Analysis of the Genome of the Sexually Transmitted Insect Virus Helicoverpa zea Nudivirus 2

Abstract
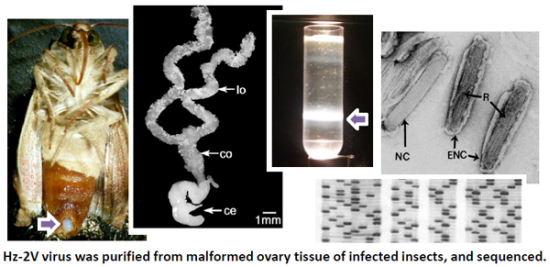
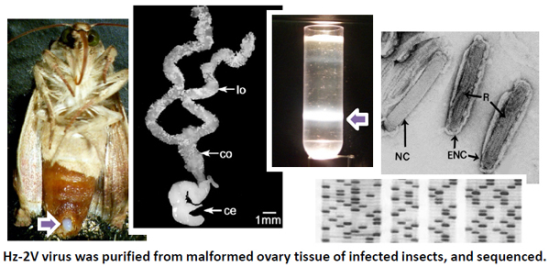
:
1. Introduction
2. Results and Discussion
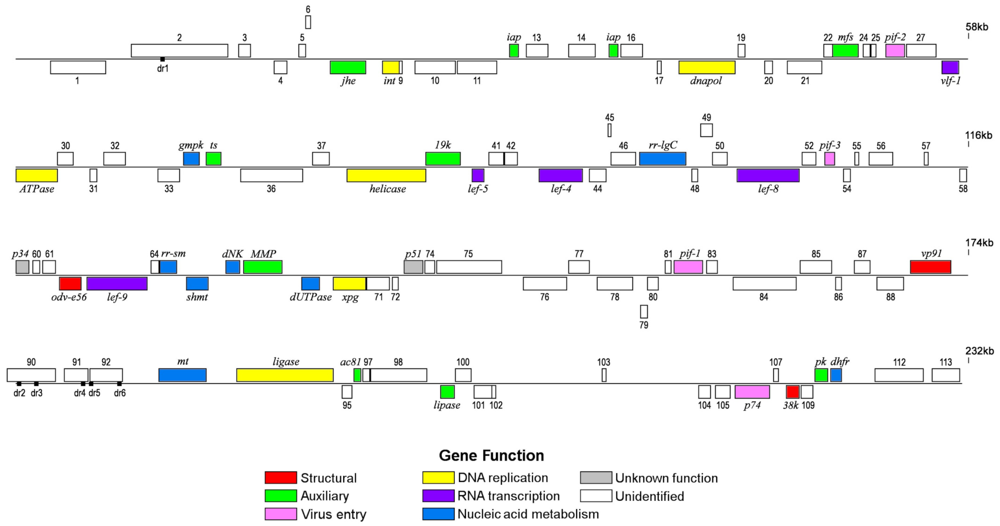
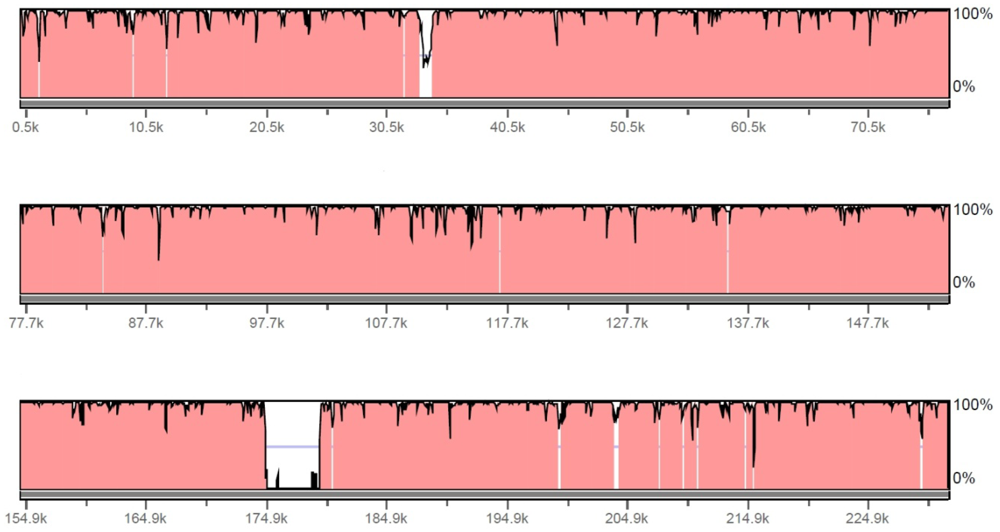
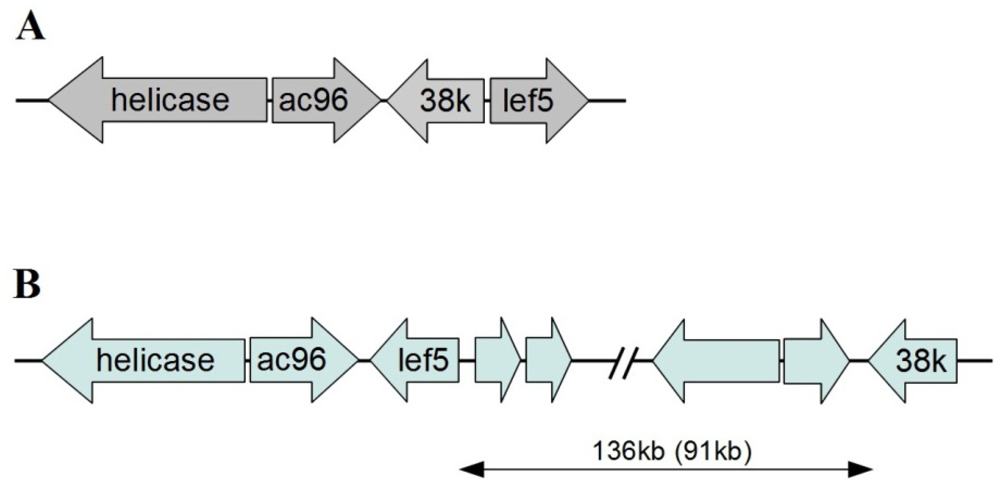
2.1. Genome Features
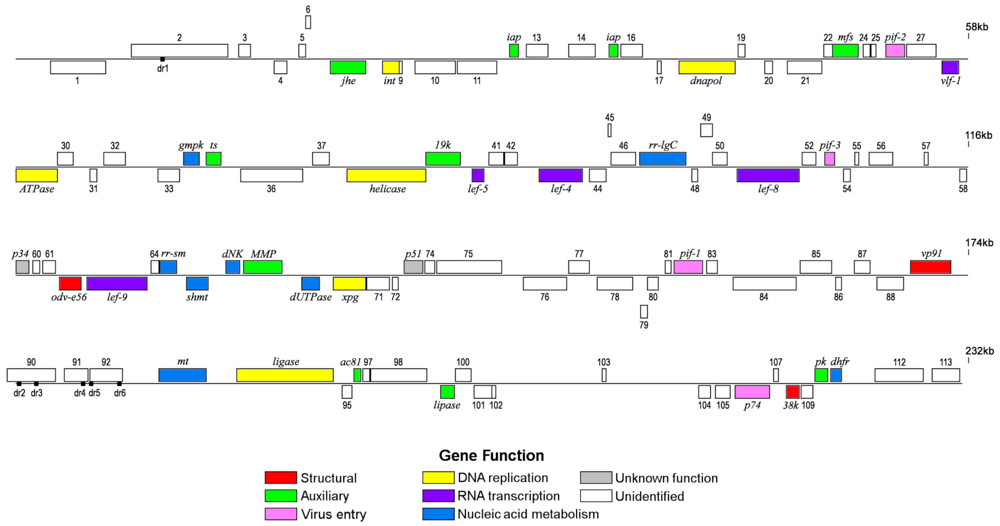

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF | Strand a | Position | Length | Best Blast match | Signature b | HzNV-1 c | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| start | end | (nt) | (aa) | Predicted function | Species/Viruses | E-value | ||||
| Hz2V001 | R | 5630 | 2298 | 3333 | 1111 | TAF, H3 | 1 | |||
| Hz2V002 | F | 7183 | 13047 | 5865 | 1955 | 154 | ||||
| Hz2V003 | F | 13677 | 14378 | 702 | 234 | 151 | ||||
| Hz2V004 | R | 16594 | 15830 | 765 | 255 | 150 | ||||
| Hz2V005 | F | 17366 | 17803 | 438 | 146 | 148 | ||||
| Hz2V006 | F | 17779 | 18063 | 285 | 95 | |||||
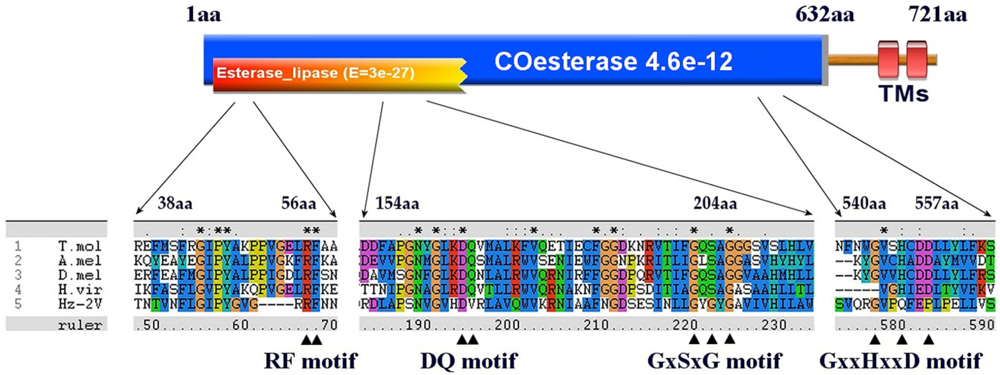
| Hz2V007 | R | 21403 | 19241 | 2163 | 721 | Carboxylesterase | Drosophila pseudoobscura | 3 × 10−21 | Coesterase | 145 |
| Hz2V008 | R | 23425 | 22421 | 1005 | 335 | Integrase | Monodon baculovirus | 1 × 10−29 | INT_REC_C | 144 |
| Hz2V009 | R | 23621 | 23430 | 192 | 64 | 143 | ||||
| Hz2V010 | R | 26839 | 24425 | 2415 | 805 | 141 | ||||
| Hz2V011 | R | 29383 | 26969 | 2415 | 805 | 140 | ||||
| Hz2V012 | F | 30140 | 30700 | 561 | 187 | Inhibitor of apoptosis protein | Trichoplusia ni SNPV | 7 × 10−25 | BIR | 138 |
| Hz2V013 | F | 31151 | 32494 | 1344 | 448 | 137 | ||||
| Hz2V014 | F | 33735 | 35312 | 1578 | 526 | |||||
| Hz2V015 | F | 36168 | 36713 | 546 | 182 | Inhibitor of apoptosis protein | Adoxophyes orana GV | 120 | BIR | 135 |
| Hz2V016 | F | 36879 | 38201 | 1323 | 441 | 134 | ||||
| Hz2V017 | R | 39326 | 39138 | 189 | 63 | 133 | ||||
| Hz2V018 | R | 43848 | 40441 | 3408 | 1136 | DNA polymerase | Oryctes rhinoceros NV | 1 × 10−25 | POLBc | 131 |
| Hz2V019 | F | 44032 | 44442 | 411 | 137 | 130 | ||||
| Hz2V020 | R | 46083 | 45655 | 429 | 143 | 129 | ||||
| Hz2V021 | R | 49108 | 47033 | 2076 | 692 | 128 | ||||
| Hz2V022 | F | 49210 | 49731 | 522 | 174 | |||||
| Hz2V023 | F | 49772 | 51307 | 1536 | 512 | Membrane transporter | Ades aegyptii | 1 × 10−135 | MFS_1 | 126 |
| Hz2V024 | F | 51575 | 52000 | 426 | 142 | 125 | ||||
| Hz2V025 | F | 52066 | 52392 | 327 | 109 | 124 | ||||
| Hz2V026 | F | 52998 | 54137 | 1140 | 380 | Per-os infectivity factor 2 | Gryllus bimaculatus NV | 5 × 10−37 | Baculo_44 | 123 |
| Hz2V027 | F | 54224 | 56023 | 1800 | 600 | 122 | ||||
| Hz2V028 | R | 57374 | 56376 | 999 | 333 | Very late expression factor 1 | Monodon baculovirus | 1 × 10−8 | 121 | |
| Hz2V029 | R | 60667 | 58199 | 2469 | 823 | DNA repair related ATPase | Gryllus bimaculatus NV | 3 × 10−4 | SbcC | 119 |
| Hz2V030 | F | 60709 | 61674 | 966 | 322 | 118 | ||||
| Hz2V031 | R | 63063 | 62671 | 393 | 131 | 117 | ||||
| Hz2V032 | F | 63532 | 64803 | 1272 | 424 | 115 | ||||
| Hz2V033 | R | 68086 | 66803 | 1284 | 428 | 112 | ||||
| Hz2V034 | F | 68319 | 69323 | 1005 | 335 | Guanosine monophosphate kinase | Monodon baculovirus | 4 × 10−5 | GMPK | 111 |
| Hz2V035 | F | 69749 | 70621 | 873 | 291 | Thymidylate synthase | Bombyx mori | 4 × 10−131 | Thymidylat_synt | 109 |
| Hz2V036 | R | 75567 | 71800 | 3768 | 1256 | 107 | ||||
| Hz2V037 | F | 76171 | 77190 | 1020 | 340 | 106 | ||||
| Hz2V038 | R | 83046 | 78304 | 4743 | 1581 | Helicase | Culex nigripalpus NPV | 2.8 | Pox_D5 | 104 |
| Hz2V039 | F | 83048 | 85156 | 2109 | 703 | 19kDa protein | Gryllus bimaculatus NV | 9 × 10−9 | Baculo_19 | 103 |
| Hz2V040 | R | 86584 | 85862 | 723 | 241 | Late expression factor 5 | Spodoptera exigua NPV | 6 × 10−3 | Baculo_LEF5 | 101 |
| Hz2V041 | F | 86863 | 87756 | 894 | 298 | 100 | ||||
| Hz2V042 | F | 87845 | 88594 | 750 | 250 | 99 | ||||
| Hz2V043 | R | 92546 | 89928 | 2619 | 873 | Late expression factor 4 | Gryllus bimaculatus NV | 3 × 10−18 | 98 | |
| Hz2V044 | R | 94003 | 92963 | 1041 | 347 | 97 | ||||
| Hz2V045 | F | 94110 | 94313 | 204 | 68 | |||||
| Hz2V046 | F | 94306 | 95775 | 1470 | 490 | 96 | ||||
| Hz2V047 | F | 96009 | 98822 | 2814 | 938 | Ribonuclease reductase | Spodoptera litura NPV | 0 | Ribonuc_red_lgC | 95 |
