Cell Surface Markers in HTLV-1 Pathogenesis


Abstract
:1. Pathogenetic Properties of Human T Cell Lymphotropic Virus Type 1 (HTLV-1)
2. Viral Effector Molecules
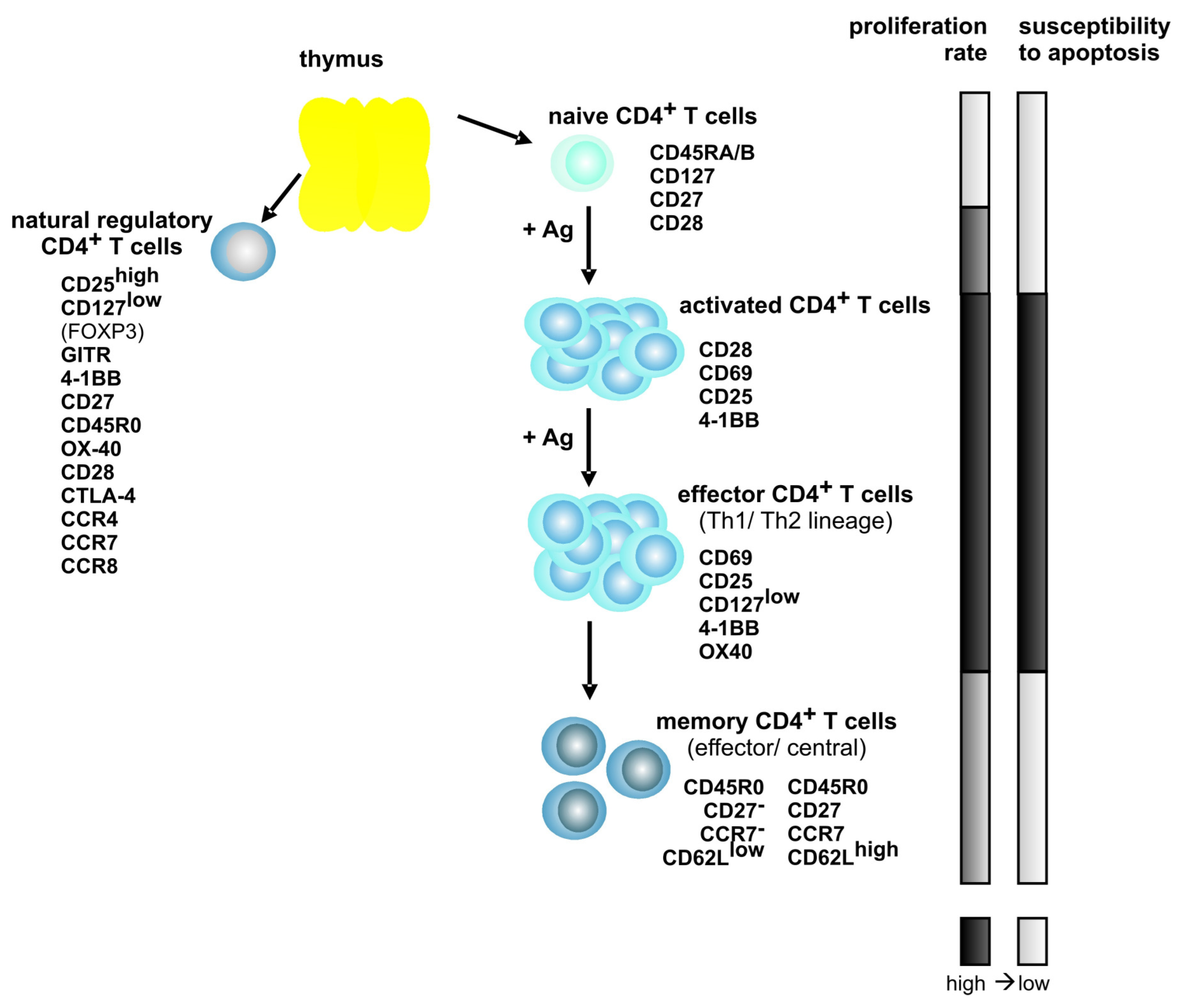
3. Differentiation of Human T Lymphocytes and HTLV-1-Persistence
4. Lineage and Activation Markers of HTLV-1-Infected Cells
5. Costimulatory Receptors of the Tumor Necrosis Factor Receptor (TNFR) Superfamily
6. Chemokine Receptors
7. Interleukin Receptors
8. Adhesion Molecules on HTLV-1-Infected Cells
9. Contribution of Cell Surface Markers to Longevity and Pathogenesis
Acknowledgments
Conflict of Interest
References and Notes
- Poiesz, B.J.; Ruscetti, F.W.; Gazdar, A.F.; Bunn, P.A.; Minna, J.D.; Gallo, R.C. Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc. Natl. Acad. Sci. U. S. A. 1980, 77, 7415–7419. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Miyoshi, I.; Hinuma, Y. Isolation and characterization of retrovirus from cell lines of human adult T-cell leukemia and its implication in the disease. Proc. Natl. Acad. Sci. U. S. A. 1982, 79, 2031–2035. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Seiki, M.; Yamaguchi, K.; Takatsuki, K. Monoclonal integration of human T-cell leukemia provirus in all primary tumors of adult T-cell leukemia suggests causative role of human T-cell leukemia virus in the disease. Proc. Natl. Acad. Sci. U. S. A. 1984, 81, 2534–2537. [Google Scholar] [CrossRef] [PubMed]
- Gessain, A.; Barin, F.; Vernant, J.C.; Gout, O.; Maurs, L.; Calender, A.; de The, G. Antibodies to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis. Lancet 1985, 2, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Osame, M.; Usuku, K.; Izumo, S.; Ijichi, N.; Amitani, H.; Igata, A.; Matsumoto, M.; Tara, M. HTLV-I associated myelopathy, a new clinical entity. Lancet 1986, 1, 1031–1032. [Google Scholar] [CrossRef]
- Matsuoka, M.; Jeang, K.T. Human T-cell leukaemia virus type 1 (HTLV-1) infectivity and cellular transformation. Nat. Rev. Cancer 2007, 7, 270–280. [Google Scholar] [CrossRef]
- Verdonck, K.; Gonzalez, E.; Van, D.S.; Vandamme, A.M.; Vanham, G.; Gotuzzo, E. Human T-lymphotropic virus 1: recent knowledge about an ancient infection. Lancet Infect. Dis. 2007, 7, 266–281. [Google Scholar] [CrossRef] [PubMed]
- Bangham, C.R.; Osame, M. Cellular immune response to HTLV-1. Oncogene 2005, 24, 6035–6046. [Google Scholar] [CrossRef]
- Richardson, J.H.; Edwards, A.J.; Cruickshank, J.K.; Rudge, P.; Dalgleish, A.G. In vivo cellular tropism of human T-cell leukemia virus type 1. J. Virol. 1990, 64, 5682–5687. [Google Scholar] [CrossRef]
