Nuclear Actin and Lamins in Viral Infections

Abstract
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Viruses and Nuclear Actin
2.1. Herpesviruses
2.1.1. Nuclear Actin in the Assembly and Transport of Viral Capsids
2.1.2. Nuclear Actin and Morphological Changes of Infected Nuclei
2.1.3. Monomeric versus Polymeric Nuclear Actin
2.2. Baculoviruses
2.2.1. Nuclear Actin Filaments and Nucleocapsid Morphogenesis
2.2.2. Actin Relocalization to the Nucleus
2.2.3. Mechanism of Nuclear Actin Polymerization
2.2.4. F-Actin and the Nuclear Egress of AcMNPV
2.3. Retroviruses
3. Viruses and Nuclear Lamins
3.1. Herpesviruses
3.1.1. HSV-1
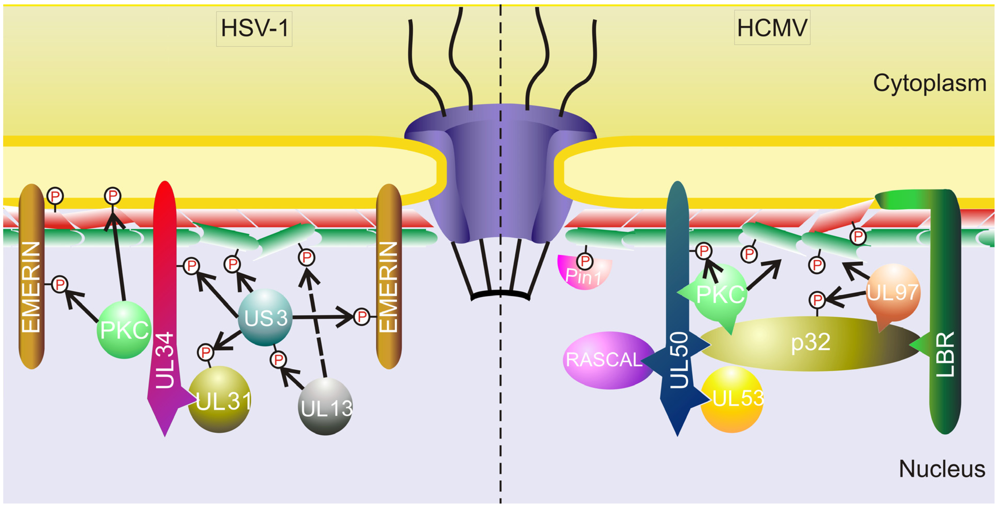
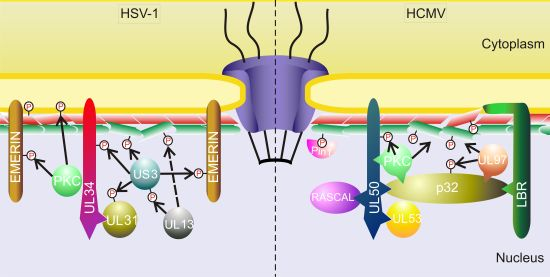
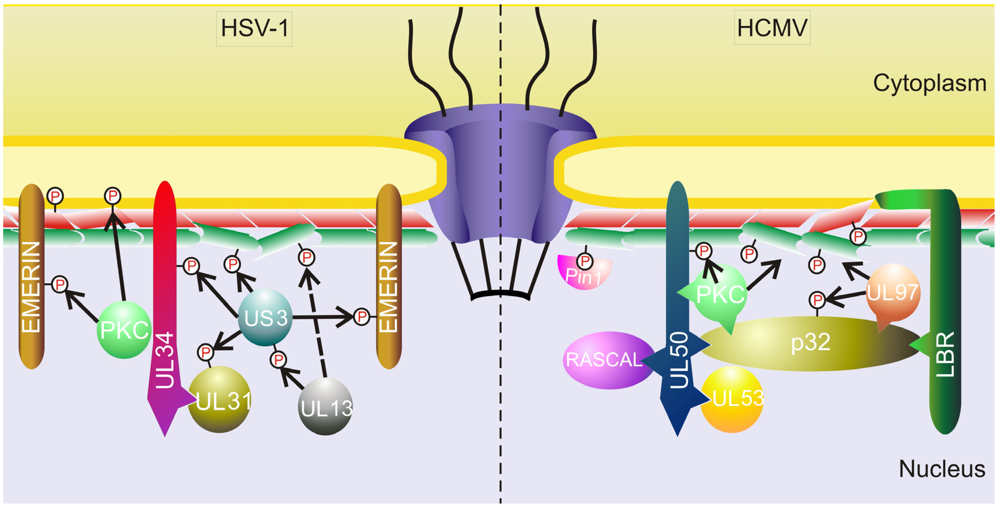
3.1.2. Other Herpesviruses
3.2. Other Viruses
3.2.1. Retroviruses
3.2.2. Polyomaviruses
3.2.3. Parvoviruses
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Radtke, K.; Dohner, K.; Sodeik, B. Viral interactions with the cytoskeleton: A hitchhiker’s guide to the cell. Cell. Microbiol. 2006, 8, 387–400. [Google Scholar]
- Clark, T.G.; Rosenbaum, J.L. An actin filament matrix in hand-isolated nuclei of x. Laevis oocytes. Cell 1979, 18, 1101–1108. [Google Scholar] [CrossRef]
- Scheer, U.; Hinssen, H.; Franke, W.W.; Jockusch, B.M. Microinjection of actin-binding proteins and actin antibodies demonstrates involvement of nuclear actin in transcription of lampbrush chromosomes. Cell 1984, 39, 111–122. [Google Scholar]
- Miralles, F.; Visa, N. Actin in transcription and transcription regulation. Curr. Opin. Cell Biol. 2006, 18, 261–266. [Google Scholar]
- Zheng, B.; Han, M.; Bernier, M.; Wen, J.K. Nuclear actin and actin-binding proteins in the regulation of transcription and gene expression. FEBS J. 2009, 276, 2669–2685. [Google Scholar]
- Olave, I.A.; Reck-Peterson, S.L.; Crabtree, G.R. Nuclear actin and actin-related proteins in chromatin remodeling. Annu. Rev. Biochem. 2002, 71, 755–781. [Google Scholar]
- Hofmann, W.A.; Stojiljkovic, L.; Fuchsova, B.; Vargas, G.M.; Mavrommatis, E.; Philimonenko, V.; Kysela, K.; Goodrich, J.A.; Lessard, J.L.; Hope, T.J.; et al. Actin is part of pre-initiation complexes and is necessary for transcription by rna polymerase ii. Nat. Cell Biol. 2004, 6, 1094–1101. [Google Scholar] [CrossRef]
- Hu, P.; Wu, S.; Hernandez, N. A role for beta-actin in rna polymerase iii transcription. Genes Dev. 2004, 18, 3010–3015. [Google Scholar]
- Philimonenko, V.V.; Zhao, J.; Iben, S.; Dingová, H.; Kyselá, K.; Kahle, M.; Zentgraf, H.; Hofmann, W.A.; de Lanerolle, P.; Hozák, P.; et al. Nuclear actin and myosin i are required for rna polymerase i transcription. Nat. Cell Biol. 2004, 6, 1165–1172. [Google Scholar] [CrossRef]
- Percipalle, P.; Zhao, J.; Pope, B.; Weeds, A.; Lindberg, U.; Daneholt, B. Actin bound to the heterogeneous nuclear ribonucleoprotein hrp36 is associated with balbiani ring mrna from the gene to polysomes. J. Cell Biol. 2001, 153, 229–236. [Google Scholar]
- Percipalle, P.; Jonsson, A.; Nashchekin, D.; Karlsson, C.; Bergman, T.; Guialis, A.; Daneholt, B. Nuclear actin is associated with a specific subset of hnrnp a/b-type proteins. Nucleic Acids Res. 2002, 30, 1725–1734. [Google Scholar]
- Kukalev, A.; Nord, Y.; Palmberg, C.; Bergman, T.; Percipalle, P. Actin and hnrnp u cooperate for productive transcription by rna polymerase ii. Nat. Struct. Mol. Biol. 2005, 12, 238–244. [Google Scholar]
- Sjölinder, M.; Björk, P.; Söderberg, E.; Sabri, N.; Farrants, A.K.; Visa, N. The growing pre-mrna recruits actin and chromatin-modifying factors to transcriptionally active genes. Genes Dev. 2005, 19, 1871–1884. [Google Scholar]
- Zhao, K.; Wang, W.; Rando, O.J.; Xue, Y.; Swiderek, K.; Kuo, A.; Crabtree, G.R. Rapid and phosphoinositol-dependent binding of the swi/snf-like baf complex to chromatin after t lymphocyte receptor signaling. Cell 1998, 95, 625–636. [Google Scholar]
- Nowak, G.; Pestic-Dragovich, L.; Hozák, P.; Philimonenko, A.; Simerly, C.; Schatten, G.; de Lanerolle, P. Evidence for the presence of myosin i in the nucleus. J. Biol. Chem. 1997, 272, 17176–17181. [Google Scholar]
- Pestic-Dragovich, L.; Stojiljkovic, L.; Philimonenko, A.A.; Nowak, G.; Ke, Y.; Settlage, R.E.; Shabanowitz, J.; Hunt, D.F.; Hozak, P.; de Lanerolle, P. A myosin i isoform in the nucleus. Science 2000, 290, 337–341. [Google Scholar]