| Hz2V048 | R | 99573 | 99214 | 360 | 120 | 94 | ||||
| Hz2V049 | F | 99730 | 100473 | 744 | 248 | 93 | ||||
| Hz2V050 | F | 100458 | 101336 | 879 | 293 | |||||
| Hz2V051 | R | 105733 | 101939 | 3795 | 1265 | Late expression factor 8 | Gryllus bimaculatus NV | 1 × 10−12 | RNA_pol_Rpb2_6 | 90 |
| Hz2V052 | F | 105870 | 106712 | 843 | 281 | 89 | ||||
| Hz2V053 | F | 107253 | 107894 | 642 | 214 | Per-os infectivity factor 3 | Gryllus bimaculatus NV | 2 × 10−10 | 88 | |
| Hz2V054 | R | 108853 | 108413 | 441 | 147 | 87 | ||||
| Hz2V055 | F | 109110 | 109391 | 282 | 94 | 85 | ||||
| Hz2V056 | F | 109953 | 111365 | 1413 | 471 | 83 | ||||
| Hz2V057 | F | 113318 | 113530 | 213 | 71 | |||||
| Hz2V058 | R | 115909 | 115496 | 414 | 138 | 81 | ||||
| Hz2V059 | F | 116202 | 116963 | 762 | 254 | p34 late protein | Helicoverpa zea NV-1 d | 8 × 10−147 | 79 | |
| Hz2V060 | F | 117224 | 117595 | 372 | 124 | 78 | ||||
| Hz2V061 | F | 117803 | 118570 | 768 | 256 | 77 | ||||
| Hz2V062 | R | 120105 | 118786 | 1320 | 440 | Odv-e56 structural protein | Gryllus bimaculatus NV | 2 × 10−9 | 76 | |
| Hz2V063 | R | 124121 | 120513 | 3609 | 1203 | Late expression factor 9 | Monodon baculovirus | 1 × 10−60 | 75 | |
| Hz2V064 | F | 124357 | 124818 | 462 | 154 | 74 | ||||
| Hz2V065 | F | 124932 | 125930 | 999 | 333 | Ribonuclease reductase | Xenopus tropicalis | 5 × 10−128 | Ribonuc_red_sm | 73 |
| Hz2V066 | R | 127832 | 126510 | 1323 | 441 | Serine hydroxymethyltransferase | Bombyx mori | 0 | SHMT | 72 |
| Hz2V067 | F | 128939 | 129760 | 822 | 274 | Deoxynucleotide kinase | Drosophila melanogaster | 1 × 10−38 | dNK | 71 |
| Hz2V068 | F | 129972 | 132338 | 2367 | 789 | Matrix metalloprotease | Acyrthosiphon pisum | 3 × 10−69 | ZnMc_MMP, HX | 70 |
| Hz2V069 | R | 134585 | 133536 | 1050 | 350 | dUTPase | Culex quinquefasciatus | 2 × 10−38 | dUTPase | 69 |
| Hz2V070 | R | 137400 | 135430 | 1971 | 657 | DNA excision repair enzyme | Musca domestica SGHV | 2 × 10−7 | XPG | 68 |
| Hz2V071 | R | 138863 | 137478 | 1386 | 462 | 67 | ||||
| Hz2V072 | R | 139354 | 139019 | 336 | 112 | 66 | ||||
| Hz2V073 | F | 139721 | 140869 | 1149 | 383 | p51 late protein | Helicoverpa zea NV-1 d | 0 | 64 | |
| Hz2V074 | F | 141013 | 141564 | 552 | 184 | 63 | ||||
| Hz2V075 | F | 141693 | 145637 | 3945 | 1315 | 62 | ||||
| Hz2V076 | R | 149608 | 146981 | 2628 | 876 | 60 | ||||
| Hz2V077 | F | 149749 | 150978 | 1230 | 410 | 59 | ||||
| Hz2V078 | R | 153629 | 151443 | 2187 | 729 | 58 | ||||
| Hz2V079 | R | 154524 | 154090 | 435 | 145 | |||||
| Hz2V080 | R | 155191 | 154481 | 711 | 237 | |||||
| Hz2V081 | F | 155556 | 155936 | 381 | 127 | 56 | ||||
| Hz2V082 | F | 156150 | 157853 | 1704 | 568 | Per-os infectivity factor 1 | Gryllus bimaculatus NV | 6 × 10−23 | DUF686 | 55 |
| Hz2V083 | F | 158080 | 158742 | 663 | 221 | 54 | ||||
| Hz2V084 | R | 163557 | 159694 | 3864 | 1288 | 52 | ||||
| Hz2V085 | F | 163804 | 165702 | 1899 | 633 | 51 | ||||
| Hz2V086 | R | 166298 | 165954 | 345 | 115 | |||||
| Hz2V087 | F | 167038 | 168003 | 966 | 322 | 49 | ||||
| Hz2V088 | R | 170084 | 168429 | 1656 | 552 | 47 | ||||
| Hz2V089 | F | 170456 | 172948 | 2493 | 831 | vp91 capsid protein | Gryllus bimaculatus NV | 4 × 10−14 | 46 | |
| Hz2V090 | F | 173649 | 176570 | 2922 | 974 | |||||
| Hz2V091 | F | 177136 | 178524 | 1389 | 463 | |||||
| Hz2V092 | F | 178677 | 180614 | 1938 | 646 | |||||
| Hz2V093 | F | 182846 | 185689 | 2844 | 948 | Methyltransferase | Helicoverpa zea NPV | 8 × 10−28 | FtsJ | 37 |
| Hz2V094 | F | 187572 | 193430 | 5859 | 1953 | DNA ligase | Apis mellifera | 4 × 10−86 | DNA_ligase_A_M | 36 |
| Hz2V095 | R | 194548 | 193964 | 585 | 195 | 34 | ||||
| Hz2V096 | F | 194663 | 195130 | 468 | 156 | Ac81 | Gryllus bimaculatus NV | 6 × 10−12 | DUF845 | 33 |
| Hz2V097 | F | 195232 | 195630 | 399 | 133 | 32 | ||||
| Hz2V098 | F | 195699 | 199133 | 3435 | 1145 | 31 | ||||
| Hz2V099 | R | 200794 | 199931 | 864 | 288 | Esterase/lipase | Psychromonas ingrahamii | 0.16 | Aes | 30 |
| Hz2V100 | F | 200858 | 201796 | 939 | 313 | 29 | ||||
| Hz2V101 | R | 203059 | 201992 | 1068 | 356 | 28 | ||||
| Hz2V102 | R | 203298 | 203092 | 207 | 69 | |||||
| Hz2V103 | F | 209783 | 209980 | 198 | 66 | |||||
| Hz2V104 | R | 216330 | 215599 | 732 | 244 | 13 | ||||