- Asquith, B.; Zhang, Y.; Mosley, A.J.; de Lara, C.M.; Wallace, D.L.; Worth, A.; Kaftantzi, L.; Meekings, K.; Griffin, G.E.; Tanaka, Y.; et al. In vivo T lymphocyte dynamics in humans and the impact of human T-lymphotropic virus 1 infection. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 8035–8040. [Google Scholar] [CrossRef]
- Mesnard, J.M.; Barbeau, B.; Devaux, C. HBZ, a new important player in the mystery of adult T-cell leukemia. Blood 2006, 108, 3979–3982. [Google Scholar] [CrossRef] [PubMed]
- Nicot, C.; Harrod, R.L.; Ciminale, V.; Franchini, G. Human T-cell leukemia/lymphoma virus type 1 nonstructural genes and their functions. Oncogene 2005, 24, 6026–6034. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, M. HTLV-1 bZIP factor gene: Its roles in HTLV-1 pathogenesis. Mol. Aspects Med. 2010, 31, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Kashanchi, F.; Brady, J.N. Transcriptional and post-transcriptional gene regulation of HTLV-1. Oncogene 2005, 24, 5938–5951. [Google Scholar] [CrossRef]
- Bogerd, H.P.; Tiley, L.S.; Cullen, B.R. Specific binding of the human T-cell leukemia virus type I Rex protein to a short RNA sequence located within the Rex-response element. J. Virol. 1992, 66, 7572–7575. [Google Scholar] [CrossRef]
- Grone, M.; Koch, C.; Grassmann, R. The HTLV-1 Rex protein induces nuclear accumulation of unspliced viral RNA by avoiding intron excision and degradation. Virology 1996, 218, 316–325. [Google Scholar] [CrossRef]
- Grassmann, R.; Dengler, C.; Muller-Fleckenstein, I.; Fleckenstein, B.; McGuire, K.; Dokhelar, M.C.; Sodroski, J.G.; Haseltine, W.A. Transformation to continuous growth of primary human T lymphocytes by human T-cell leukemia virus type I X-region genes transduced by a Herpesvirus saimiri vector. Proc. Natl. Acad. Sci. U. S. A. 1989, 86, 3351–3355. [Google Scholar] [CrossRef]
- Grassmann, R.; Berchtold, S.; Radant, I.; Alt, M.; Fleckenstein, B.; Sodroski, J.G.; Haseltine, W.A.; Ramstedt, U. Role of human T-cell leukemia virus type 1 X region proteins in immortalization of primary human lymphocytes in culture. J. Virol. 1992, 66, 4570–4575. [Google Scholar] [CrossRef]
- Akagi, T.; Shimotohno, K. Proliferative response of Tax1-transduced primary human T cells to anti-CD3 antibody stimulation by an interleukin-2-independent pathway. J. Virol. 1993, 67, 1211–1217. [Google Scholar] [CrossRef]