- Hofmann, W.A.; Vargas, G.M.; Ramchandran, R.; Stojiljkovic, L.; Goodrich, J.A.; de Lanerolle, P. Nuclear myosin i is necessary for the formation of the first phosphodiester bond during transcription initiation by rna polymerase ii. J. Cell Biochem. 2006, 99, 1001–1009. [Google Scholar]
- Percipalle, P.; Fomproix, N.; Cavellán, E.; Voit, R.; Reimer, G.; Krüger, T.; Thyberg, J.; Scheer, U.; Grummt, I.; Farrants, A.K. The chromatin remodelling complex wstf-snf2h interacts with nuclear myosin 1 and has a role in rna polymerase i transcription. EMBO Rep. 2006, 7, 525–530. [Google Scholar]
- Philimonenko, V.V.; Janacek, J.; Harata, M.; Hozak, P. Transcription-dependent rearrangements of actin and nuclear myosin i in the nucleolus. Histochem. Cell Biol. 2010, 134, 243–249. [Google Scholar]
- Fomproix, N.; Percipalle, P. An actin-myosin complex on actively transcribing genes. Exp. Cell Res. 2004, 294, 140–148. [Google Scholar]
- Kyselá, K.; Philimonenko, A.A.; Philimonenko, V.V.; Janácek, J.; Kahle, M.; Hozák, P. Nuclear distribution of actin and myosin i depends on transcriptional activity of the cell. Histochem. Cell Biol. 2005, 124, 347–358. [Google Scholar]
- Pederson, T.; Aebi, U. Actin in the nucleus: What form and what for? J. Struct. Biol. 2002, 140, 3–9. [Google Scholar] [CrossRef]
- Bettinger, B.T.; Gilbert, D.M.; Amberg, D.C. Actin up in the nucleus. Nat. Rev. Mol. Cell Biol. 2004, 5, 410–415. [Google Scholar]
- Gonsior, S.M.; Platz, S.; Buchmeier, S.; Scheer, U.; Jockusch, B.M.; Hinssen, H. Conformational difference between nuclear and cytoplasmic actin as detected by a monoclonal antibody. J. Cell Sci. 1999, 112, 797–809. [Google Scholar]
- Schoenenberger, C.A.; Buchmeier, S.; Boerries, M.; Sütterlin, R.; Aebi, U.; Jockusch, B.M. Conformation-specific antibodies reveal distinct actin structures in the nucleus and the cytoplasm. J. Struct. Biol. 2005, 152, 157–168. [Google Scholar]
- McDonald, D.; Carrero, G.; Andrin, C.; de Vries, G.; Hendzel, M.J. Nucleoplasmic beta-actin exists in a dynamic equilibrium between low-mobility polymeric species and rapidly diffusing populations. J. Cell Biol. 2006, 172, 541–552. [Google Scholar]
- Kiseleva, E.; Drummond, S.P.; Goldberg, M.W.; Rutherford, S.A.; Allen, T.D.; Wilson, K.L. Actin- and protein-4.1-containing filaments link nuclear pore complexes to subnuclear organelles in xenopus oocyte nuclei. J. Cell Sci. 2004, 117, 2481–2490. [Google Scholar] [CrossRef]
- Hofmann, W.; Reichart, B.; Ewald, A.; Müller, E.; Schmitt, I.; Stauber, R.H.; Lottspeich, F.; Jockusch, B.M.; Scheer, U.; Hauber, J.; et al. Cofactor requirements for nuclear export of rev response element (rre)- and constitutive transport element (cte)-containing retroviral rnas. An unexpected role for actin. J. Cell Biol. 2001, 152, 895–910. [Google Scholar] [CrossRef]
- Kimura, T.; Hashimoto, I.; Yamamoto, A.; Nishikawa, M.; Fujisawa, J.I. Rev-dependent association of the intron-containing hiv-1 gag mrna with the nuclear actin bundles and the inhibition of its nucleocytoplasmic transport by latrunculin-b. Genes Cells 2000, 5, 289–307. [Google Scholar]
- Castano, E.; Philimonenko, V.V.; Kahle, M.; Fukalová, J.; Kalendová, A.; Yildirim, S.; Dzijak, R.; Dingová-Krásna, H.; Hozák, P. Actin complexes in the cell nucleus: New stones in an old field. Histochem. Cell Biol. 2010, 133, 607–626. [Google Scholar]
- Chen, M.; Shen, X. Nuclear actin and actin-related proteins in chromatin dynamics. Curr. Opin. Cell Biol. 2007, 19, 326–330. [Google Scholar]
- Yahara, I.; Aizawa, H.; Moriyama, K.; Iida, K.; Yonezawa, N.; Nishida, E.; Hatanaka, H.; Inagaki, F. A role of cofilin/destrin in reorganization of actin cytoskeleton in response to stresses and cell stimuli. Cell Struct. Funct. 1996, 21, 421–424. [Google Scholar]
- van Hal, S.J.; Dwyer, D.E. Herpes Simplex: Viruses and Infections. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2001. [Google Scholar]
- Grünewald, K.; Desai, P.; Winkler, D.C.; Heymann, J.B.; Belnap, D.M.; Baumeister, W.; Steven, A.C. Three-dimensional structure of herpes simplex virus from cryo-electron tomography. Science 2003, 302, 1396–1398. [Google Scholar]
- Randall, R.E.; Dinwoodie, N. Intranuclear localization of herpes simplex virus immediate-early and delayed-early proteins: Evidence that icp 4 is associated with progeny virus DNA. J. Gen. Virol. 1986, 67, 2163–2177. [Google Scholar]