| Hz2V105 | R | 217543 | 216668 | 876 | 292 | 12 | ||||
| Hz2V106 | R | 219936 | 217846 | 2091 | 697 | p74 envelope protein | Gryllius bimaculus NV | 5 × 10−57 | Baculo_p74 | 11 |
| Hz2V107 | F | 220177 | 220464 | 288 | 96 | |||||
| Hz2V108 | R | 221727 | 220951 | 777 | 259 | 38kDa protein | Monodon baculovirus | 3 × 10−15 | DUF705 | 10 |
| Hz2V109 | R | 222547 | 221819 | 729 | 243 | 9 | ||||
| Hz2V110 | F | 222677 | 223438 | 762 | 254 | Protein kinase | Trichomonas vaginalis | 5 × 10−18 | S_TKc | 8 |
| Hz2V111 | F | 223656 | 224297 | 642 | 214 | Dihydrofolate reductase | Heliothis virescens | 1 × 10−45 | DHFR | 7 |
| Hz2V112 | F | 226341 | 229232 | 2892 | 964 | 4 | ||||
| Hz2V113 | F | 229783 | 231468 | 1686 | 562 | 3 | ||||
| start | end | score | size | Count | %id | consensus | |
|---|---|---|---|---|---|---|---|
| dr1 | 11711 | 12310 | 134 | 24 | 25 | 63.2 | atgaagctgaggatgaatctgaac |
| dr2 | 174285 | 174572 | 132 | 36 | 8 | 79.2 | gaaactcctaaatcaaaggatgaa cctaaagcaaag |
| dr3 | 175417 | 175656 | 124 | 60 | 4 | 88.3 | atgaaaaagcaaaggctgaggcga aggctaaagccgatgctgctgcaa aagccaaagctg |
| dr4 | 178103 | 178390 | 182 | 24 | 12 | 85.8 | ttataccagagagcaagccagaaa |
| dr5 | 178517 | 178921 | 104 | 81 | 5 | 72.8 | acctaaagttgaatctaaagtagt ggaaccacctaaagcggaatctaa aacagtggaagctcctactaaaac agttgaagt |
| dr6 | 180236 | 180445 | 156 | 30 | 7 | 94.3 | agctgccgctaaacgcaaagccg aggctga |
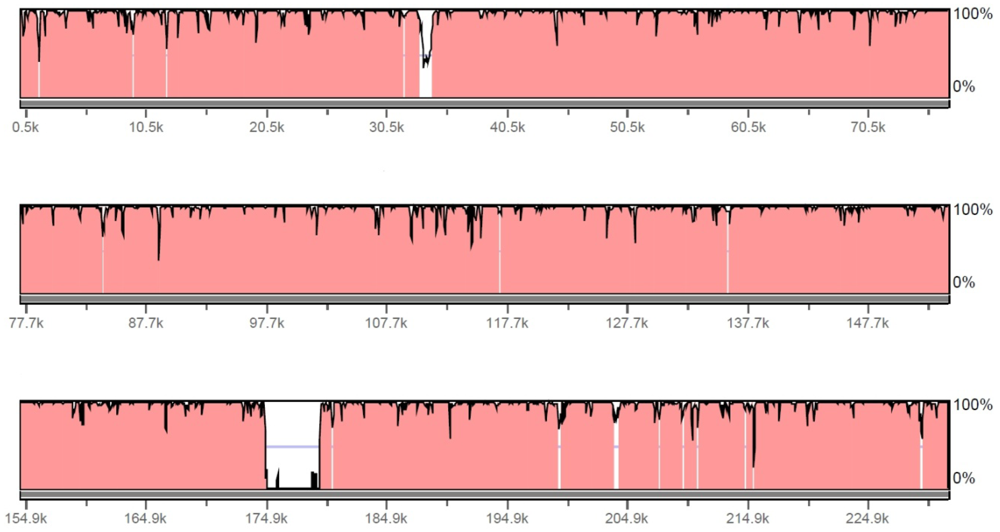
2.2. Evaluation of ORFs
2.3. Enzymes Related to DNA Replication and Repair
2.4. Proteins Involved in RNA Transcription
2.5. Genes Involved in Virus Entry
2.6. Enzymes Involved in Nucleic Acid Metabolism
2.7. Structural Proteins
2.8. Auxiliary and Undefined Genes
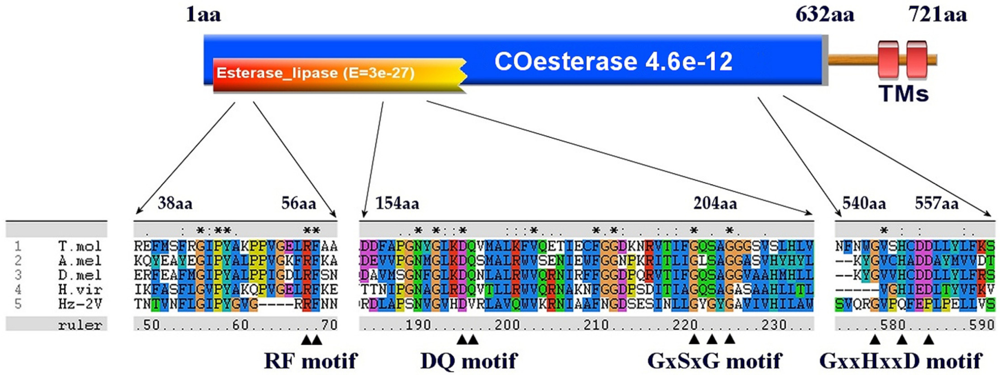
2.9. Comparison of HzNV-2 and Baculovirus Genome Contents
| Gene Function | Genes present in HzNV-2 | Genes absent in HzNV-2 |
|---|---|---|
| Replication | dnapol (ac65), helicase (ac95) | lef-1 (ac14), lef-2 (ac6) |
| Transcription | lef-4 (ac90), lef-5 (ac99), lef-8 (ac50), lef-9 (ac62), vlf-1 (ac77) | p47 (ac40) |
| Virus entry | p74 (ac138), pif-1 (ac119), pif-2 (ac22), pif-3 (ac115) | ld130 (ac23) |
| Structural | p91 (ac83), 38K (ac98), odv-e56 (ac148) | odv-e27 (ac144), gp41 (ac80), p6.9 (ac100), vp39 (ac89), vp1054 (ac54) |
| Auxiliary | alk-exo (ac133) | |
| Unknown | ac81, 19K (ac96) | ac68, ac92, ac109, ac142 |
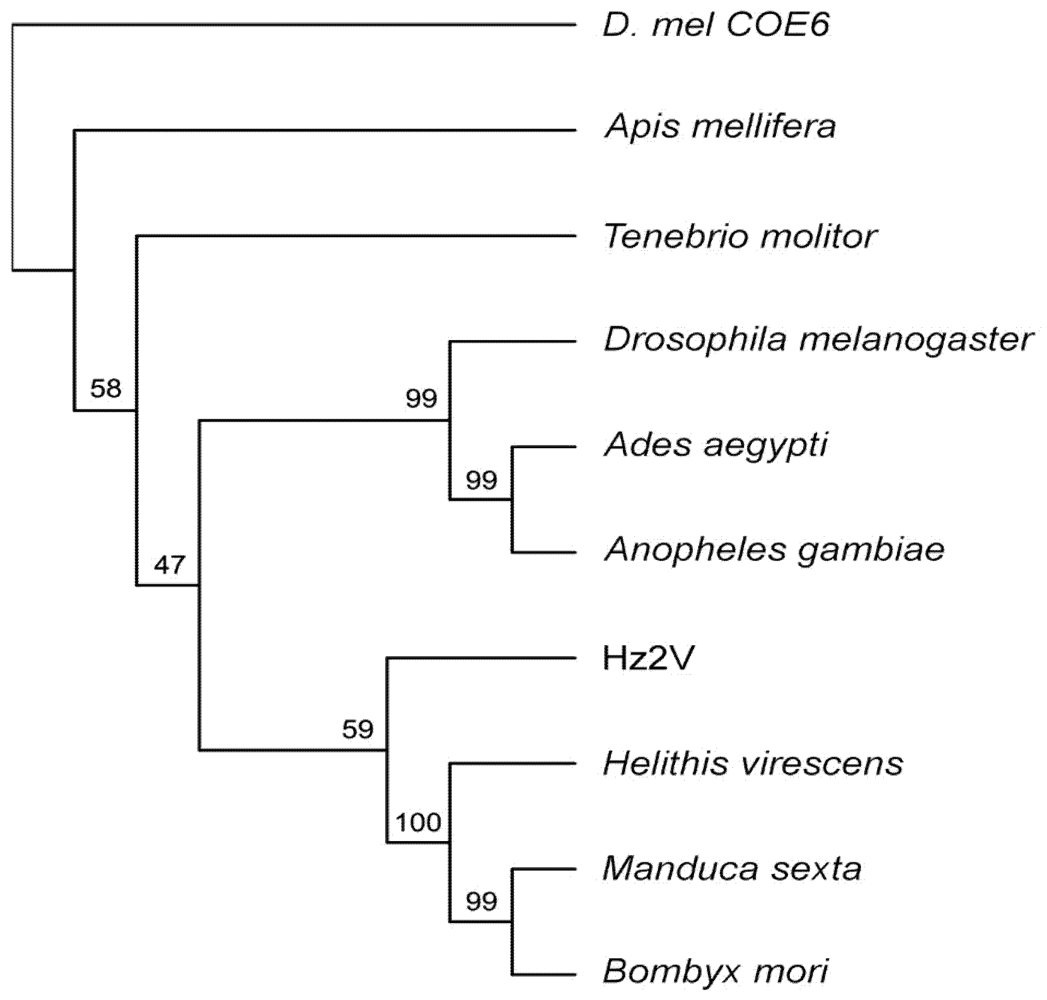
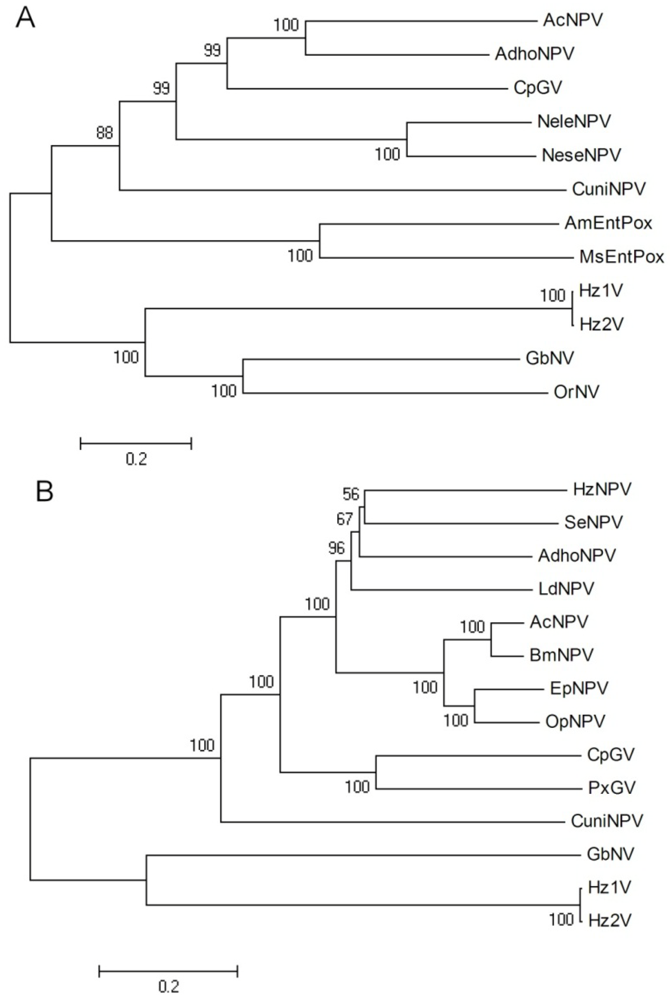
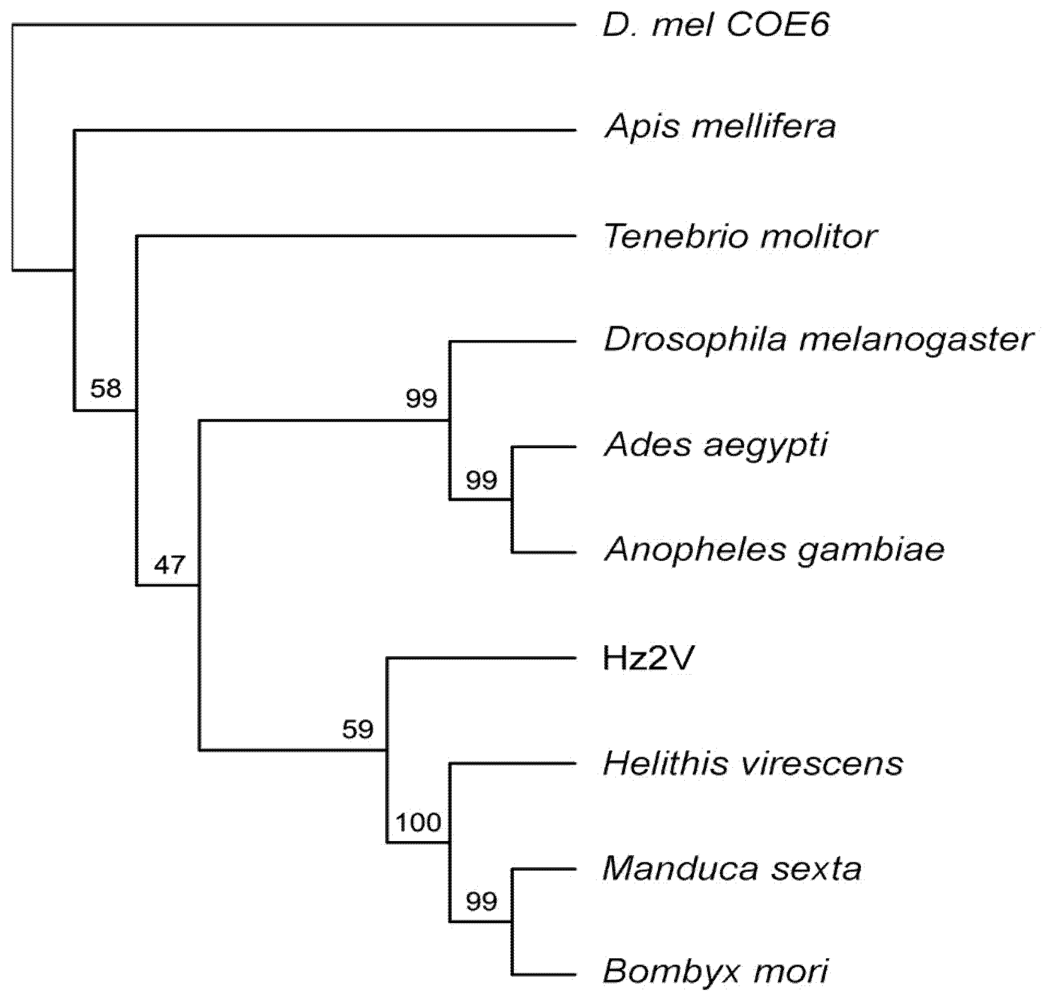
2.11. Phylogenetic Analysis
3. Experimental Section
3.1. Source of Viruses
3.2. Virus Purification and DNA Extraction
3.3. DNA Sequencing
3.4. Sequence Analysis
4. Summary and Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Wang, Y.-J.; Burand, J.P.; Jehle, J.A. Nudivirus genomics: Diversity and classification. Virol. Sin. 2007, 22, 128–136. [Google Scholar]
- Wang, Y.; Bininda-Emonds, O.R.P.; van Oers, M.M.; Vlak, J.M.; Jehle, J.A. The genome of oryctes rhinoceros nudivirus provides novel insight into the evolution of nuclear arthropod-specific large circular double-stranded DNA viruses. Virus Genes 2011, 42, 444–456. [Google Scholar]
- Wang, Y.; Jehle, J.A. Nudiviruses and other large, double-stranded circular DNA viruses of invertebrates: New insights on an old topic RID C-2243-2009. J. Invertebr. Pathol. 2009, 101, 187–193. [Google Scholar]
- Bezier, A.; Annaheim, M.; Herbiniere, J.; Wetterwald, C.; Gyapay, G.; Bernard-Samain, S.; Wincker, P.; Roditi, I.; Heller, M.; Belghazi, M.; et al. Polydnaviruses of braconid wasps derive from an ancestral nudivirus. Science 2009, 323, 926–930. [Google Scholar] [PubMed]
- Hamm, J.J.; Carpenter, J.E.; Styer, E.L. Oviposition day effect on incidence of agonadal progeny of Helicoverpa zea (Lepidoptera: Noctuidae) infected with a virus. Ann. Entomol. Soc. Am. 1996, 89, 266–275. [Google Scholar]
- Rallis, C.P.; Burand, J.P. Pathology and ultrastructure of Hz-2V infection in the agonadal female corn earworm, Helicoverpa zea. J. Invertebr. Pathol. 2002, 81, 33–44. [Google Scholar]
- Burand, J.P.; Rallis, C.P. In vivo dose-response of insects to Hz-2V infection. Virol. J. 2004, 1, 15. [Google Scholar] [CrossRef] [PubMed]
- Burand, J.P.; Tan, W.; Kim, W.; Nojima, S.; Roelofs, W. Infection with the insect virus Hz-2v alters mating behavior and pheromone production in female Helicoverpa zea moths. J. Insect Sci. 2005, 5, 6. [Google Scholar]
