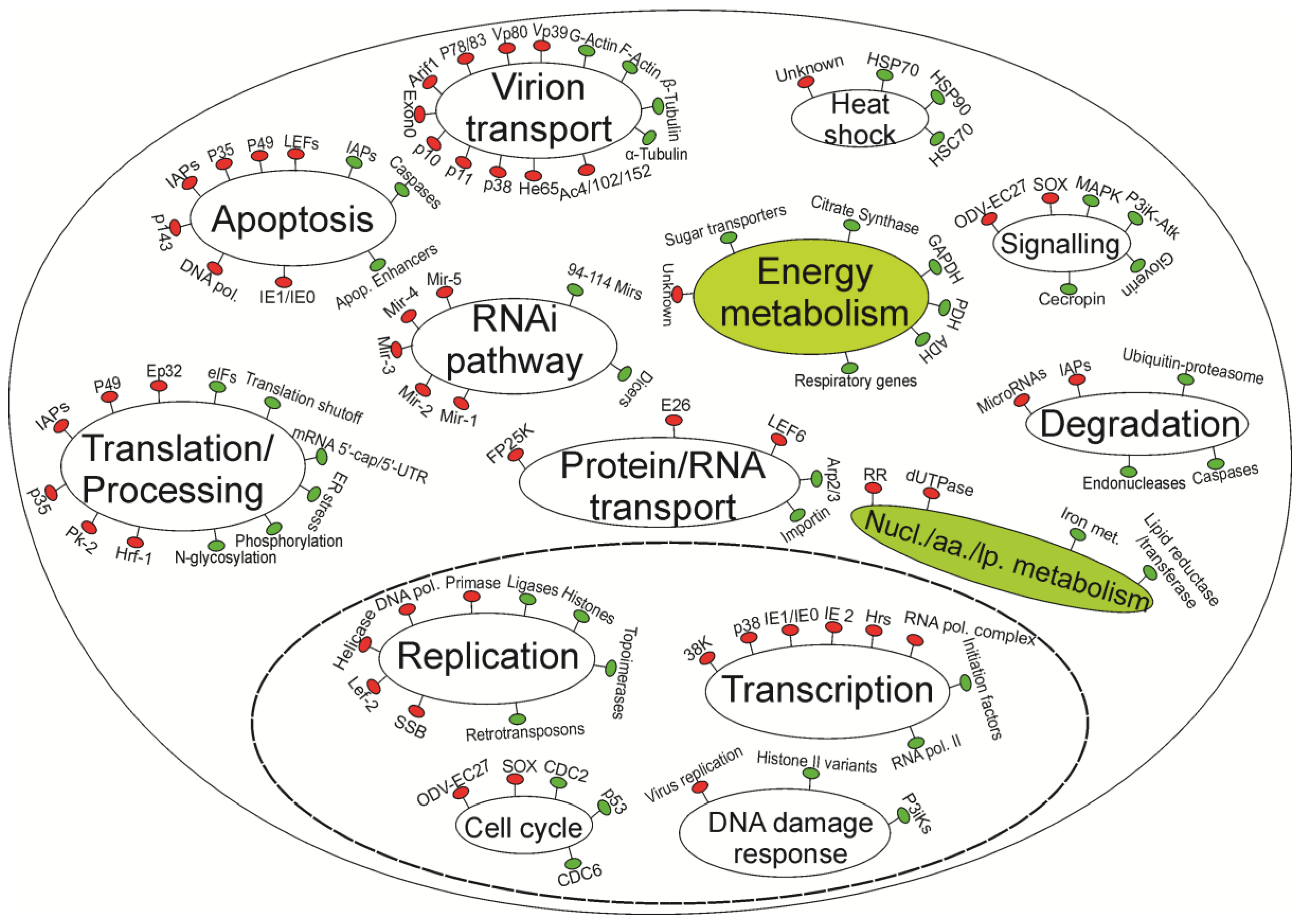{kind=link}
| Functional groups | Baculovirus genes/pathways | Baculovirus strains [6] | Functions | Ref. |
|---|---|---|---|---|
| Virus genes interacting with host cell receptors | GP-64, F protein | GP-64 only in group I alpha NPV‚ while the F protein in group II alpha-, beta- and delta- NPV | Virus-cell receptor attachment, facilitate entry by clathrin-mediated endocytosis processes | [34,35] |
| Per OS infectivity factors (Pif-1, 2, 3, p-74)‚ and possibly Pif-4 and 5 | All 5 Pifs and p-74 are core baculorius genes, and found in other invertebrate DNA viruses that replicate in the nucleus | Pif-1, 2, and 3 and p74 form a complex and facilitate ODV binding to midgut epithelial cell receptors | [36] | |
| Apoptosis | IAP-1, IAP-2, IAP-3, IAP-4, and IAP-5 | IAP1-4 in both NPVs and Granulosis viruses (GVs), while IAP-5 in GVs only. Each baculovirus strain has several/not all IAPs. IAP orthologs found in a number of hosts | Baculovirus IAPs mediate protein-protein interactions to block selected caspases. The IAP RING domain functions as an E3 ubiquitin ligase to trigger proteasome degradation of targeted caspases | [37] |
| P35 | AcMPNV, BmNPV, Culex nigripalpus NPV, Leucania separata MNPV, Maruca vitrata MNPV, T. ni MNPV and Clostera anachoreta | Binds to and inactivates host’s effector caspases | [38] | |
| p49 | SpltMNPV, LsMNPV, SlNPV, AcMNPV, and HearNPV | Inhibits host’s initiator caspases (upstream of p35 but downstream of IAPs) and several host’s effector caspases | [39] | |
| Replicative lefs (lef-1, 2, 3, and 11) p-143, DNA pol and IE1/IE0 | Lef-3 in Lepidopteran NPV and GV; Lef-1, Lef-2, Lef-11, and p-143 in all baculoviruses (except for Lef-11 not in CuniNPV), IE1/IE0 in all group I and II alpha baculoviruses | Trigger host DNA damage response and induce apoptosis | [40] | |
| Cell cycle | ODV-EC27 (A virus multifunctional Cyclin) | In all baculoviruses | Interacts with host’s cdc-2 for cell cycle arrest at G2/M phase, or with host’s cdc-6 to override host check-point to allow DNA replication | [41] |
| P33-sulfhydryloxidase (SOX) | In all baculoviruses | Forms stable complex with host’s p53 protein, preventing p53-induced apoptosis | [42] | |
| Cytoskeleton and nucleocapsid transport | Protein kinase-1, Protein kinase-2 | PK-1 found in Lepidopteran NPVs, and GVs, similar to some insect PK; PK-2 found in AcMNPV, BmNPV, PlxyNPV and RoMNPV | Actin cytoskeleton remodeling (protein-protein interaction prediction) | [11] |
| Arif-1 | All group I and most group II Alpha baculoviruses | Accumulates F-actin at the plasma membrane | [43] | |
| VP80 (a Paramyosin-like protein)/P78-83/VP39 | VP80 and p78/83 in all group I and II Lepidopteran NPVs; VP39 in all baculovirus genomes | Interact with host’s F-actin filaments to transport nucleocapsids in the cytoplasm | [44,45] | |
| VP80 | Vp80 found in all group I and II Lepidopteran NPVs | Interact with myosin motor proteins and F-actin to transport nucleocapsids to the nucleus periphery | [44] | |
| IE-1, PE38, HE65, Ac004, Ac102, Ac152 | IE1 all baculoviruses; PE38 in all Group I NPV and four GV genomes | Accumulate host’s monomeric G-Actin into nucleus | [46] | |
| P78/83 (N-WASP-homologous protein) and ODV-C42 | p78/83 all group I and II Lepidopteran NPVs | At early infection, transports nucleocapsid into nucleus by activating nuclear actin polymerization via an actin related protein (Arp2/3) complex. At late infection, facilitates actin assembly to form F-filament inside nucleus | [47,48] | |
| EXON0 | In all Lepidopteran NPVs | Interacts with β-tubulin to facilitate binding of nucleocapsids to microtubules | [49] | |
| P10 | In all group I and II NPVs and most GVs | Interacts with α-tubulin and mediates nuclear disintegration and cell lysis | [50] | |
| Nucleo-cytoplasmic transport of viral proteins | FP25K and E26 | FP25K in all Lepidopteran NPVs and GVs. E26 in group I Lepidopteran NPV | Together with host Importin-α-16, transport viral proteins into the inner nuclear membrane (INM) | [51] |
| Metabolism | ADP ribose pyrophosphatase (Ac38) | All Lepidopteran NPVs and GVs | The enzyme hydrolyzes ADP-ribose, an intermediate of metabolism of NAD+, mono- or poly-ADP-ribosylated proteins and cyclic ADP-ribose, thereby conferring detoxification effects | [52] |
| P33-sulfhydryl oxidase (SOX) | In all baculoviruses | Flavin adenine dinucleotide (FAD)-binding sulfhydryl oxidase can play roles in protein disulphide bond formation and protection from oxidative stress | [53,54] | |
| Super oxide dismutase (SOD) | In most Lepidopteran baculoviruses | Converts superoxide into Hydrogen peroxide (possibly active in BmNPV, but this activity is not confirmed in AcMNPV) | [55,56] | |
| Replication | Ribonucleotide reductase | Three GVs, 10 NPVs group II, OpMNPV and LdMNPV | Catalysis of ribonucleotides to deoxyribonucleotides for DNA synthesis | [57,58] |