- Knipe, D.M.; Senechek, D.; Rice, S.A.; Smith, J.L. Stages in the nuclear association of the herpes simplex virus transcriptional activator protein icp4. J. Virol. 1987, 61, 276–284. [Google Scholar]
- Phelan, A.; Dunlop, J.; Patel, A.H.; Stow, N.D.; Clements, J.B. Nuclear sites of herpes simplex virus type 1 DNA replication and transcription colocalize at early times postinfection and are largely distinct from rna processing factors. J. Virol. 1997, 71, 1124–1132. [Google Scholar]
- Ward, P.L.; Ogle, W.O.; Roizman, B. Assemblons: Nuclear structures defined by aggregation of immature capsids and some tegument proteins of herpes simplex virus 1. J. Virol. 1996, 70, 4623–4631. [Google Scholar]
- de Bruyn Kops, A.; Uprichard, S.L.; Chen, M.; Knipe, D.M. Comparison of the intranuclear distributions of herpes simplex virus proteins involved in various viral functions. Virology 1998, 252, 162–178. [Google Scholar]
- Leopardi, R.; Ward, P.L.; Ogle, W.O.; Roizman, B. Association of herpes simplex virus regulatory protein icp22 with transcriptional complexes containing eap, icp4, rna polymerase ii, and viral DNA requires posttranslational modification by the u(l)13 proteinkinae. J. Virol. 1997, 71, 1133–1139. [Google Scholar]
- Quinlan, M.P.; Chen, L.B.; Knipe, D.M. The intranuclear location of a herpes simplex virus DNA-binding protein is determined by the status of viral DNA replication. Cell 1984, 36, 857–868. [Google Scholar]
- de Bruyn Kops, A.; Knipe, D.M. Preexisting nuclear architecture defines the intranuclear location of herpesvirus DNA replication structures. J. Virol. 1994, 68, 3512–3526. [Google Scholar]
- Taylor, T.J.; McNamee, E.E.; Day, C.; Knipe, D.M. Herpes simplex virus replication compartments can form by coalescence of smaller compartments. Virology 2003, 309, 232–247. [Google Scholar]
- Monier, K.; Armas, J.C.; Etteldorf, S.; Ghazal, P.; Sullivan, K.F. Annexation of the interchromosomal space during viral infection. Nat. Cell Biol. 2000, 2, 661–665. [Google Scholar]
- Scott, E.S.; O’Hare, P. Fate of the inner nuclear membrane protein lamin b receptor and nuclear lamins in herpes simplex virus type 1 infection. J. Virol. 2001, 75, 8818–8830. [Google Scholar]
- Simpson-Holley, M.; Colgrove, R.C.; Nalepa, G.; Harper, J.W.; Knipe, D.M. Identification and functional evaluation of cellular and viral factors involved in the alteration of nuclear architecture during herpes simplex virus 1 infection. J. Virol. 2005, 79, 12840–12851. [Google Scholar]
- Simpson-Holley, M.; Baines, J.; Roller, R.; Knipe, D.M. Herpes simplex virus 1 u(l)31 and u(l)34 gene products promote the late maturation of viral replication compartments to the nuclear periphery. J. Virol. 2004, 78, 5591–5600. [Google Scholar]
- Feierbach, B.; Piccinotti, S.; Bisher, M.; Denk, W.; Enquist, L.W. Alpha-herpesvirus infection induces the formation of nuclear actin filaments. PLoS Pathog. 2006, 2, e85. [Google Scholar]
- Ecob-Johnston, M.S.; Whetsell, W.O. Host-cell response to herpes virus infection in central and peripheral nervous tissue in vitro. J. Gen. Virol. 1979, 44, 747–757. [Google Scholar]
- Forest, T.; Barnard, S.; Baines, J.D. Active intranuclear movement of herpesvirus capsids. Nat. Cell Biol. 2005, 7, 429–431. [Google Scholar]
- Nalepa, G.; Harper, J.W. Visualization of a highly organized intranuclear network of filaments in living mammalian cells. Cell Motil. Cytoskelet. 2004, 59, 94–108. [Google Scholar]
- Wong, M.L.; Chen, C.H. Evidence for the internal location of actin in the pseudorabies virion. Virus Res. 1998, 56, 191–197. [Google Scholar]
- del Rio, T.; DeCoste, C.J.; Enquist, L.W. Actin is a component of the compensation mechanism in pseudorabies virus virions lacking the major tegument protein vp22. J. Virol. 2005, 79, 8614–8619. [Google Scholar]
- Varnum, S.M.; Streblow, D.N.; Monroe, M.E.; Smith, P.; Auberry, K.J.; Pasa-Tolic, L.; Wang, D.; Camp, D.G.; Rodland, K.; Wiley, S.; et al. Identification of proteins in human cytomegalovirus (hcmv) particles: The hcmv proteome. J. Virol. 2004, 78, 10960–10966. [Google Scholar]
- Kattenhorn, L.M.; Mills, R.; Wagner, M.; Lomsadze, A.; Makeev, V.; Borodovsky, M.; Ploegh, H.L.; Kessler, B.M. Identification of proteins associated with murine cytomegalovirus virions. J. Virol. 2004, 78, 11187–11197. [Google Scholar]