- Lu, H. Characterization of a novel baculovirus, gonad-specific virus, GSV. Ph.D. dissertation, University of Massachusetts, Amherst, MA, USA, 1997. [Google Scholar]
- Cochran, M.A.; Faulkner, P. Location of homologous DNA sequences interspersed at five regions in the baculovirus AcMNPV genome. J. Virol. 1983, 45, 961–970. [Google Scholar]
- Cheng, C.H.; Liu, S.M.; Chow, T.Y.; Hsiao, Y.Y.; Wang, D.P.; Huang, J.J.; Chen, H.H. Analysis of the complete genome sequence of the Hz-1 virus suggests that it is related to members of the Baculoviridae. J. Virol. 2002, 76, 9024–9034. [Google Scholar]
- Wang, Y.; Kleespies, R.G.; Huger, A.M.; Jehle, J.A. The genome of Gryllus bimaculatus nudivirus indicates an ancient diversification of baculovirus-related nonoccluded nudiviruses of insects. J. Virol. 2007, 81, 5395–5406. [Google Scholar]
- Pearson, M.N.; Rohrmann, G.F. Lymantria dispar nuclear polyhedrosis virus homologous regions: characterization of their ability to function as replication origins. J. Virol. 1995, 69, 213–221. [Google Scholar]
- Shamoo, Y.; Steitz, T.A. Building a replisome from interacting pieces: Sliding clamp complexed to a peptide from DNA polymerase and a polymerase editing complex. Cell 1999, 99, 155–166. [Google Scholar]
- Bressan, D.A.; Baxter, B.K.; Petrini, J.H. The Mre11-Rad50-Xrs2 protein complex facilitates homologous recombination-based double-strand break repair in Saccharomyces cerevisiae. Mol. Cell Biol. 1999, 19, 7681–7687. [Google Scholar]
- Hopfner, K.P.; Karcher, A.; Shin, D.S.; Craig, L.; Arthur, L.M.; Carney, J.P.; Tainer, J.A. Structural biology of Rad50 ATPase: ATP-driven conformational control in DNA double-strand break repair and the ABC-ATPase superfamily. Cell 2000, 101, 789–800. [Google Scholar]
- Mascarenhas, J.; Sanchez, H.; Tadesse, S.; Kidane, D.; Krisnamurthy, M.; Alonso, J.C.; Graumann, P.L. Bacillus subtilis SbcC protein plays an important role in DNA inter-strand cross-link repair. BMC Mol. Biol. 2006, 7, 20. [Google Scholar]
- Evans, E.; Klemperer, N.; Ghosh, R.; Traktman, P. The vaccinia virus D5 protein, which is required for DNA replication, is a nucleic acid-independent nucleoside triphosphatase. J. Virol. 1995, 69, 5353–5361. [Google Scholar] [PubMed]
- O'Donovan, A.; Davies, A.A.; Moggs, J.G.; West, S.C.; Wood, R.D. XPG endonuclease makes the 3' incision in human DNA nucleotide excision repair. Nature 1994, 371, 432–435. [Google Scholar]
- Cooper, P.K.; Nouspikel, T.; Clarkson, S.G.; Leadon, S.A. Defective transcription-coupled repair of oxidative base damage in Cockayne syndrome patients from XP group G. Science 1997, 275, 990–993. [Google Scholar]
- Hosfield, D.J.; Mol, C.D.; Shen, B.; Tainer, J.A. Structure of the DNA repair and replication endonuclease and exonuclease FEN-1: Coupling DNA and PCNA binding to FEN-1 activity. Cell 1998, 95, 135–146. [Google Scholar]
- Kim, K.; Biade, S.; Matsumoto, Y. Involvement of flap endonuclease 1 in base excision DNA repair. J. Biol. Chem. 1998, 273, 8842–8848. [Google Scholar]
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 151, 1257–1266. [Google Scholar]
- Guarino, L.A.; Xu, B.; Jin, J.; Dong, W. A virus-encoded RNA polymerase purified from baculovirus-infected cells. J. Virol. 1998, 72, 7985–7991. [Google Scholar]
- Mclachlin, J.R.; Miller, L.K. Identification and characterization of Vlf-1, a baculovirus gene involved in very late gene-expression. J. Virol. 1994, 68, 7746–7756. [Google Scholar]
- Yang, S.; Miller, L.K. Activation of baculovirus very late promoters by interaction with very late factor 1. J. Virol. 1999, 73, 3404–3409. [Google Scholar]
- Vanarsdall, A.L.; Okano, K.; Rohrmann, G.F. Characterization of the role of very late expression factor 1 in baculovirus capsid structure and DNA processing. J. Virol. 2006, 80, 1724–1733. [Google Scholar]
- Guttieri, M.C.; Burand, J.P. Nucleotide sequence, temporal expression, and transcriptional mapping of the p34 late gene of the Hz-1 insect virus. Virology 1996, 223, 370–375. [Google Scholar] [PubMed]
- Guarino, L.A.; Dong, W.; Jin, J. In vitro activity of the baculovirus late expression factor LEF-5. J. Virol. 2002, 76, 12663–12675. [Google Scholar] [PubMed]