| DNA polymerase complex (Dnapol, helicase, primase, SSB, and LEF-2) | All baculoviruses | May require host’s DNA topoimerases and DNA ligases | [6,59] | |
| dUTPase | In nine group II NPVs, OpMNPV, and two GV genomes | Prevents incorporation of dUTP into DNA | [58,60] | |
| Transcription | IE1/IE0, IE2, hrs, ADPRase (ADP-ribose pyrophosphatase) | IE1/IE0, hrs and ADPRase in all baculoviruses. IE2 in all Group I Lepidopteran NPVs but not others. pe38 in all Group I NPV and four GV genomes | Bind to host transcription factors | [61] |
| Lef-6 | All Lepidopteran NPVs and GVs | Lef-6 has a TAP (TIP associating domain), which can interact with nuclearporins for mRNA export to the cytoplasm | [6,62] | |
| Ac98-38 K protein | All baculoviruses | Predicted to have carboxyl terminal domain (CTD) phosphatase activities that negatively regulate RNA polymerase II by inhibiting RNA elongation | [6] | |
| Translation arrest | P35, IAPs and P49 | As mentioned before | Enhance early host translation arrest | [63] |
| Protein kinase 2 (Pk-2) | PK2 found in AcMNPV, BmNPV, PlxyNPV and RoMNPV | Represses translation arrest, which is caused by host eIF2α kinase, by blocking eIF2α access to translation initiation factors | [64,65] | |
| Host range factor 1 (Hrf-1), | Only found in viruses of Lymantria dispar host, including LdMNPV and Orgyia pseudotsugata MNPV | Inhibits translational arrest by an unknown mechanism | [66] | |
| Hycu-ep32 gene | Hyphantri acunea NPV and OpMNPV | Induces host translation arrest by an unknown mechanism | [67] | |
| IAP-1 and IAP-2 | As mentioned before | IAP1 and IAP2 possess ubiquitin ligase activities, enabling polyubiquitination of insect proteins, thus marking them for degradation | [68] | |
| Growth & development | Protein tyrosine phosphatase (PTP) | All Lepidopteran Group I NPVs, not others, orthologs found in insect host | Induces host hyperactive behaviours | [69,70] |
| Viral Fibroblast growth factor (vFGF) | All baculoviruses, orthologs found in insect hosts | Increases host larvae motility by facilitating systemic infection | [71,72] | |
| Chitinase and Cathepsin | Chitinase and Cathepsin in all Group I (except AgMNPV), all Group II (except AdhoNPV for Chitinase) and four GVs | Chitinase breaks larvae chitin layer, Cathepsin is a viral proteinase | [73] | |
| Ecdysteroid UDP glucosyl transferase (EGT) | All Lepidopteran Group I NPV, not others | Prevents moulting to extend insect life and virus propagation time (transfers glucose group to inactivate insect molting hormone ecdysteroids), induces host hyperactive behaviours | [74,75] | |
| MicroRNA | BmNPV-miR-1 | Conserved in AcMNPV, BomaNPV, PxMNPV, RoMNPV, and MaviNPV | Down-regulates the transport of host small-RNA from the nucleus to the cytoplasm, thereby reducing active population of host small RNAs | [76] |
| BmNPV-miR-2 to 4 | Conserved in AcMNPV, BomaNPV, PxMNPV, RoMNPV, and MaviNPV | Potentially targets 8 viral genes and 64 host genes | [77] |
| Functional groups | Insect genes/pathways | Functions | Virus-host systems and expression time | Ref. |
|---|---|---|---|---|
| Immune genes | Gloverin | An antibacterial and antiviral protein that interacts with the lipid envelope surrounding viral nucleocapsids | BmNPV-B. mori larvae and AcMNPV-S. exigua larvae (induced from 6–12 h.p.i) | [19,31,78] |
| Cecropin | A cationic antimicrobial peptide that has positively charged regions in its α-helical peptide and interferes with the lipid membrane | H. virescens larvae-HzSNPV and HzAM1 cells-HearNPV(induced from 12 h.p.i) | [7,33,79] | |
| Apoptosis genes | Down regulation of host IAPs, and up-regulation of host apoptosis enhancers and Caspases trigger apoptosis upon virus infections | Epiphyas postvittana larvae-EppoNPV (5 d.p.i); HzAM1 cells-HearNPV (12 h.p.i); and Sf21-AcMNPV (before 24 h.p.i) | [15,18,80] | |
| Signal transduction | Phosphatidylinositol 3 kinases (Pi3K)-Akt pathway | Elevation of this pathway prevents apoptosis and creates inductive environment for virus propagation | AcMNPV-Sf9 (induced from 1–18 h.p.i) | [81,82] |
| MAPK pathways | Extracellular signal-regulated kinase (ERK) and c-Jun NH2-terminal kinase (JNK) pathways are activated at late infection and important for virus production | BmN4 cells-BmNPV (induced from 4–24 h.p.i) | [11,83] | |
| DNA damage response kinases | Triggered by virus replication‚ leading to cell death | AcMNPV-Sf21 (induced from 2–24 h.p.i) | [9] | |
| Metabolic genes | ABC transporters and sugar transporters | ABC transporters transport a broad spectrum of substrates, including degradation products from cytosol to ER | BmNPV-B. mori cell line and AcMNPV-S. exigua larvae (induced from 2–12 h.p.i); HearNPV-HzAM1 (induced from 12–18 h.p.i) | [7,19,32,84] |
| Citrate synthetase | Important for energy generation (a key enzyme in the Citric acid cycle, TCA) | AcMNPV-Sf9 and BmNPV-B. mori cell line (induced from 2 h.p.i) | [14,85] | |