- Bechtel, J.T.; Winant, R.C.; Ganem, D. Host and viral proteins in the virion of kaposi’s sarcoma-associated herpesvirus. J. Virol. 2005, 79, 4952–4964. [Google Scholar]
- Zhu, F.X.; Chong, J.M.; Wu, L.; Yuan, Y. Virion proteins of kaposi’s sarcoma-associated herpesvirus. J. Virol. 2005, 79, 800–811. [Google Scholar]
- Baines, J.D.; Hsieh, C.E.; Wills, E.; Mannella, C.; Marko, M. Electron tomography of nascent herpes simplex virus virions. J. Virol. 2007, 81, 2726–2735. [Google Scholar]
- Guarino, L. Baculoviruses. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2001. [Google Scholar]
- Nagamine, T.; Kawasaki, Y.; Abe, A.; Matsumoto, S. Nuclear marginalization of host cell chromatin associated with expansion of two discrete virus-induced subnuclear compartments during baculovirus infection. J. Virol. 2008, 82, 6409–6418. [Google Scholar]
- Volkman, L.E.; Goldsmith, P.A.; Hess, R.T. Evidence for microfilament involvement in budded autographa californica nuclear polyhedrosis virus production. Virology 1987, 156, 32–39. [Google Scholar]
- Volkman, L.E. Autographa californica mnpv nucleocapsid assembly: Inhibition by cytochalasin d. Virology 1988, 163, 547–553. [Google Scholar]
- Hess, R.T.; Goldsmith, P.A.; Volkman, L.E. Effect of cytochalasin d on cell morphology and acmnpv replication in a spodoptera frugiperda cell line. J. Invertebr. Pathol. 1989, 53, 169–182. [Google Scholar]
- Volkman, L.E.; Talhouk, S.N.; Oppenheimer, D.I.; Charlton, C.A. Nuclear f-actin - a functional component of baculovirus-infected lepidopteran cells. J. Cell Sci. 1992, 103, 15–22. [Google Scholar]
- Charlton, C.A.; Volkman, L.E. Sequential rearrangement and nuclear polymerization of actin in baculovirus-infected spodoptera frugiperda cells. J. Virol. 1991, 65, 1219–1227. [Google Scholar]
- Ohkawa, T.; Volkman, L.E. Nuclear f-actin is required for acmnpv nucleocapsid morphogenesis. Virology 1999, 264, 1–4. [Google Scholar]
- Kasman, L.M.; Volkman, L.E. Filamentous actin is required for lepidopteran nucleopolyhedrovirus progeny production. J. Gen. Virol. 2000, 81, 1881–1888. [Google Scholar]
- Ohkawa, T.; Rowe, A.R.; Volkman, L.E. Identification of six autographa californica multicapsid nucleopolyhedrovirus early genes that mediate nuclear localization of g-actin. J. Virol. 2002, 76, 12281–12289. [Google Scholar]
- Lanier, L.M.; Volkman, L.E. Actin binding and nucleation by autographa california m nucleopolyhedrovirus. Virology 1998, 243, 167–177. [Google Scholar]
- Charlton, C.A.; Volkman, L.E. Penetration of autographa californica nuclear polyhedrosis virus nucleocapsids into iplb sf 21 cells induces actin cable formation. Virology 1993, 197, 245–254. [Google Scholar]
- Goley, E.D.; Ohkawa, T.; Mancuso, J.; Woodruff, J.B.; D’Alessio, J.A.; Cande, W.Z.; Volkman, L.E.; Welch, M.D. Dynamic nuclear actin assembly by arp2/3 complex and a baculovirus wasp-like protein. Science 2006, 314, 464–467. [Google Scholar]
- Machesky, L.M.; Insall, R.H.; Volkman, L.E. Wasp homology sequences in baculoviruses. Trends Cell Biol. 2001, 11, 286–287. [Google Scholar]
- Wang, Y.; Wang, Q.; Liang, C.; Song, J.; Li, N.; Shi, H.; Chen, X. Autographa californica multiple nucleopolyhedrovirus nucleocapsid protein bv/odv-c42 mediates the nuclear entry of p78/83. J. Virol. 2008, 82, 4554–4561. [Google Scholar]
- Li, K.; Wang, Y.; Bai, H.; Wang, Q.; Song, J.; Zhou, Y.; Wu, C.; Chen, X. The putative pocket protein binding site of autographa californica nucleopolyhedrovirus bv/odv-c42 is required for virus-induced nuclear actin polymerization. J. Virol. 2010, 84, 7857–7868. [Google Scholar]
- Wang, Q.; Liang, C.; Song, J.; Chen, X. Ha2 from the helicoverpa armigera nucleopolyhedrovirus: A wasp-related protein that activates arp2/3-induced actin filament formation. Virus Res. 2007, 127, 81–87. [Google Scholar]
- Marek, M.; Merten, O.W.; Galibert, L.; Vlak, J.M.; van Oers, M.M. Baculovirus vp80 protein and the f-actin cytoskeleton interact connecting the viral replication factory with the nuclear periphery. J. Virol. 2011, 85, 5350–5362. [Google Scholar]
- Marek, M.; van Oers, M.M.; Devaraj, F.F.; Vlak, J.M.; Merten, O.W. Engineering of baculovirus vectors for the manufacture of virion-free biopharmaceuticals. Biotechnol. Bioeng. 2011, 108, 1056–1067. [Google Scholar]