- Hasegawa, H.; Sawa, H.; Lewis, M.J.; Orba, Y.; Sheehy, N.; Yamamoto, Y.; Ichinohe, T.; Tsunetsugu-Yokota, Y.; Katano, H.; Takahashi, H.; et al. Thymus-derived leukemia-lymphoma in mice transgenic for the Tax gene of human T-lymphotropic virus type I. Nat. Med. 2006, 12, 466–472. [Google Scholar] [CrossRef]
- Boxus, M.; Twizere, J.C.; Legros, S.; Dewulf, J.F.; Kettmann, R.; Willems, L. The HTLV-1 Tax interactome. Retrovirology 2008, 5, 76. [Google Scholar] [CrossRef] [PubMed]
- Grassmann, R.; Aboud, M.; Jeang, K.T. Molecular mechanisms of cellular transformation by HTLV-1 Tax. Oncogene 2005, 24, 5976–5985. [Google Scholar] [CrossRef] [PubMed]
- Kannian, P.; Green, P.L. Human T Lymphotropic Virus Type 1 (HTLV-1): Molecular Biology and Oncogenesis. Viruses 2010, 2, 2037–2077. [Google Scholar] [CrossRef]
- Kress, A.K.; Kalmer, M.; Rowan, A.G.; Grassmann, R.; Fleckenstein, B. The tumor marker Fascin is strongly induced by the Tax oncoprotein of HTLV-1 through NF-kappaB signals. Blood 2011, 117, 3609–3612. [Google Scholar] [CrossRef] [PubMed]
- Peloponese, J.M.; Yeung, M.L.; Jeang, K.T. Modulation of nuclear factor-kappaB by human T cell leukemia virus type 1 Tax protein: Implications for oncogenesis and inflammation. Immunol. Res. 2006, 34, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Fujii, M. Distinct functions of HTLV-1 Tax1 from HTLV-2 Tax2 contribute key roles to viral pathogenesis. Retrovirology 2009, 6, 117. [Google Scholar] [CrossRef]
- Fujii, M.; Chuhjo, T.; Minamino, T.; Masaaki, N.; Miyamoto, K.; Seiki, M. Identification of the Tax interaction region of serum response factor that mediates the aberrant induction of immediate early genes through CArG boxes by HTLV-I Tax. Oncogene 1995, 11, 7–14. [Google Scholar]
- Jeang, K.T. HTLV-1 and adult T-cell leukemia: insights into viral transformation of cells 30 years after virus discovery. J. Formos. Med. Assoc. 2010, 109, 688–693. [Google Scholar] [CrossRef]
- Blom, B.; Spits, H. Development of human lymphoid cells. Annu.Rev.Immunol. 2006, 24, 287–320. [Google Scholar] [CrossRef]
- Lee, M.S.; Hanspers, K.; Barker, C.S.; Korn, A.P.; McCune, J.M. Gene expression profiles during human CD4+ T cell differentiation. Int. Immunol. 2004, 16, 1109–1124. [Google Scholar] [CrossRef]