| Pyruvate dehydrogenase/Aldehyde dehydrogenase | Important for energy generation and diversion of substrates to lipid biosynthesis | AcMNPV-Sf9 induced from (6 h.p.i) | [12] | |
| Lipid reductases and lipid desaturases | Fatty acid metabolism | HzNPV-H. virescens (induced from 12 h.p.i) | [33] | |
| Genes involved in cellular iron (iron ion transport, ferric iron binding, and cellular iron ion homeostasis) | Iron is important for processes such as DNA replication and ATP generation | BmNPV-B. mori cell line (induced from 3–6 h.p.i); H. virescens larvae-HzSNPV (induced from 24–72 h.p.i) | [11,86] | |
| Mitochondrial respiratory genes | Important for energy generation | BmNPV-B. mori cell line (induced from 1.5–24 h.p.i) | [7,11] | |
| Translation | Heat shock protein (HSP) 70, HSP90, and Heat shock protein cognate (HSC) 70 | Protein folding and facilitate several cellular processes conducive for virus replication such as ubiquitin-proteasome pathway | BmNPV-B. mori larvae (6–12 h.p.i); AcMNPV-Sf21 cell line (induced from 6–48 h.p.i);AcMNPV-S. exigua larvae (induced from 12 h.p.i) | [10,19,31,87] |
| ER proteins (reduced) | ER stress | AcMNPV-Sf21 and AcMNPV-Sf9 (induced from 12–48 h.p.i) | [10,12] | |
| Translation initiation factors (TIFs) | Enhance translation | BmNPV-B. mori cell line (induced from 1.5–24 h.p.i) | [11,19] | |
| eIF2α | Phosphorylation of eIF2α causes translation arrest | AcMNPV-Sf9 cell line (Before 36 h.p.i) | [64,88] | |
| Replication | Histone genes | Regulate host chromatin structure, which affects DNA replication | BmNPV-B. mori cell line (induced from 1.5–24 h.p.i) | [11] |
| Transcription | Host’s polyhedrin promoter binding protein (PPBP) | Binds to promoters of both p10 and polyhedrin genes to enhance their transcription | AcMNPV-Sf9 (induced late) | [89] |
| Transcription initiation factors | Enhance transcription | BmNPV-B. mori cell line (induced from 6–24 h.p.i); HearNPV-H.zea (induced from 18 h.p.i) | [7,11] | |
| mRNA and protein degradation | Alkaline nuclease | mRNA degradation | E. postvittana larvae-EppoNPV (induced from 5 d.p.i); BmNPV-B. mori cell line (induced from 6 h.p.i) | [15,32] |
| Ubiquitin-proteasome pathway | Protein degradation | BmNPV-B. mori cell line (induced from 2–6 h.p.i); Sf9-AcMNPV (induced from 1.5–48 h.p.i) | [ 11,14,90] | |
| Cytoskeleton | Dynein | A motor protein involved in microtubule transport | BmNPV-B. mori cell line (induced from 6–12 h.p.i) | [32] |
| Development | Juvenile hormones | Maintaining juvenile hormones at high level extends growth and inhibits moulting | H. virescens larvae-HzSNPV (induced from 12 h.p.i); E. postvittana larvae-EppoNPV (induced from 5 d.p.i) | [15,33] |
| MicroRNAs | miRNA (90 miRNAs in Sf, 114 in B.mori) | Play roles in antiviral response by degrading viral transcripts (e.g. bmo-miR-8 potentially targets IE1) | Sf9-AcMNPV (Usually induced at late infection, 24–72 h.p.i) | [13,91] |
| Dicer 2 | Produce viral short interfering siRNAs that degrade viral transcripts | H. armigera larvae—HearNPV (induced from 48 h.p.i) | [92] | |
| Transposition of host DNA | Host transposable elements (retrotransposons), reverse transcriptase, gag/pol-like proteins | DNA transposition into baculovirus genomes, contributing to virus genome instability. | Sf9 cells, T.ni cells, H. virescens larvae-HzSNPV and AcMNPV-S. exigua larvae (During in vitro serial passaging) | [19,33,93,94] |
| Detoxification | Glutathione S-transferase (GST) | Convert glutathione into water-soluble, less toxic metabolites | AcMNPV-S. exigua larvae (induced from 12 h.p.i) | [19] |
2.2. Metabolism
2.3. Translation
2.4. Transport
2.5. Replication
2.6. Transcription
2.7. Immune responses
3. Manipulation of Host Cells or Larvae to Study Baculovirus-Insect Interactions
4. Conclusions and Future Perspectives
Acknowledgments
Conflicts of Interest
References
- Van Oers, M.M.; Vlak, J.M. Baculovirus genomics. Curr. Drug Targets 2007, 8, 1051–1068. [Google Scholar] [CrossRef]
- Thiem, S.M.; Cheng, X.-W. Baculovirus Host-Range. Virol. Sin. 2009, 24, 436–457. [Google Scholar] [CrossRef]
- Chen, C.Y.; Wu, H.H.; Chen, C.P.; Chern, S.R.; Hwang, S.M.; Huang, S.F.; Lo, W.H.; Chen, G.Y.; Hu, Y.C. Biosafety assessment of human mesenchymal stem cells engineered by hybrid baculovirus vectors. Mol. Pharm. 2011, 8, 1505–1514. [Google Scholar] [CrossRef]
- Chen, C.Y.; Lin, C.Y.; Chen, G.Y.; Hu, Y.C. Baculovirus as a gene delivery vector: Recent understandings of molecular alterations in transduced cells and latest applications. Biotechnol. Adv. 2011, 29, 618–631. [Google Scholar] [CrossRef]
- Mena, J.A.; Kamen, A.A. Insect cell technology is a versatile and robust vaccine manufacturing platform. Expert Rev. Vaccines 2011, 10, 1063–1081. [Google Scholar] [CrossRef]
- Rohrmann, G.F. Baculovirus Molecular Biology; National Library of Medicine, National Center for Biotechnology Information: Bethesda, MD, USA, 2011, 2nd ed. [Online]. Available online: http://www.ncbi.nlm.nih.gov/books/NBK49500/ (accessed 2 June 2013).