- Cullen, B.R. Nuclear mrna export: Insights from virology. Trends Biochem. Sci. 2003, 28, 419–424. [Google Scholar]
- Aebi, U.; Cohn, J.; Buhle, L.; Gerace, L. The nuclear lamina is a meshwork of intermediate-type filaments. Nature 1986, 323, 560–564. [Google Scholar]
- Goldberg, M.W.; Huttenlauch, I.; Hutchison, C.J.; Stick, R. Filaments made from a- and b-type lamins differ in structure and organization. J. Cell Sci. 2008, 121, 215–225. [Google Scholar]
- Bridger, J.M.; Kill, I.R.; O’Farrell, M.; Hutchison, C.J. Internal lamin structures within g1 nuclei of human dermal fibroblasts. J. Cell Sci. 1993, 104, 297–306. [Google Scholar]
- Goldman, A.E.; Moir, R.D.; Montag-Lowy, M.; Stewart, M.; Goldman, R.D. Pathway of incorporation of microinjected lamin a into the nuclear envelope. J. Cell Biol. 1992, 119, 725–735. [Google Scholar]
- Moir, R.D.; Montag-Lowy, M.; Goldman, R.D. Dynamic properties of nuclear lamins: Lamin b is associated with sites of DNA replication. J. Cell Biol. 1994, 125, 1201–1212. [Google Scholar]
- Hozák, P.; Sasseville, A.M.; Raymond, Y.; Cook, P.R. Lamin proteins form an internal nucleoskeleton as well as a peripheral lamina in human cells. J. Cell Sci. 1995, 108, 635–644. [Google Scholar]
- Barboro, P.; D’Arrigo, C.; Diaspro, A.; Mormino, M.; Alberti, I.; Parodi, S.; Patrone, E.; Balbi, C. Unraveling the organization of the internal nuclear matrix: Rna-dependent anchoring of numa to a lamin scaffold. Exp. Cell Res. 2002, 279, 202–218. [Google Scholar]
- Barboro, P.; D’Arrigo, C.; Mormino, M.; Coradeghini, R.; Parodi, S.; Patrone, E.; Balbi, C. An intranuclear frame for chromatin compartmentalization and higher-order folding. J. Cell. Biochem. 2003, 88, 113–120. [Google Scholar]
- Neri, L.M.; Raymond, Y.; Giordano, A.; Capitani, S.; Martelli, A.M. Lamin a is part of the internal nucleoskeleton of human erythroleukemia cells. J. Cell. Physiol. 1999, 178, 284–295. [Google Scholar]
- Prokocimer, M.; Davidovich, M.; Nissim-Rafinia, M.; Wiesel-Motiuk, N.; Bar, D.Z.; Barkan, R.; Meshorer, E.; Gruenbaum, Y. Nuclear lamins: Key regulators of nuclear structure and activities. J. Cell. Mol. Med. 2009, 13, 1059–1085. [Google Scholar]
- Worman, H.J.; Bonne, G. Laminopathies”: A wide spectrum of human diseases. Exp. Cell Res. 2007, 313, 2121–2133. [Google Scholar]
- Panté, N.; Kann, M. Nuclear pore complex is able to transport macromolecules with diameters of about 39 nm. Mol. Biol. Cell 2002, 13, 425–434. [Google Scholar]
- Skepper, J.N.; Whiteley, A.; Browne, H.; Minson, A. Herpes simplex virus nucleocapsids mature to progeny virions by an envelopment --> deenvelopment --> reenvelopment pathway. J. Virol. 2001, 75, 5697–5702. [Google Scholar]
- Nagel, C.H.; Döhner, K.; Fathollahy, M.; Strive, T.; Borst, E.M.; Messerle, M.; Sodeik, B. Nuclear egress and envelopment of herpes simplex virus capsids analyzed with dual-color fluorescence hsv1(17+). J. Virol. 2008, 82, 3109–3124. [Google Scholar]
- Lee, C.P.; Chen, M.R. Escape of herpesviruses from the nucleus. Rev. Med. Virol. 2010, 20, 214–230. [Google Scholar]
- Roller, R.J.; Zhou, Y.; Schnetzer, R.; Ferguson, J.; DeSalvo, D. Herpes simplex virus type 1 u(l)34 gene product is required for viral envelopment. J. Virol. 2000, 74, 117–129. [Google Scholar]
- Chang, Y.E.; Roizman, B. The product of the ul31 gene of herpes simplex virus 1 is a nuclear phosphoprotein which partitions with the nuclear matrix. J. Virol. 1993, 67, 6348–6356. [Google Scholar]
- Reynolds, A.E.; Ryckman, B.J.; Baines, J.D.; Zhou, Y.; Liang, L.; Roller, R.J. U(l)31 and u(l)34 proteins of herpes simplex virus type 1 form a complex that accumulates at the nuclear rim and is required for envelopment of nucleocapsids. J. Virol. 2001, 75, 8803–8817. [Google Scholar]
- Reynolds, A.E.; Wills, E.G.; Roller, R.J.; Ryckman, B.J.; Baines, J.D. Ultrastructural localization of the herpes simplex virus type 1 ul31, ul34, and us3 proteins suggests specific roles in primary envelopment and egress of nucleocapsids. J. Virol. 2002, 76, 8939–8952. [Google Scholar]
- Liang, L.; Baines, J.D. Identification of an essential domain in the herpes simplex virus 1 ul34 protein that is necessary and sufficient to interact with ul31 protein. J. Virol. 2005, 79, 3797–3806. [Google Scholar]