- Jin, J.; Dong, W.; Guarino, L.A. The LEF-4 subunit of baculovirus RNA polymerase has RNA 5'-triphosphatase and ATPase activities. J. Virol. 1998, 72, 10011–10019. [Google Scholar]
- Knebel-Morsdorf, D.; Quadt, I.; Li, Y.; Montier, L.; Guarino, L.A. Expression of baculovirus late and very late genes depends on LEF-4, a component of the viral RNA polymerase whose guanyltransferase function is essential. J. Virol. 2006, 80, 4168–4173. [Google Scholar]
- Shuman, S.; Liu, Y.; Schwer, B. Covalent catalysis in nucleotidyl transfer reactions: essential motifs in Saccharomyces cerevisiae RNA capping enzyme are conserved in Schizosaccharomyces pombe and viral capping enzymes and among polynucleotide ligases. Proc. Natl. Acad. Sci. U. S. A. 1994, 91, 12046–12050. [Google Scholar]
- Passarelli, A.L.; Todd, J.W.; Miller, L.K. A baculovirus gene involved in late gene expression predicts a large polypeptide with a conserved motif of RNA polymerases. J. Virol. 1994, 68, 4673–4678. [Google Scholar]
- Lu, A.; Miller, L.K. Identification of three late expression factor genes within the 33.8- to 43.4-map-unit region of Autographa californica nuclear polyhedrosis virus. J. Virol. 1994, 68, 6710–6718. [Google Scholar] [PubMed]
- Broyles, S.S.; Moss, B. Homology between RNA polymerases of poxviruses, prokaryotes, and eukaryotes: Nucleotide sequence and transcriptional analysis of vaccinia virus genes encoding 147-kDa and 22-kDa subunits. Proc. Natl. Acad. Sci. U. S. A. 1986, 83, 3141–3145. [Google Scholar] [CrossRef] [PubMed]
- Acharya, A.; Gopinathan, K.P. Characterization of late gene expression factors lef-9 and lef-8 from Bombyx mori nucleopolyhedrovirus. J. Gen. Virol. 2002, 83, 2015–2023. [Google Scholar]
- Pijlman, G.P.; Pruijssers, A.J.; Vlak, J.M. Identification of pif-2, a third conserved baculovirus gene required for per os infection of insects. J. Gen. Virol. 2003, 84, 2041–2049. [Google Scholar]
- Li, X.; Song, J.; Jiang, T.; Liang, C.; Chen, X. The N-terminal hydrophobic sequence of Autographa californica nucleopolyhedrovirus PIF-3 is essential for oral infection. Arch. Virol. 2007, 152, 1851–1858. [Google Scholar]
- Ohkawa, T.; Washburn, J.O.; Sitapara, R.; Sid, E.; Volkman, L.E. Specific binding of Autographa californica M nucleopolyhedrovirus occlusion-derived virus to midgut cells of Heliothis virescens larvae is mediated by products of pif genes Ac119 and Ac022 but not by Ac115. J. Virol. 2005, 79, 15258–15264. [Google Scholar]
- Kikhno, I.; Gutierrez, S.; Croizier, L.; Croizier, G.; Ferber, M.L. Characterization of pif, a gene required for the per os infectivity of Spodoptera littoralis nucleopolyhedrovirus. J. Gen. Virol. 2002, 83, 3013–3022. [Google Scholar]
- Haas-Stapleton, E.J.; Washburn, J.O.; Volkman, L.E. P74 mediates specific binding of Autographa californica M nucleopolyhedrovirus occlusion-derived virus to primary cellular targets in the midgut epithelia of Heliothis virescens larvae. J. Virol. 2004, 78, 6786–6791. [Google Scholar]
- Rashidan, K.K.; Nassoury, N.; Tazi, S.; Giannopoulos, P.N.; Guertin, C. Choristoneura fumiferana Granulovirus p74 protein, a highly conserved baculoviral envelope protein. J. Biochem. Mol. Biol. 2003, 36, 475–487. [Google Scholar]
- Carreras, C.W.; Santi, D.V. The catalytic mechanism and structure of thymidylate synthase. Annu. Rev. Biochem. 1995, 64, 721–762. [Google Scholar]
- Perryman, S.M.; Rossana, C.; Deng, T.L.; Vanin, E.F.; Johnson, L.F. Sequence of a cDNA for mouse thymidylate synthase reveals striking similarity with the prokaryotic enzyme. Mol. Biol. Evol. 1986, 3, 313–321. [Google Scholar]
- Chen, H.H.; Tso, D.J.; Yeh, W.B.; Cheng, H.J.; Wu, T.F. The thymidylate synthase gene of Hz-1 virus: A gene captured from its lepidopteran host. Insect Mol. Biol. 2001, 10, 495–503. [Google Scholar]
- Garcia-Maruniak, A.; Maruniak, J.E.; Farmerie, W.; Boucias, D.G. Sequence analysis of a non-classified, non-occluded DNA virus that causes salivary gland hypertrophy of Musca domestica, MdSGHV. Virology 2008, 377, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Jordan, A.; Reichard, P. Ribonucleotide reductases. Annu. Rev. Biochem. 1998, 67, 71–98. [Google Scholar]
- Blakley, R.L. The interconversion of serine and glycine: Participation of pyridoxal phosphate. Biochem. J. 1955, 61, 315–323. [Google Scholar]