- Nguyen, Q.; Chan, L.C.; Nielsen, L.K.; Reid, S. Genome scale analysis of differential mRNA expression of Helicoverpa zea insect cells infected with a H. armigera baculovirus. Virology 2013, 444, 158–170. [Google Scholar] [CrossRef]
- Chen, Y.-R.; Zhong, S.; Fei, Z.; Hashimoto, Y.; Xiang, J.Z.; Zhang, S.; Blissard, G.W. The transcriptome of the baculovirus Autographa californica Multiple Nucleopolyhedrovirus (AcMNPV) in Trichoplusia ni cells. J. Virol. 2013, 87, 6391–6405. [Google Scholar] [CrossRef]
- Mitchell, J.K.; Friesen, P.D. Baculoviruses modulate a proapoptotic DNA damage response to promote virus multiplication. J. Virol. 2012, 86, 13542–13553. [Google Scholar] [CrossRef]
- Salem, T.Z.; Zhang, F.; Xie, Y.; Thiem, S.M. Comprehensive analysis of host gene expression in Autographa californica nucleopolyhedrovirus-infected Spodoptera frugiperda cells. Virology 2011, 412, 167–178. [Google Scholar] [CrossRef]
- Xue, J.; Qiao, N.; Zhang, W.; Cheng, R.-L.; Zhang, X.-Q.; Bao, Y.-Y.; Xu, Y.-P.; Gu, L.-Z.; Han, J.-D.J.; Zhang, C.-X. Dynamic interactions between Bombyx mori nucleopolyhedrovirus and its host cells revealed by transcriptome analysis. J. Virol. 2012, 86, 7345–7359. [Google Scholar] [CrossRef]
- Carinhas, N.; Robitaille, A.M.; Moes, S.; Carrondo, M.J.T.; Jenoe, P.; Oliveira, R.; Alves, P.M. Quantitative proteomics of Spodoptera frugiperda cells during growth and baculovirus infection. PLoS One 2011, 6, e26444. [Google Scholar]
- Mehrabadi, M.; Hussain, M.; Asgari, S. MicroRNAome of Spodoptera frugiperda cells (Sf9) and its alteration following baculovirus infection. J. Gen. Virol. 2013, 94, 1385–1397. [Google Scholar] [CrossRef]
- Iwanaga, M.; Shimada, T.; Kobayashi, M.; Kang, W. Identification of differentially expressed host genes in Bombyx mori nucleopolyhedrovirus infected cells by using subtractive hybridization. Appl. Entomol. Zool. 2007, 42, 151–159. [Google Scholar] [CrossRef]
- Gatehouse, H.S.; Poulton, J.; Markwick, N.P.; Gatehouse, L.N.; Ward, V.K.; Young, V.L.; Luo, Z.; Schaffer, R.; Christeller, J.T. Changes in gene expression in the permissive larval host lightbrownapple moth (Epiphyas postvittana, Tortricidae) in response to EppoNPV (Baculoviridae) infection. Insect Mol. Biol. 2009, 18, 635–648. [Google Scholar] [CrossRef]
- Van Oers, M.M.; Doitsidou, M.; Thomas, A.A.M.; de Maagd, R.A.; Vlak, J.M. Translation of both 5'TOP and non-TOP host mRNAs continues into the late phase of baculovirus infection. Insect Mol. Biol. 2003, 12, 75–84. [Google Scholar] [CrossRef]
- Huynh, H.; Tran, T.B.; Chan, L.L.; Nielsen, L.; Reid, S. Decline in baculovirus-expressed recombinant protein production with increasing cell density is strongly correlated to impairment of virus replication and mRNA expression. Appl. Microbiol. Biotechnol. 2013, 97, 5245–5257. [Google Scholar] [CrossRef]
- Nguyen, Q.; Palfreyman, R.W.; Chan, L.C.L.; Reid, S.; Nielsen, L.K. Transcriptome sequencing of and microarray development for a Helicoverpa zea cell line to investigate in vitro insect cell-baculovirus interactions. PLoS One 2012, 7, e36324. [Google Scholar]
- Choi, J.Y.; Roh, J.Y.; Wang, Y.; Zhen, Z.; Tao, X.Y.; Lee, J.H.; Liu, Q.; Kim, J.S.; Shin, S.W.; Je, Y.H. Analysis of genes expression of Spodoptera exigua Larvae upon AcMNPV infection. PLoS One 2012, 7, e42462. [Google Scholar] [CrossRef]
- Yamagishi, J.; Isobe, R.; Takebuchi, T.; Bando, H. DNA microarrays of baculovirus genomes: Differential expression of viral genes in two susceptible insect cell lines. Arch. Virol. 2003, 148, 587–597. [Google Scholar] [CrossRef]
- Jiang, S.S.; Chang, I.S.; Huang, L.W.; Chen, P.C.; Wen, C.C.; Liu, S.C.; Chien, L.C.; Lin, C.Y.; Hsiung, C.A.; Juang, J.L. Temporal transcription program of recombinant autographa californica multiple nucleopolyhedrosis Virus. J. Virol. 2006, 80, 8989–8999. [Google Scholar] [CrossRef]
- The International Silkworm Genome Consortium. The genome of a lepidopteran model insect, the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1036–1045. [Google Scholar] [CrossRef]
- You, M.; Yue, Z.; He, W.; Yang, X.; Yang, G.; Xie, M.; Zhan, D.; Baxter, S.W.; Vasseur, L.; Gurr, G.M.; et al. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 2013, 45, 220–225. [Google Scholar] [CrossRef]
- Deng, Y.; Dong, Y.; Thodima, V.; Clem, R.J.; Passarelli, A.L. Analysis and functional annotation of expressed sequence tags from the fall armyworm Spodoptera frugiperda. BMC Genomics 2006, 7. [Google Scholar] [CrossRef]
- Landais, I.; Ogliastro, M.; Mita, K.; Nohata, J.; Ferber, L.M.; Cerutti, M.D.; Shimada, T.; Fournier, P.; Devauchelle, G. Annotation pattern of ESTs from Spodoptera frugiperda Sf9 cells and analysis of the ribosomal protein genes reveal insect-specific features and unexpectedly low codon usage bias. Bioinformatics 2003, 19, 2343–2350. [Google Scholar] [CrossRef]
- Shelby, K.S.; Popham, H.J.R. Analysis of ESTs generated from immune-stimulated hemocytes of larval Heliothis virescens. J. Invertebr. Pathol. 2009, 101, 86–95. [Google Scholar] [CrossRef]
- Pauchet, Y.; Wilkinson, P.; Vogel, H.; Nelson, D.R.; Reynolds, S.E.; Heckel, D.G.; Ffrench-Constant, R.H. Pyrosequencing the Manduca sexta larval midgut transcriptome: Messages for digestion, detoxification and defence. Insect Mol. Biol. 2009, 19, 61–75. [Google Scholar]
- Vogel, H.; Heidel, A.J.; Heckel, D.G.; Groot, A.T. Transcriptome analysis of the sex pheromone gland of the noctuid moth Heliothis virescens. BMC Genomics 2010, 11. [Google Scholar] [CrossRef]
- Vogel, H.; Altincicek, B.; Glockner, G.; Vilcinskas, A. A comprehensive transcriptome and immune-gene repertoire of the lepidopteran model host Galleria mellonella. BMC Genomics 2011, 12. [Google Scholar] [CrossRef]
- Sparks, M.E.; Gundersen-Rindal, D.E. The Lymantria dispar IPLB-Ld652Y cell line transcriptome comprises diverse virus-associated transcripts. Viruses 2011, 3, 2339–2350. [Google Scholar] [CrossRef]