- Mou, F.; Wills, E.G.; Park, R.; Baines, J.D. Effects of lamin a/c, lamin b1, and viral us3 kinase activity on viral infectivity, virion egress, and the targeting of herpes simplex virus u(l)34-encoded protein to the inner nuclear membrane. J. Virol. 2008, 82, 8094–8104. [Google Scholar] [CrossRef]
- Bjerke, S.L.; Roller, R.J. Roles for herpes simplex virus type 1 ul34 and us3 proteins in disrupting the nuclear lamina during herpes simplex virus type 1 egress. Virology 2006, 347, 261–276. [Google Scholar]
- Reynolds, A.E.; Liang, L.; Baines, J.D. Conformational changes in the nuclear lamina induced by herpes simplex virus type 1 require genes u(l)31 and u(l)34. J. Virol. 2004, 78, 5564–5575. [Google Scholar]
- Ryckman, B.J.; Roller, R.J. Herpes simplex virus type 1 primary envelopment: Ul34 protein modification and the us3-ul34 catalytic relationship. J. Virol. 2004, 78, 399–412. [Google Scholar]
- Mou, F.; Wills, E.; Baines, J.D. Phosphorylation of the u(l)31 protein of herpes simplex virus 1 by the u(s)3-encoded kinase regulates localization of the nuclear envelopment complex and egress of nucleocapsids. J. Virol. 2009, 83, 5181–5191. [Google Scholar]
- Mou, F.; Forest, T.; Baines, J.D. Us3 of herpes simplex virus type 1 encodes a promiscuous protein kinase that phosphorylates and alters localization of lamin a/c in infected cells. J. Virol. 2007, 81, 6459–6470. [Google Scholar]
- Morris, J.B.; Hofemeister, H.; O’Hare, P. Herpes simplex virus infection induces phosphorylationand delocalization of emerin, a key inner nuclear membrane protein. J. Virol. 2007, 81, 4429–4437. [Google Scholar]
- Leach, N.; Bjerke, S.L.; Christensen, D.K.; Bouchard, J.M.; Mou, F.; Park, R.; Baines, J.; Haraguchi, T.; Roller, R.J. Emerin is hyperphosphorylated and redistributed in herpes simplex virus type 1-infected cells in a manner dependent on both ul34 and us3. J. Virol. 2007, 81, 10792–10803. [Google Scholar]
- Leach, N.R.; Roller, R.J. Significance of host cell kinases in herpes simplex virus type 1 egress and lamin-associated protein disassembly from the nuclear lamina. Virology 2010, 406, 127–137. [Google Scholar]
- Kato, A.; Yamamoto, M.; Ohno, T.; Tanaka, M.; Sata, T.; Nishiyama, Y.; Kawaguchi, Y. Herpes simplex virus 1-encoded protein kinase ul13 phosphorylates viral us3 protein kinase and regulates nuclear localization of viral envelopment factors ul34 and ul31. J. Virol. 2006, 80, 1476–1486. [Google Scholar]
- Cano-Monreal, G.L.; Wylie, K.M.; Cao, F.; Tavis, J.E.; Morrison, L.A. Herpes simplex virus 2 ul13 protein kinase disrupts nuclear lamins. Virology 2009, 392, 137–147. [Google Scholar]
- Park, R.; Baines, J.D. Herpes simplex virus type 1 infection induces activation and recruitment of protein kinase c to the nuclear membrane and increased phosphorylation of lamin b. J. Virol. 2006, 80, 494–504. [Google Scholar]
- Shiba, C.; Daikoku, T.; Goshima, F.; Takakuwa, H.; Yamauchi, Y.; Koiwai, O.; Nishiyama, Y. The ul34 gene product of herpes simplex virus type 2 is a tail-anchored type ii membrane protein that is significant for virus envelopment. J. Gen. Virol. 2000, 81, 2397–2405. [Google Scholar]
- Yamauchi, Y.; Shiba, C.; Goshima, F.; Nawa, A.; Murata, T.; Nishiyama, Y. Herpes simplex virus type 2 ul34 protein requires ul31 protein for its relocation to the internal nuclear membrane in transfected cells. J. Gen. Virol. 2001, 82, 1423–1428. [Google Scholar]
- Klupp, B.G.; Granzow, H.; Mettenleiter, T.C. Primary envelopment of pseudorabies virus at the nuclear membrane requires the ul34 gene product. J. Virol. 2000, 74, 10063–10073. [Google Scholar]
- Fuchs, W.; Klupp, B.G.; Granzow, H.; Osterrieder, N.; Mettenleiter, T.C. The interacting ul31 and ul34 gene products of pseudorabies virus are involved in egress from the host-cell nucleus and represent components of primary enveloped but not mature virions. J. Virol. 2002, 76, 364–378. [Google Scholar]
- Camozzi, D.; Pignatelli, S.; Valvo, C.; Lattanzi, G.; Capanni, C.; Dal Monte, P.; Landini, M.P. Remodelling of the nuclear lamina during human cytomegalovirus infection: Role of the viral proteins pul50 and pul53. J. Gen. Virol. 2008, 89, 731–740. [Google Scholar]
- Muranyi, W.; Haas, J.; Wagner, M.; Krohne, G.; Koszinowski, U.H. Cytomegalovirus recruitment of cellular kinases to dissolve the nuclear lamina. Science 2002, 297, 854–857. [Google Scholar]