- Blakley, R.L. The Biochemistry of Folic Acid and Related Pteridines; North-Holland Pub. Co.: Amsterdam, The Netherlands, 1969; p. xxi, 570. [Google Scholar]
- Ives, D.H.; Ikeda, S. Life on the salvage path: The deoxynucleoside kinase of Lactobacillus acidophilus R-26. Progr. Nucleic Acid Res. Mol. Biol. 1998, 59, 205–255. [Google Scholar]
- Elder, J.H.; Lerner, D.L.; Hasselkus-Light, C.S.; Fontenot, D.J.; Hunter, E.; Luciw, P.A.; Montelaro, R.C.; Phillips, T.R. Distinct subsets of retroviruses encode dUTPase. J. Virol. 1992, 66, 1791–1794. [Google Scholar]
- Chen, R.; Wang, H.; Mansky, L.M. Roles of uracil-DNA glycosylase and dUTPase in virus replication. J. Gen. Virol. 2002, 83, 2339–2345. [Google Scholar]
- Miller, R.J.; Cairns, J.S.; Bridges, S.; Sarver, N. Human immunodeficiency virus and AIDS: Insights from animal lentiviruses. J. Virol. 2000, 74, 7187–7195. [Google Scholar]
- Wu, X.; Guarino, L.A. Autographa californica nucleopolyhedrovirus orf69 encodes an RNA cap (nucleoside-2'-O)-methyltransferase. J. Virol. 2003, 77, 3430–3440. [Google Scholar]
- Schnell, J.R.; Dyson, H.J.; Wright, P.E. Structure, dynamics, and catalytic function of dihydrofolate reductas. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 119–140. [Google Scholar]
- Braunagel, S.C.; Elton, D.M.; Ma, H.; Summers, M.D. Identification and analysis of an Autographa californica nuclear polyhedrosis virus structural protein of the occlusion-derived virus envelope: ODV-E56. Virology 1996, 217, 97–110. [Google Scholar]
- Russell, R.L.; Rohrmann, G.F. Characterization of P91, a protein associated with virions of an Orgyia pseudotsugata baculovirus. Virology 1997, 233, 210–223. [Google Scholar]
- Wu, W.; Liang, H.; Kan, J.; Liu, C.; Yuan, M.; Liang, C.; Yang, K.; Pang, Y. Autographa californica multiple nucleopolyhedrovirus 38K is a novel nucleocapsid protein that interacts with VP1054, VP39, VP80, and itself. J. Virol. 2008, 82, 12356–12364. [Google Scholar] [PubMed]
- Gilbert, L.I.; Granger, N.A.; Roe, R.M. The juvenile hormones: Historical facts and speculations on future research directions. Insect Biochem. Mol. Biol. 2000, 30, 617–644. [Google Scholar]
- Ward, V.K.; Bonning, B.C.; Huang, T.; Shiotsuki, T.; Griffeth, V.N.; Hammock, B.D. Analysis of the catalytic mechanism of juvenile-hormone esterase by site-directed mutagenesis. Int. J. Biochem. 1992, 24, 1933–1941. [Google Scholar]
- Crook, N.E.; Clem, R.J.; Miller, L.K. An apoptosis-inhibiting baculovirus gene with a zinc finger-like motif. J. Virol. 1993, 67, 2168–2174. [Google Scholar]
- Uren, A.G.; Coulson, E.J.; Vaux, D.L. Conservation of baculovirus inhibitor of apoptosis repeat proteins (BIRPs) in viruses, nematodes, vertebrates and yeasts. Trends Biochem. Sci. 1998, 23, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.K. An exegesis of IAPs: Salvation and surprises from BIR motifs. Trends Cell Biol. 1999, 9, 323–328. [Google Scholar]
- Joazeiro, C.A.; Weissman, A.M. RING finger proteins: Mediators of ubiquitin ligase activity. Cell 2000, 102, 549–552. [Google Scholar]
- Wilson, R.; Goyal, L.; Ditzel, M.; Zachariou, A.; Baker, D.A.; Agapite, J.; Steller, H.; Meier, P. The DIAP1 RING finger mediates ubiquitination of Dronc and is indispensable for regulating apoptosis. Nat. Cell Biol. 2002, 4, 445–450. [Google Scholar]
- Pao, S.S.; Paulsen, I.T.; Saier, M.H., Jr. Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 1998, 62, 1–34. [Google Scholar]
- Konrad, M. Cloning and expression of the essential gene for guanylate kinase from yeast. J. Biol. Chem. 1992, 267, 25652–25655. [Google Scholar]
- Hall, S.W.; Kuhn, H. Purification and properties of guanylate kinase from bovine retinas and rod outer segments. Eur. J. Biochem. 1986, 161, 551–556. [Google Scholar]
- Guttieri, M.C.; Burand, J.P. Location, nucleotide sequence, and regulation of the p51 late gene of the Hz-1 insect virus: Identification of a putative late regulatory element. Virus Genes 2001, 23, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.J.; Murphy, G.; Reynolds, J.J. The origin of matrix metalloproteinases and their familial relationships. FEBS Lett. 1991, 289, 4–7. [Google Scholar]