- Bao, Y.Y.; Lu, Z.Y.; Liu, Z.B.; Xue, J.; Xu, Y.P.; Zhang, C.X. Comparative analysis of Bombyx mori nucleopolyhedrovirus responsive genes in fat body and haemocyte of B. mori resistant and susceptible strains. Insect Mol. Biol. 2010, 19, 347–358. [Google Scholar] [CrossRef]
- Sagisaka, A.; Fujita, K.; Nakamura, Y.; Ishibashi, J.; Noda, H.; Imanishi, S.; Mita, K.; Yamakawa, M.; Tanaka, H. Genome-wide analysis of host gene expression in the silkworm cells infected with Bombyx mori nucleopolyhedrovirus. Virus Res. 2010, 147, 166–175. [Google Scholar] [CrossRef]
- Breitenbach, J.E.; Shelby, K.S.; Popham, H.J.R. Baculovirus induced transcripts in hemocytes from the larvae of Heliothis virescens. Viruses 2011, 3, 2047–2064. [Google Scholar] [CrossRef]
- Wang, M.L.; Yin, F.F.; Shen, S.; Tan, Y.; Deng, F.; Vlak, J.M.; Hu, Z.H.; Wang, H.L. Partial functional rescue of Helicoverpa armigera single nucleocapsid nucleopolyhedrovirus infectivity by replacement of F protein with GP64 from Autographa californica multicapsid nucleopolyhedrovirus. J. Virol. 2010, 84, 11505–11514. [Google Scholar] [CrossRef]
- Long, G.; Pan, X.; Kormelink, R.; Vlak, J.M. Functional entry of baculovirus into insect and mammalian cells is dependent on clathrin-mediated endocytosis. J. Virol. 2006, 80, 8830–8833. [Google Scholar] [CrossRef]
- Peng, K.; van Oers, M.M.; Hu, Z.; van Lent, J.W.; Vlak, J.M. Baculovirus per os infectivity factors form a complex on the surface of occlusion-derived virus. J. Virol. 2010, 84, 9497–9504. [Google Scholar]
- Clem, R.J. Baculoviruses and apoptosis: A diversity of genes and responses. Curr. Drug Targets 2007, 8, 1069–1074. [Google Scholar] [CrossRef]
- Bump, N.J.; Hackett, M.; Hugunin, M.; Seshagiri, S.; Brady, K.; Chen, P.; Ferenz, C.; Franklin, S.; Ghayur, T.; Li, P.; et al. Inhibition of ICE family proteases by baculovirus antiapoptotic protein p35. Science 1995, 269, 1885–1888. [Google Scholar]
- Guy, M.P.; Friesen, P.D. Reactive-site cleavage residues confer target specificity to baculovirus P49, a dimeric member of the P35 family of caspase inhibitors. J. Virol. 2008, 82, 7504–7514. [Google Scholar] [CrossRef]
- Schultz, K.L.W.; Friesen, P.D. Baculovirus DNA replication-specific expression factors trigger apoptosis and shutoff of host protein synthesis during infection. J. Virol. 2009, 83, 11123–11132. [Google Scholar] [CrossRef]
- Belyavskyi, M.; Braunagel, S.C.; Summers, M.D. The structural protein ODV-EC27 of Autographa californica nucleopolyhedrovirus is a multifunctional viral cyclin. Proc. Natl. Acad. Sci. USA. 1998, 95, 11205–11210. [Google Scholar] [CrossRef]
- Wu, W.; Passarelli, A.L. Autographa californica multiple nucleopolyhedrovirus Ac92 (ORF92, P33) is required for budded virus production and multiply enveloped occlusion-derived virus formation. J. Virol. 2010, 84, 12351–12361. [Google Scholar] [CrossRef]
- Dreschers, S.; Roncarati, R.; Knebel-Morsdorf, D. Actin rearrangement-inducing factor of baculoviruses is tyrosine phosphorylated and colocalizes to F-actin at the plasma membrane. J. Virol. 2001, 75, 3771–3778. [Google Scholar] [CrossRef]
- Marek, M.; Merten, O.-W.; Galibert, L.; Vlak, J.M.; van Oers, M.M. Baculovirus VP80 protein and the F-Actin cytoskeleton interact and connect the viral replication factory with the nuclear periphery. J. Virol. 2011, 85, 5350–5362. [Google Scholar] [CrossRef]
- Ohkawa, T.; Volkman, L.E.; Welch, M.D. Actin-based motility drives baculovirus transit to the nucleus and cell surface. J. Cell Biol. 2010, 190, 187–195. [Google Scholar] [CrossRef]
- Ohkawa, T.; Rowe, A.R.; Volkman, L.E. Identification of six Autographa californica multicapsid nucleopolyhedrovirus early genes that mediate nuclear localization of G-actin. J. Virol. 2002, 76, 12281–12289. [Google Scholar] [CrossRef]
- Li, K.; Wang, Y.; Bai, H.; Wang, Q.; Song, J.; Zhou, Y.; Wu, C.; Chen, X. The putative pocket protein binding site of Autographa californica nucleopolyhedrovirus BV/ODV-C42 is required for virus-induced nuclear actin polymerization. J. Virol. 2010, 84, 7857–7868. [Google Scholar]
- Goley, E.D.; Ohkawa, T.; Mancuso, J.; Woodruff, J.B.; D’Alessio, J.A.; Cande, W.Z.; Volkman, L.E.; Welch, M.D. Dynamic nuclear actin assembly by Arp2/3 complex and a baculovirus WASP-Like protein. Science 2006, 314, 464–467. [Google Scholar] [CrossRef]
- Fang, M.; Nie, Y.; Theilmann, D.A. AcMNPV EXON0 (AC141) which is required for the efficient egress of budded virus nucleocapsids interacts with beta-tubulin. Virology 2009, 385, 496–504. [Google Scholar] [CrossRef]
- Carpentier, D.C.; Griffiths, C.M.; King, L.A. The baculovirus P10 protein of Autographa californica nucleopolyhedrovirus forms two distinct cytoskeletal-like structures and associates with polyhedral occlusion bodies during infection. Virology 2008, 371, 278–291. [Google Scholar] [CrossRef]
- Braunagel, S.C.; Cox, V.; Summers, M.D. Baculovirus data suggest a common but multifaceted pathway for sorting proteins to the inner nuclear membrane. J. Virol. 2009, 83, 1280–1288. [Google Scholar] [CrossRef]
- Ge, J.; Wei, Z.; Huang, Y.; Yin, J.; Zhou, Z.; Zhong, J. AcMNPV ORF38 protein has the activity of ADP-ribose pyrophosphatase and is important for virus replication. Virology 2007, 361, 204–211. [Google Scholar] [CrossRef]
- Long, C.M.; Rohrmann, G.F.; Merrill, G.F. The conserved baculovirus protein p33 (Ac92) is a flavin adenine dinucleotide-linked sulfhydryl oxidase. Virology 2009, 388, 231–235. [Google Scholar] [CrossRef]
- Hakim, M.; Mandelbaum, A.; Fass, D. Structure of a baculovirus sulfhydryl oxidase, a highly divergent member of the erv flavoenzyme family. J. Virol. 2011, 85, 9406–9413. [Google Scholar] [CrossRef]
- Tomalski, M.D.; Eldridge, R.; Miller, L.K. A baculovirus homolog of a Cu/Zn superoxide dismutase gene. Virology 1991, 184, 149–161. [Google Scholar] [CrossRef]