- Lake, C.M.; Hutt-Fletcher, L.M. The epstein-barr virus bfrf1 and bflf2 proteins interact and coexpression alters their cellular localization. Virology 2004, 320, 99–106. [Google Scholar]
- Farina, A.; Feederle, R.; Raffa, S.; Gonnella, R.; Santarelli, R.; Frati, L.; Angeloni, A.; Torrisi, M.R.; Faggioni, A.; Delecluse, H.J. Bfrf1 of epstein-barr virus is essential for efficient primary viral envelopment and egress. J. Virol. 2005, 79, 3703–3712. [Google Scholar]
- Gonnella, R.; Farina, A.; Santarelli, R.; Raffa, S.; Feederle, R.; Bei, R.; Granato, M.; Modesti, A.; Frati, L.; Delecluse, H.J.; et al. Characterization and intracellular localization of the epstein-barr virus protein bflf2: Interactions with bfrf1 and with the nuclear lamina. J. Virol. 2005, 79, 3713–3727. [Google Scholar]
- Milbradt, J.; Auerochs, S.; Marschall, M. Cytomegaloviral proteins pul50 and pul53 are associated with the nuclear lamina and interact with cellular protein kinase c. J. Gen. Virol. 2007, 88, 2642–2650. [Google Scholar]
- Milbradt, J.; Auerochs, S.; Sticht, H.; Marschall, M. Cytomegaloviral proteins that associate with the nuclear lamina: Components of a postulated nuclear egress complex. J. Gen. Virol. 2009, 90, 579–590. [Google Scholar]
- Marschall, M.; Marzi, A.; aus dem Siepen, P.; Jochmann, R.; Kalmer, M.; Auerochs, S.; Lischka, P.; Leis, M.; Stamminger, T. Cellular p32 recruits cytomegalovirus kinase pul97 to redistribute the nuclear lamina. J. Biol. Chem. 2005, 280, 33357–33367. [Google Scholar]
- Hamirally, S.; Kamil, J.P.; Ndassa-Colday, Y.M.; Lin, A.J.; Jahng, W.J.; Baek, M.C.; Noton, S.; Silva, L.A.; Simpson-Holley, M.; Knipe, D.M.; et al. Viral mimicry of cdc2/cyclin-dependent kinase 1 mediates disruption of nuclear lamina during human cytomegalovirus nuclear egress. PLoS Pathog. 2009, 5, e1000275. [Google Scholar]
- Prichard, M.N. Function of human cytomegalovirus ul97 kinase in viral infection and its inhibition by maribavir. Rev. Med. Virol. 2009, 19, 215–229. [Google Scholar]
- Milbradt, J.; Webel, R.; Auerochs, S.; Sticht, H.; Marschall, M. Novel mode of phosphorylation-triggered reorganization of the nuclear lamina during nuclear egress of human cytomegalovirus. J. Biol. Chem. 2010, 285, 13979–13989. [Google Scholar]
- Lee, C.P.; Huang, Y.H.; Lin, S.F.; Chang, Y.; Chang, Y.H.; Takada, K.; Chen, M.R. Epstein-barr virus bglf4 kinase induces disassembly of the nuclear lamina to facilitate virion production. J. Virol. 2008, 82, 11913–11926. [Google Scholar]
- Miller, M.S.; Furlong, W.E.; Pennell, L.; Geadah, M.; Hertel, L. Rascal is a new human cytomegalovirus-encoded protein that localizes to the nuclear lamina and in cytoplasmic vesicles at late times postinfection. J. Virol. 2010, 84, 6483–6496. [Google Scholar]
- Klupp, B.G.; Granzow, H.; Mettenleiter, T.C. Effect of the pseudorabies virus us3 protein on nuclear membrane localization of the ul34 protein and virus egress from the nucleus. J. Gen. Virol. 2001, 82, 2363–2371. [Google Scholar]
- Morimoto, T.; Arii, J.; Tanaka, M.; Sata, T.; Akashi, H.; Yamada, M.; Nishiyama, Y.; Uema, M.; Kawaguchi, Y. Differences in the regulatory and functional effects of the us3 protein kinase activities of herpes simplex virus 1 and 2. J. Virol. 2009, 83, 11624–11634. [Google Scholar]
- de Noronha, C.M.; Sherman, M.P.; Lin, H.W.; Cavrois, M.V.; Moir, R.D.; Goldman, R.D.; Greene, W.C. Dynamic disruptions in nuclear envelope architecture and integrity induced by hiv-1 vpr. Science 2001, 294, 1105–1108. [Google Scholar]
- Segura-Totten, M.; Wilson, K.L. Hiv--breaking the rules for nuclear entry. Science 2001, 294, 1016–1017. [Google Scholar]
- Pelkmans, L.; Kartenbeck, J.; Helenius, A. Caveolar endocytosis of simian virus 40 reveals a new two-step vesicular-transport pathway to the er. Nat. Cell Biol. 2001, 3, 473–483. [Google Scholar]
- Gilbert, J.M.; Benjamin, T.L. Early steps of polyomavirus entry into cells. J. Virol. 2000, 74, 8582–8588. [Google Scholar]
- Gilbert, J.M.; Goldberg, I.G.; Benjamin, T.L. Cell penetration and trafficking of polyomavirus. J. Virol. 2003, 77, 2615–2622. [Google Scholar]
- Richterova, Z.; Liebl, D.; Horak, M.; Palkova, Z.; Stokrova, J.; Hozak, P.; Korb, J.; Forstova, J. Caveolae are involved in the trafficking of mouse polyomavirus virions and artificial vp1 pseudocapsids toward cell nuclei. J. Virol. 2001, 75, 10880–10891. [Google Scholar]