- Nagase, H.; Woessner, J.F., Jr. Matrix metalloproteinases. J. Biol. Chem. 1999, 274, 21491–21494. [Google Scholar]
- Wang, P.; Granados, R.R. An intestinal mucin is the target substrate for a baculovirus enhancin. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 6977–6982. [Google Scholar]
- Herniou, E.A.; Olszewski, J.A.; Cory, J.S.; O'Reilly, D.R. The genome sequence and evolution of baculoviruses. Annu. Rev. Entomol. 2003, 48, 211–234. [Google Scholar]
- Chen, H.Q.; Chen, K.P.; Yao, Q.; Guo, Z.J.; Wang, L.L. Characterization of a late gene, ORF67 from Bombyx mori nucleopolyhedrovirus. FEBS Lett. 2007, 581, 5836–5842. [Google Scholar]
- Peist, R.; Koch, A.; Bolek, P.; Sewitz, S.; Kolbus, T.; Boos, W. Characterization of the aes gene of Escherichia coli encoding an enzyme with esterase activity. J. Bacteriol. 1997, 179, 7679–7686. [Google Scholar]
- Joly, N.; Danot, O.; Schlegel, A.; Boos, W.; Richet, E. The Aes protein directly controls the activity of MalT, the central transcriptional activator of the Escherichia coli maltose regulon. J. Biol. Chem. 2002, 277, 16606–16613. [Google Scholar]
- Edelman, A.M.; Blumenthal, D.K.; Krebs, E.G. Protein serine/threonine kinases. Annu. Rev. Biochem. 1987, 56, 567–613. [Google Scholar]
- Mikhailov, V.S.; Rohrmann, G.F. Baculovirus replication factor LEF-1 is a DNA primase. J. Virol. 2002, 76, 2287–2297. [Google Scholar]
- Evans, J.T.; Leisy, D.J.; Rohrmann, G.F. Characterization of the interaction between the baculovirus replication factors LEF-1 and LEF-2. J. Virol. 1997, 71, 3114–3119. [Google Scholar]
- Guarino, L.A.; Summers, M.D. Interspersed homologous DNA of autographa californica nuclear polyhedrosis virus enhances delayed-early gene expression. J. Virol. 1986, 60, 215–223. [Google Scholar]
- Grula, M.A.; Buller, P.L.; Weaver, R.F. alpha-Amanitin-Resistant viral RNA synthesis in nuclei isolated from nuclear polyhedrosis virus-infected Heliothis zea Larvae and Spodoptera frugiperda cells. J. Virol. 1981, 38, 916–921. [Google Scholar]
- Pearson, M.N.; Groten, C.; Rohrmann, G.F. Identification of the lymantria dispar nucleopolyhedrovirus envelope fusion protein provides evidence for a phylogenetic division of the Baculoviridae. J. Virol. 2000, 74, 6126–6131. [Google Scholar]
- Garcia-Maruniak, A.; Maruniak, J.E.; Zanotto, P.M.; Doumbouya, A.E.; Liu, J.C.; Merritt, T.M.; Lanoie, J.S. Sequence analysis of the genome of the Neodiprion sertifer nucleopolyhedrovirus. J. Virol. 2004, 78, 7036–7051. [Google Scholar]
- Lauzon, H.A.; Lucarotti, C.J.; Krell, P.J.; Feng, Q.; Retnakaran, A.; Arif, B.M. Sequence and organization of the Neodiprion lecontei nucleopolyhedrovirus genome. J. Virol. 2004, 78, 7023–7035. [Google Scholar]
- Belyavskyi, M.; Braunagel, S.C.; Summers, M.D. The structural protein ODV-EC27 of Autographa californica nucleopolyhedrovirus is a multifunctional viral cyclin. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 11205–11210. [Google Scholar]
- Mikhailov, V.S.; Okano, K.; Rohrmann, G.F. Baculovirus alkaline nuclease possesses a 5'-->3' exonuclease activity and associates with the DNA-binding protein LEF-3. J. Virol. 2003, 77, 2436–2444. [Google Scholar]
- Li, L.; Rohrmann, G.F. Characterization of a baculovirus alkaline nuclease. J. Virol. 2000, 74, 6401–6407. [Google Scholar]
- Raina, A.K.; Adams, J.R. Gonad-specific virus of corn-earworm. Nature 1995, 374, 770–770. [Google Scholar]
- Sanger, F.; Air, G.M.; Barrell, B.G.; Brown, N.L.; Coulson, A.R.; Fiddes, J.C.; Hutchison, C.A.; Slocombe, P.M.; Smith, M. Nucleotide-sequence of bacteriophage Phichi174 DNA. Nature 1977, 265, 687–695. [Google Scholar]
- Ewing, B.; Hillier, L.; Wendl, M.C.; Green, P. Base-calling of automated sequencer traces using phred. I. Accuracy assessment. Genome Res. 1998, 8, 175–185. [Google Scholar] [PubMed]
- Ewing, W.R.; Choi, Y.M.; Becker, M.; Manetta, V.; Green, D.; Davis, R.; Mason, H.; Ly, C.; Cha, D.; McGarry, D.; et al. Sulfonamidopyrrolidinones: Design, SAR and parenteral activity of a novel class of factor Xa inhibitors. Abstr. Paper. Am. Chem. Soc. 1998, 215, U920–U920. [Google Scholar]
- Sutton, G.G.; White, O.; Adams, M.D.; Kerlavage, A.R. TIGR assembler: A new tool for assembling large shotgun sequencing projects. Genome Sci. Technol. 1995, 1, 9–19. [Google Scholar]