- Wang, W.; Wu, J.; Wu, X. Deletion of superoxide dismutase gene of Bombyx mori nuclear polyhedrosis virus affects viral DNA replication. Int. J. Ind. Entomol. 2004, 9, 225–228. [Google Scholar]
- Kuzio, J.; Pearson, M.N.; Harwood, S.H.; Funk, C.J.; Evans, J.T.; Slavicek, J.M.; Rohrmann, G.F. Sequence and analysis of the genome of a baculovirus pathogenic for Lymantria dispar. Virology 1999, 253, 17–34. [Google Scholar] [CrossRef]
- Hughes, A.L.; Friedman, R. Genome-wide survey for genes horizontally transferred from cellular organisms to baculoviruses. Mol. Biol. Evol. 2003, 20, 979–987. [Google Scholar] [CrossRef]
- Vanarsdall, A.L.; Mikhailov, V.S.; Rohrmann, G.F. Baculovirus DNA replication and processing. Curr. Drug Targets 2007, 8, 1096–1102. [Google Scholar] [CrossRef]
- Baldo, A.M.; McClure, M.A. Evolution and horizontal transfer of dUTPase-encoding genes in viruses and their hosts. J. Virol. 1999, 73, 7710–7721. [Google Scholar]
- Landais, I.; Vincent, R.; Bouton, M.; Devauchelle, G.; Duonor-Cerutti, M.; Ogliastro, M. Functional analysis of evolutionary conserved clustering of bZIP binding sites in the baculovirus homologous regions (hrs) suggests a cooperativity between host and viral transcription factors. Virology 2006, 344, 421–431. [Google Scholar] [CrossRef]
- Matzat, L.H.; Berberoglu, S.; Levesque, L. Formation of a Tap/NXF1 homotypic complex is mediated through the amino-terminal domain of Tap and enhances interaction with nucleoporins. Mol. Biol. Cell 2008, 19, 327–338. [Google Scholar] [CrossRef]
- Thiem, S.M.; Chejanovsky, N. The role of baculovirus apoptotic suppressors in AcMNPV-mediated translation arrest in Ld652Y cells. Virology 2004, 319, 292–305. [Google Scholar] [CrossRef]
- Dever, T.E.; Sripriya, R.; McLachlin, J.R.; Lu, J.; Fabian, J.R.; Kimball, S.R.; Miller, L.K. Disruption of cellular translational control by a viral truncated eukaryotic translation initiation factor 2alpha kinase homolog. Proc. Natl. Acad. Sci. USA 1998, 95, 4164–4169. [Google Scholar]
- Li, Y.; Miller, L.K. Expression and functional analysis of a baculovirus gene encoding a truncated protein kinase homolog. Virology 1995, 206, 314–323. [Google Scholar] [CrossRef]
- Thiem, S.M.; Du, X.; Quentin, M.E.; Berner, M.M. Identification of baculovirus gene that promotes Autographa californica nuclear polyhedrosis virus replication in a nonpermissive insect cell line. J. Virol. 1996, 70, 2221–2229. [Google Scholar]
- Shirata, N.; Ikeda, M.; Kobayashi, M. Identification of a Hyphantria cunea nucleopolyhedrovirus (NPV) gene that is involved in global protein synthesis shutdown and restricted Bombyx mori NPV multiplication in a B. mori cell line. Virology 2010, 398, 149–157. [Google Scholar] [CrossRef]
- Imai, N.; Matsuda, N.; Tanaka, K.; Nakano, A.; Matsumoto, S.; Kang, W. Ubiquitin ligase activities of Bombyx mori nucleopolyhedrovirus RING finger proteins. J. Virol. 2003, 77, 923–930. [Google Scholar] [CrossRef]
- Katsuma, S.; Koyano, Y.; Kang, W.; Kokusho, R.; Kamita, S.G.; Shimada, T. The Baculovirus uses a captured host phosphatase to induce enhanced locomotory activity in host caterpillars. PLoS Pathog. 2012, 8, e1002644. [Google Scholar] [CrossRef]
- Van Houte, S.; Ros, V.I.D.; Mastenbroek, T.G.; Vendrig, N.J.; Hoover, K.; Spitzen, J.; van Oers, M.M. Protein Tyrosine phosphatase-induced hyperactivity is a conserved strategy of a subset of baculoVIruses to manipulate lepidopteran host behavior. PLoS One 2012, 7, e46933. [Google Scholar] [CrossRef]
- Means, J.C.; Passarelli, A.L. Viral fibroblast growth factor, matrix metalloproteases, and caspases are associated with enhancing systemic infection by baculoviruses. Proc. Natl. Acad. Sci. USA 2010, 107, 9825–9830. [Google Scholar] [CrossRef]
- Detvisitsakun, C.; Berretta, M.F.; Lehiy, C.; Passarelli, A.L. Stimulation of cell motility by a viral fibroblast growth factor homolog: Proposal for a role in viral pathogenesis. Virology 2005, 336, 308–317. [Google Scholar] [CrossRef]
- Hawtin, R.E.; Zarkowska, T.; Arnold, K.; Thomas, C.J.; Gooday, G.W.; King, L.A.; Kuzio, J.A.; Possee, R.D. Liquefaction of Autographa californica nucleopolyhedrovirus-infected insects is dependent on the integrity of virus-encoded chitinase and cathepsin genes. Virology 1997, 238, 243–253. [Google Scholar] [CrossRef]
- Hoover, K.; Grove, M.; Gardner, M.; Hughes, D.P.; McNeil, J.; Slavicek, J. A gene for an extended phenotype. Science 2011, 333. [Google Scholar] [CrossRef]
- O’Reilly, D.R.; Miller, L.K. A baculovirus blocks insect molting by producing ecdysteroid UDP-glucosyl transferase. Science 1989, 245, 1110–1112. [Google Scholar]
- Singh, C.P.; Singh, J.; Nagaraju, J. A baculovirus-encoded MicroRNA (miRNA) suppresses its host miRNA biogenesis by regulating the exportin-5 cofactor Ran. J. Virol. 2012, 86, 7867–7879. [Google Scholar] [CrossRef]
- Singha, J.; Singha, C.P.; Bhavania, A.; Nagaraju, J. Discovering microRNAs from Bombyx mori nucleopolyhedrosis virus. Virology 2010, 407, 120–128. [Google Scholar] [CrossRef]
- Moreno-Habel, D.A.; Biglang-awa, I.M.; Dulce, A.; Luu, D.D.; Garcia, P.; Weers, P.M.M.; Haas-Stapleton, E.J. Inactivation of the budded virus of Autographa californica M nucleopolyhedrovirus by gloverin. J. Invertebr. Pathol. 2012, 110, 92–101. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; He, H.-J.; Zhao, X.-F.; Wang, J.-X. Immune responses of Helicoverpa armigera to different kinds of pathogens. BMC Immunol. 2010, 11. [Google Scholar] [CrossRef]
- Vandergaast, R.; Schultz, K.L.W.; Cerio, R.J.; Friesen, P.D. Active depletion of host cell inhibitor-of-apoptosis IAP triggers apoptosis upon Baculovirus DNA replication. J. Virol. 2011, 85, 8348–8358. [Google Scholar] [CrossRef]