- Mannova, P.; Forstova, J. Mouse polyomavirus utilizes recycling endosomes for a traffic pathway independent of copi vesicle transport. J. Virol. 2003, 77, 1672–1681. [Google Scholar]
- Liebl, D.; Difato, F.; Hornikova, L.; Mannova, P.; Stokrova, J.; Forstova, J. Mouse polyomavirus enters early endosomes, requires their acidic ph for productive infection, and meets transferrin cargo in rab11-positive endosomes. J. Virol. 2006, 80, 4610–4622. [Google Scholar]
- Qian, M.; Cai, D.; Verhey, K.J.; Tsai, B. A lipid receptor sorts polyomavirus from the endolysosome to the endoplasmic reticulum to cause infection. PLoS Pathog. 2009, 5, e1000465. [Google Scholar]
- Maul, G.G.; Rovera, G.; Vorbrodt, A.; Abramczuk, J. Membrane fusion as a mechanism of simian virus 40 entry into different cellular compartments. J. Virol. 1978, 28, 936–944. [Google Scholar]
- Mackay, R.L.; Consigli, R.A. Early events in polyoma virus infection: Attachment, penetration, and nuclear entry. J. Virol. 1976, 19, 620–636. [Google Scholar]
- Nakanishi, A.; Shum, D.; Morioka, H.; Otsuka, E.; Kasamatsu, H. Interaction of the vp3 nuclear localization signal with the importin alpha 2/beta heterodimer directs nuclear entry of infecting simian virus 40. J. Virol. 2002, 76, 9368–9377. [Google Scholar]
- Gilbert, J.; Ou, W.; Silver, J.; Benjamin, T. Downregulation of protein disulfide isomerase inhibits infection by the mouse polyomavirus. J. Virol. 2006, 80, 10868–10870. [Google Scholar]
- Lilley, B.N.; Gilbert, J.M.; Ploegh, H.L.; Benjamin, T.L. Murine polyomavirus requires the endoplasmic reticulum protein derlin-2 to initiate infection. J. Virol. 2006, 80, 8739–8744. [Google Scholar]
- Schelhaas, M.; Malmstrom, J.; Pelkmans, L.; Haugstetter, J.; Ellgaard, L.; Grunewald, K.; Helenius, A. Simian virus 40 depends on er protein folding and quality control factors for entry into host cells. Cell 2007, 131, 516–529. [Google Scholar]
- Kuksin, D.; Norkin, L.C. Disassembly of simian virus 40 during passage through the endoplasmic reticulum and in the cytoplasm. J. Virol. 2012, 86, 1555–1562. [Google Scholar]
- Butin-Israeli, V.; Ben-Nun-Shaul, O.; Kopatz, I.; Adam, S.A.; Shimi, T.; Goldman, R.D.; Oppenheim, A. Simian virus 40 induces lamin a/c fluctuations and nuclear envelope deformation during cell entry. Nucleus 2011, 2. [Epub ahead of print]. [Google Scholar]
- Daniels, R.; Rusan, N.M.; Wadsworth, P.; Hebert, D.N. Sv40 vp2 and vp3 insertion into er membranes is controlled by the capsid protein vp1: Implications for DNA translocation out of the er. Mol. Cell 2006, 24, 955–966. [Google Scholar]
- Rainey-Barger, E.K.; Magnuson, B.; Tsai, B. A chaperone-activated nonenveloped virus perforates the physiologically relevant endoplasmic reticulum membrane. J. Virol. 2007, 81, 12996–13004. [Google Scholar]
- Huerfano, S.; Zila, V.; Boura, E.; Spanielova, H.; Stokrova, J.; Forstova, J. Minor capsid proteins of mouse polyomavirus are inducers of apoptosis when produced individually but are only moderate contributors to cell death during the late phase of viral infection. FEBS J. 2010, 277, 1270–1283. [Google Scholar]
- Cohen, S.; Behzad, A.R.; Carroll, J.B.; Pante, N. Parvoviral nuclear import: Bypassing the host nuclear-transport machinery. J. Gen. Virol. 2006, 87, 3209–3213. [Google Scholar]
- Cohen, S.; Marr, A.K.; Garcin, P.; Pante, N. Nuclear envelope disruption involving host caspases plays a role in the parvovirus replication cycle. J. Virol. 2011, 85, 4863–4874. [Google Scholar]
- Hansen, J.; Qing, K.; Srivastava, A. Infection of purified nuclei by adeno-associated virus 2. Mol. Ther. 2001, 4, 289–296. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cibulka, J.; Fraiberk, M.; Forstova, J. Nuclear Actin and Lamins in Viral Infections. Viruses 2012, 4, 325-347. https://doi.org/10.3390/v4030325
Cibulka J, Fraiberk M, Forstova J. Nuclear Actin and Lamins in Viral Infections. Viruses. 2012; 4(3):325-347. https://doi.org/10.3390/v4030325
Chicago/Turabian StyleCibulka, Jakub, Martin Fraiberk, and Jitka Forstova. 2012. "Nuclear Actin and Lamins in Viral Infections" Viruses 4, no. 3: 325-347. https://doi.org/10.3390/v4030325
APA StyleCibulka, J., Fraiberk, M., & Forstova, J. (2012). Nuclear Actin and Lamins in Viral Infections. Viruses, 4(3), 325-347. https://doi.org/10.3390/v4030325