- Lanzavecchia, A.; Sallusto, F. Understanding the generation and function of memory T cell subsets. Curr. Opin. Immunol. 2005, 17, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Pacholczyk, R.; Ignatowicz, H.; Kraj, P.; Ignatowicz, L. Origin and T cell receptor diversity of Foxp3+CD4+CD25+ T cells. Immunity 2006, 25, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Bacchetta, R.; Gambineri, E.; Roncarolo, M.G. Role of regulatory T cells and FOXP3 in human diseases. J. Allergy Clin. Immunol. 2007, 120, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Pfoertner, S.; Jeron, A.; Probst-Kepper, M.; Guzman, C.A.; Hansen, W.; Westendorf, A.M.; Toepfer, T.; Schrader, A.J.; Franzke, A.; Buer, J.; et al. Signatures of human regulatory T cells: an encounter with old friends and new players. Genome Biol. 2006, 7, R54.1–R54.18. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Miyara, M.; Costantino, C.M.; Hafler, D.A. FOXP3+ regulatory T cells in the human immune system. Nat. Rev. Immunol. 2010, 10, 490–500. [Google Scholar] [CrossRef]
- Yi, H.; Zhen, Y.; Jiang, L.; Zheng, J.; Zhao, Y. The phenotypic characterization of naturally occurring regulatory CD4+CD25+ T cells. Cell Mol. Immunol. 2006, 3, 189–195. [Google Scholar]
- Sallusto, F.; Geginat, J.; Lanzavecchia, A. Central memory and effector memory T cell subsets: Function, generation, and maintenance. Annu. Rev. Immunol. 2004, 22, 745–763. [Google Scholar] [CrossRef]
- Croft, M. Costimulation of T cells by OX40, 4–1BB, and CD27. Cytokine Growth Factor Rev. 2003, 14, 265–273. [Google Scholar] [CrossRef]
- Harhaj, E.W.; Good, L.; Xiao, G.; Sun, S.C. Gene expression profiles in HTLV-I-immortalized T cells: Deregulated expression of genes involved in apoptosis regulation. Oncogene 1999, 18, 1341–1349. [Google Scholar] [CrossRef]
- Pise-Masison, C.A.; Radonovich, M.; Mahieux, R.; Chatterjee, P.; Whiteford, C.; Duvall, J.; Guillerm, C.; Gessain, A.; Brady, J.N. Transcription profile of cells infected with human T-cell leukemia virus type I compared with activated lymphocytes. Cancer Res. 2002, 62, 3562–3571. [Google Scholar]
- Ruckes, T.; Saul, D.; Van Snick, J.; Hermine, O.; Grassmann, R. Autocrine antiapoptotic stimulation of cultured adult T-cell leukemia cells by overexpression of the chemokine I-309. Blood 2001, 98, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Ng, P.W.; Iha, H.; Iwanaga, Y.; Bittner, M.; Chen, Y.; Jiang, Y.; Gooden, G.; Trent, J.M.; Meltzer, P.; Jeang, K.T.; Zeichner, S.L. Genome-wide expression changes induced by HTLV-1 Tax: Evidence for MLK-3 mixed lineage kinase involvement in Tax-mediated NF-kappaB activation. Oncogene 2001, 20, 4484–4496. [Google Scholar] [CrossRef] [PubMed]