- Xiao, W.; Yang, Y.; Weng, Q.; Lin, T.; Yuan, M.; Yang, K.; Pang, Y. The role of the PI3K-Akt signal transduction pathway in Autographa californica multiple nucleopolyhedrovirus infection of Spodoptera frugiperda cells. Virology 2009, 391, 83–89. [Google Scholar] [CrossRef]
- Cooray, S. The pivotal role of phosphatidylinositol 3-kinase-Akt signal transduction in virus survival. J. Gen. Virol. 2004, 85, 1065–1076. [Google Scholar] [CrossRef]
- Katsuma, S.; Mita, K.; Shimada, T. ERK- and JNK-dependent signaling pathways contribute to Bombyx mori nucleopolyhedrovirus infection. J. Virol. 2007, 81, 13700–13709. [Google Scholar] [CrossRef]
- Broehan, G.; Kroeger, T.; Lorenzen, M.; Merzendorfer, H. Functional analysis of the ATP-binding cassette (ABC) transporter gene family of Tribolium castaneum. BMC Genomics 2013, 14, 1471–2164. [Google Scholar]
- Bernal, V.; Carinhas, N.; Yokomizo, A.Y.; Carrondo, M.J.T.; Alves, P.M. Cell density effect in the baculovirus-insect cells system: A quantitative analysis of energetic metabolism. Biotechnol. Bioeng. 2009, 104, 162–180. [Google Scholar] [CrossRef]
- Popham, H.; Sun, R.; Shelby, K.; Robertson, J. Iron levels change in larval Heliothis virescens tissues following baculovirus infection. Biol. Trace Elem. Res. 2012, 148, 356–362. [Google Scholar] [CrossRef]
- Mayer, M.P. Recruitment of Hsp70 chaperones: A crucial part of viral survival strategies. Rev. Physiol. Biochem. Pharmacol. 2005, 153, 1–46. [Google Scholar] [CrossRef]
- Aarti, I.; Rajesh, K.; Ramaiah, K.A. Phosphorylation of eIF2 alpha in Sf9 cells: A stress, survival and suicidal signal. Apoptosis 2010, 15, 679–692. [Google Scholar] [CrossRef]
- Ghosh, S.; Jain, A.; Mukherjee, B.; Habib, S.; Hasnain, S.E. The host factor polyhedrin promoter binding protein (PPBP) is involved in transcription from the baculovirus polyhedrin gene promoter. J. Virol. 1998, 72, 7484–7493. [Google Scholar]
- Lyupina, Y.V.; Abaturova, S.B.; Erokhov, P.A.; Orlova, O.V.; Beljelarskaya, S.N.; Mikhailov, V.S. Proteotoxic stress induced by Autographa californica nucleopolyhedrovirus infection of Spodoptera frugiperda Sf9 cells. Virology 2013, 436, 49–58. [Google Scholar] [CrossRef]
- Yu, X.; Zhou, Q.; Li, S.C.; Luo, Q.; Cai, Y.; Lin, W.C.; Chen, H.; Yang, Y.; Hu, S.; Yu, J. The silkworm (Bombyx mori) microRNAs and their expressions in multiple developmental stages. PLoS One 2008, 3, 0002997. [Google Scholar]
- Jayachandran, B.; Hussain, M.; Asgari, S. RNA Interference as a cellular defense mechanism against the DNA virus baculovirus. J. Virol. 2012, 86, 13729–13734. [Google Scholar] [CrossRef]
- Menzel, T.; Rohrmann, G. Diversity of errantivirus (retrovirus) sequences in two cell lines used for baculovirus expression, Spodoptera frugiperda and Trichoplusia ni. Virus Genes 2008, 36, 583–586. [Google Scholar] [CrossRef]
- Giri, L.; Feiss, M.G.; Bonning, B.C.; Murhammer, D.W. Production of baculovirus defective interfering particles during serial passage is delayed by removing transposon target sites in fp25k. J. Gen. Virol. 2012, 93, 389–399. [Google Scholar] [CrossRef]
- Fuller, D.N.; Raymer, D.M.; Kottadiel, V.I.; Rao, V.B.; Smith, D.E. Single phage T4 DNA packaging motors exhibit large force generation, high velocity, and dynamic variability. Proc. Natl. Acad. Sci. USA. 2007, 104, 16868–16873. [Google Scholar] [CrossRef]
- Nahlik, K.W.; Mleczko, A.K.; Gawlik, M.K.; Rokita, H.B. Modulation of GAPDH expression and cellular localization after vaccinia virus infection of human adherent monocytes. Acta Biochim. Pol. 2003, 50, 667–676. [Google Scholar]
- Cho, Y.S.; Lee, S.Y.; Kim, K.H.; Nam, Y.K. Differential modulations of two glyceraldehyde 3-phosphate dehydrogenase mRNAs in response to bacterial and viral challenges in a marine teleost Oplegnathus fasciatus (Perciformes). Fish Shellfish Immunol. 2008, 25, 472–476. [Google Scholar] [CrossRef]
- Scheper, G.C.; Vries, R.G.; Broere, M.; Usmany, M.; Voorma, H.O.; Vlak, J.M.; Thomas, A.A. Translational properties of the untranslated regions of the p10 messenger RNA of Autographa californica multicapsid nucleopolyhedrovirus. J. Gen. Virol. 1997, 78, 687–696. [Google Scholar]
- Van Oers, M.M.; van der Veken, L.T.J.N.; Vlak, J.M.; Thomas, A.A.M. Effect of baculovirus infection on the mRNA and protein levels of the Spodoptera frugiperda eukaryotic initiation factor 4E. Insect Mol. Biol. 2001, 10, 255–264. [Google Scholar] [CrossRef]
- Volkman, L.E. Baculovirus infectivity and the actin cytoskeleton. Curr. Drug Targets 2007, 8, 1075–1083. [Google Scholar] [CrossRef]
- Peng, K.; Wu, M.Z.; Deng, F.; Song, J.J.; Dong, C.S.; Wang, H.L.; Hu, Z.H. Identification of protein-protein interactions of the occlusion-derived virus-associated proteins of Helicoverpa armigera nucleopolyhedrovirus. J. Gen. Virol. 2010, 91, 659–670. [Google Scholar] [CrossRef]
- Braunagel, S.C.; Summers, M.D. Molecular biology of the baculovirus occlusion-derived virus envelope. Curr. Drug Targets 2007, 8, 1084–1095. [Google Scholar] [CrossRef]
- Vanarsdall, A.L.; Okano, K.; Rohrmann, G.F. Characterization of the replication of a baculovirus mutant lacking the DNA polymerase gene. Virology 2005, 331, 175–180. [Google Scholar] [CrossRef]
- Ahrens, C.H.; Rohrmann, G.F. The DNA polymerase and helicase genes of a baculovirus of Orgyia pseudosugata. J. Gen. Virol. 1996, 77, 825–837. [Google Scholar] [CrossRef]
- Braunagel, S.C.; Parr, R.; Belyavskyi, M.; Summers, M.D. Autographa californica nucleopolyhedrovirus infection results in Sf9 cell cycle arrest at G2/M phase. Virology 1998, 244, 195–211. [Google Scholar] [CrossRef]
- Weitzman, M.D.; Lilley, C.E.; Chaurushiya, M.S. Genomes in conflict: Maintaining genome integrity during virus infection. Annu. Rev. Microbiol. 2010, 64, 61–81. [Google Scholar] [CrossRef]
- Jin, J.; Guarino, L.A. 3'-End Formation of Baculovirus Late RNAs. J. Virol. 2000, 74, 8930–8937. [Google Scholar] [CrossRef]