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res. 1999, 9, 868–877. [Google Scholar]
- Hexamer. Available online: ftp://ftp.sanger.ac.uk/pub/rd/ (accessed on 1 January 2012).
- Salzberg, S.L.; Delcher, A.L.; Kasif, S.; White, O. Microbial gene identification using interpolated Markov models. Nucleic Acids Res. 1998, 26, 544–548. [Google Scholar]
- Delcher, A.L.; Harmon, D.; Kasif, S.; White, O.; Salzberg, S.L. Improved microbial gene identification with GLIMMER. Nucleic Acids Res. 1999, 27, 4636–4641. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997, 25, 3389–3402. [Google Scholar]
- Pearson, W.R. Rapid and sensitive sequence comparison with FASTP and FASTA. Meth. Enzymol. 1990, 183, 63–98. [Google Scholar]
- Eddy, S.R.; Mitchison, G.; Durbin, R. Maximum discrimination hidden Markov models of sequence consensus. J. Comput. Biol. 1995, 2, 9–23. [Google Scholar]
- Burks, C. Molecular biology database list. Nucleic Acids Res. 1999, 27, 1–9. [Google Scholar]
- Loots, G.; Ovcharenko, I. VISTA 2.0: Evolutionary analysis of transcription factor binding sites. Nucleic Acids Res. 2004, 32, W217–W221. [Google Scholar] [PubMed]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: identification of signaling domains. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 5857–5864. [Google Scholar]
- Marchler-Bauer, A.; Bryant, S.H. CD-Search: Protein domain annotations on the fly. Nucleic Acids Res. 2004, 32, W327–331. [Google Scholar]
- Marchler-Bauer, A.; Anderson, J.B.; DeWeese-Scott, C.; Fedorova, N.D.; Geer, L.Y.; He, S.; Hurwitz, D.I.; Jackson, J.D.; Jacobs, A.R.; Lanczycki, C.J.; et al. CDD: A curated Entrez database of conserved domain alignments. Nucleic Acids Res. 2003, 31, 383–387. [Google Scholar] [PubMed]
- Devereux, J.; Haeberli, P.; Smithies, O. A comprehensive set of sequence analysis programs for the VAX. Nucleic Acids Res. 1984, 12, 387–395. [Google Scholar]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The european molecular biology open software suite. Trends Genet. 2000, 16, 276–277. [Google Scholar]
- Nakai, K.; Horton, P. PSORT: A program for detecting sorting signals in proteins and predicting their subcellular localization. Trends Biochem. Sci. 1999, 24, 34–36. [Google Scholar]
- Sonnhammer, E.L.; von Heijne, G.; Krogh, A. A hidden Markov model for predicting transmembrane helices in protein sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1998, 6, 175–182. [Google Scholar]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar]
- Bendtsen, J.D.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Deleage, G.; Combet, C.; Blanchet, C.; Geourjon, C. ANTHEPROT: An integrated protein sequence analysis software with client/server capabilities. Comput. Biol. Med. 2001, 31, 259–267. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Granados, R.R.; Nguyen, T.; Cato, B. An insect cell line persistently infected with a baculovirus-like particle. Intervirology 1978, 10, 309–317. [Google Scholar]
- Hink, W.F.; Ignoffo, C.M. Establishment of a new cell line (IMC-HZ-1) from ovaries of cotton bollworm moths, Heliothis zea (boddie). Exp. Cell Res. 1970, 60, 307–309. [Google Scholar]
- Dubrovsky, E.B. Hormonal cross talk in insect development. Trends Endocrinol. Metab. 2005, 16, 6–11. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Burand, J.P.; Kim, W.; Afonso, C.L.; Tulman, E.R.; Kutish, G.F.; Lu, Z.; Rock, D.L. Analysis of the Genome of the Sexually Transmitted Insect Virus Helicoverpa zea Nudivirus 2. Viruses 2012, 4, 28-61. https://doi.org/10.3390/v4010028
Burand JP, Kim W, Afonso CL, Tulman ER, Kutish GF, Lu Z, Rock DL. Analysis of the Genome of the Sexually Transmitted Insect Virus Helicoverpa zea Nudivirus 2. Viruses. 2012; 4(1):28-61. https://doi.org/10.3390/v4010028
Chicago/Turabian StyleBurand, John P., Woojin Kim, Claudio L. Afonso, Edan R. Tulman, Gerald F. Kutish, Zhiqiang Lu, and Daniel L. Rock. 2012. "Analysis of the Genome of the Sexually Transmitted Insect Virus Helicoverpa zea Nudivirus 2" Viruses 4, no. 1: 28-61. https://doi.org/10.3390/v4010028
APA StyleBurand, J. P., Kim, W., Afonso, C. L., Tulman, E. R., Kutish, G. F., Lu, Z., & Rock, D. L. (2012). Analysis of the Genome of the Sexually Transmitted Insect Virus Helicoverpa zea Nudivirus 2. Viruses, 4(1), 28-61. https://doi.org/10.3390/v4010028