- Ikeda, M.; Yamada, H.; Hamajima, R.; Kobayashi, M. Baculovirus genes modulating intracellular innate antiviral immunity of lepidopteran insect cells. Virology 2013, 435, 1–13. [Google Scholar] [CrossRef]
- Baluchamy, S.; Gopinathan, K.P. Characterization of a cyclin homolog from Bombyx mori nucleopolyhedrovirus. Virus Res. 2005, 108, 69–81. [Google Scholar] [CrossRef]
- Cartier, J.L.; Hershberger, P.A.; Friesen, P.D. Suppression of apoptosis in insect cells stably transfected with baculovirus p35: Dominant interference by N-terminal sequences p35(1–76). J. Virol. 1994, 68, 7728–7737. [Google Scholar]
- Kim, E.J.; Kramer, S.F.; Hebert, C.G.V.; James, J.; Bentley, W.E. Metabolic engineering of the baculovirusexpression system via inverse “Shotgun” genomic analysis and RNA interference (dsRNA) increases product yield and cell longevity. Biotechnol. Bioeng. 2007, 98, 645–654. [Google Scholar] [CrossRef]
- Terenius, O.; Papanicolaou, A.; Garbutt, J.S.; Eleftherianos, I.; Huvenne, H.; Kanginakudru, S.; Albrechtsen, M.; An, C.; Aymeric, J.-L.; Barthel, A.; et al. RNA interference in Lepidoptera: An overview of successful and unsuccessful studies and implications for experimental design. J. Insect Physiol. 2011, 57, 231–245. [Google Scholar] [CrossRef]
- Takasu, Y.; Kobayashi, I.; Beumer, K.; Uchino, K.; Sezutsu, H.; Sajwan, S.; Carroll, D.; Tamura, T.; Zurovec, M. Targeted mutagenesis in the silkworm Bombyx mori using zinc finger nuclease mRNA injection. Insect Biochem. Mol. Biol. 2010, 40, 759–765. [Google Scholar] [CrossRef]
- Ogay, I.D.; Lihoradova, O.A.; Azimova Sh, S.; Abdukarimov, A.A.; Slack, J.M.; Lynn, D.E. Transfection of insect cell lines using polyethylenimine. Cytotechnology 2006, 51, 89–98. [Google Scholar] [CrossRef]
- Lee, J.M.; Hull, J.J.; Kawai, T.; Tsuneizumi, K.; Kuriharaa, M.; Tanokura, M.; Nagata, K.; Nagasawa, H.; Matsumoto, S. Establishment of Sf9 transformants constitutively expressing PBAN receptor (PBANR) variants: Application to functional evaluation. Front. Endocrinol. 2012, 3. [Google Scholar] [CrossRef]
- Kost, T.A.; Condreay, J.P.; Jarvis, D.L. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat. Biotechnol. 2005, 23, 567–575. [Google Scholar] [CrossRef]
- Van Oers, M.M. Opportunities and challenges for the baculovirus expression system. J. Invertebr. Pathol. 2011, 107, S3–S15. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Nookaew, I.; Papini, M.; Pornputtpong, N.; Scalcinati, G.; Fagerberg, L.; Uhlén, M.; Nielsen, J. A comprehensive comparison of RNA-Seq-based transcriptome analysis from reads to differential gene expression and cross-comparison with microarrays: A case study in Saccharomyces cerevisiae. Nucleic Acids Res. 2012, 40, 10084–10097. [Google Scholar] [CrossRef]
- Kogenaru, S.; Yan, Q.; Guo, Y.; Wang, N. RNA-seq and microarray complement each other in transcriptome profiling. BMC Genomics 2012, 13. [Google Scholar] [CrossRef]
- Berkhout, B.; Coombs, K.M. Quantitative omics and its application to study virus-host interactions-a new frontier. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef]
- Popham, H.J.; Grasela, J.J.; Goodman, C.L.; McIntosh, A.H. Baculovirus infection influences host protein expression in two established insect cell lines. J. Insect Physiol. 2010, 56, 1237–1245. [Google Scholar] [CrossRef]
- Courtiade, J.; Muck, A.; Svatos, A.; Heckel, D.G.; Pauchet, Y. Comparative proteomic analysis of Helicoverpa armigera cells undergoing apoptosis. J. Proteome Res. 2011, 10, 2633–2642. [Google Scholar] [CrossRef]
- Chen, H.Q.; Yao, Q.; Bao, F.; Chen, K.P.; Liu, X.Y.; Li, J.; Wang, L. Comparative proteome analysis of silkworm in its susceptibility and resistance responses to Bombyx mori densonucleosis virus. Intervirology 2012, 55, 21–28. [Google Scholar] [CrossRef]
- Christians, U.; Klawitter, J.; Hornberger, A.; Klawitter, J. How unbiased is non-targeted metabolomics and is targeted pathway screening the solution? Curr. Pharm. Biotechnol. 2011, 12, 1053–1066. [Google Scholar] [CrossRef]
- Tran, T.T.B.; Dietmair, S.; Chan, L.C.L.; Huynh, H.T.; Nielsen, L.K.; Reid, S. Development of quenching and washing protocols for quantitative intracellular metabolite analysis of uninfected and baculovirus-infected insect cells. Methods 2012, 56, 396–407. [Google Scholar]
- Bernal, V.; Monteiro, F.; Carinhas, N.; Ambrósio, R.; Alves, P.M. An integrated analysis of enzyme activities, cofactor pools and metabolic fluxes in baculovirus-infected Spodoptera frugiperda Sf9 cells. J. Biotechnol. 2010, 150, 332–342. [Google Scholar]
- Bantscheff, M.; Lemeer, S.; Savitski, M.; Kuster, B. Quantitative mass spectrometry in proteomics: Critical review update from 2007 to the present. Anal. Bioanal. Chem. 2012, 404, 939–965. [Google Scholar] [CrossRef]
- Kafsack, B.F.C.; Llinás, M. Eating at the table of another: Metabolomics of host-parasite interactions. Cell Host Microbe 2010, 7, 90–99. [Google Scholar] [CrossRef]
- Ou, L.; Duan, D.; Wu, J.; Nice, E.; Huang, C. The application of high throughput siRNA screening technology to study host-pathogen interactions. Comb. Chem. High Throughput Screen. 2012, 15, 299–305. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nguyen, Q.; Nielsen, L.K.; Reid, S. Genome Scale Transcriptomics of Baculovirus-Insect Interactions. Viruses 2013, 5, 2721-2747. https://doi.org/10.3390/v5112721
Nguyen Q, Nielsen LK, Reid S. Genome Scale Transcriptomics of Baculovirus-Insect Interactions. Viruses. 2013; 5(11):2721-2747. https://doi.org/10.3390/v5112721
Chicago/Turabian StyleNguyen, Quan, Lars K. Nielsen, and Steven Reid. 2013. "Genome Scale Transcriptomics of Baculovirus-Insect Interactions" Viruses 5, no. 11: 2721-2747. https://doi.org/10.3390/v5112721
APA StyleNguyen, Q., Nielsen, L. K., & Reid, S. (2013). Genome Scale Transcriptomics of Baculovirus-Insect Interactions. Viruses, 5(11), 2721-2747. https://doi.org/10.3390/v5112721