- Pichler, K.; Kattan, T.; Gentzsch, J.; Kress, A.K.; Taylor, G.P.; Bangham, C.R.; Grassmann, R. Strong induction of 4-1BB, a growth and survival promoting costimulatory receptor, in HTLV-1-infected cultured and patients’ T cells by the viral Tax oncoprotein. Blood 2008, 111, 4741–4751. [Google Scholar] [CrossRef]
- Baba, M.; Okamoto, M.; Hamasaki, T.; Horai, S.; Wang, X.; Ito, Y.; Suda, Y.; Arima, N. Highly enhanced expression of CD70 on human T-lymphotropic virus type 1-carrying T-cell lines and adult T-cell leukemia cells. J. Virol. 2008, 82, 3843–3852. [Google Scholar] [CrossRef]
- Tsukasaki, K.; Hermine, O.; Bazarbachi, A.; Ratner, L.; Ramos, J.C.; Harrington, W., Jr.; O’Mahony, D.; Janik, J.E.; Bittencourt, A.L.; Taylor, G.P.; Yamaguchi, K.; Utsunomiya, A.; Tobinai, K.; Watanabe, T. Definition, prognostic factors, treatment, and response criteria of adult T-cell leukemia-lymphoma: A proposal from an international consensus meeting. J. Clin. Oncol. 2009, 27, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Kobayashi, S.; Ohno, N.; Isobe, M.; Tsuda, M.; Zaike, Y.; Watanabe, N.; Tani, K.; Tojo, A.; Uchimaru, K. Leukemic T cells are specifically enriched in a unique CD3(dim) CD7(low) subpopulation of CD4(+) T cells in acute-type adult T-cell leukemia. Cancer Sci. 2011, 102, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Vidal, C.; Matsushita, S.; Colamonici, O.R.; Trepel, J.B.; Mitsuya, H.; Neckers, L.M. Human T lymphotropic virus I infection deregulates surface expression of the transferrin receptor. J. Immunol. 1988, 141, 984–988. [Google Scholar] [CrossRef]
- Yamano, Y.; Takenouchi, N.; Li, H.C.; Tomaru, U.; Yao, K.; Grant, C.W.; Maric, D.A.; Jacobson, S. Virus-induced dysfunction of CD4+CD25+ T cells in patients with HTLV-I-associated neuroimmunological disease. J. Clin. Invest. 2005, 115, 1361–1368. [Google Scholar] [CrossRef]
- Tonks, N.K.; Charbonneau, H.; Diltz, C.D.; Fischer, E.H.; Walsh, K.A. Demonstration that the leukocyte common antigen CD45 is a protein tyrosine phosphatase. Biochemistry 1988, 27, 8695–8701. [Google Scholar] [CrossRef]
- Hermiston, M.L.; Xu, Z.; Weiss, A. CD45: A critical regulator of signaling thresholds in immune cells. Annu. Rev. Immunol. 2003, 21, 107–137. [Google Scholar] [CrossRef]
- Holmes, N. CD45: All is not yet crystal clear. Immunology 2006, 117, 145–155. [Google Scholar] [CrossRef]
- Richardson, J.H.; Edwards, A.J.; Cruickshank, J.K.; Rudge, P.; Dalgleish, A.G. In vivo cellular tropism of human T-cell leukemia virus type 1. J. Virol. 1990, 64, 5682–5687. [Google Scholar] [CrossRef] [PubMed]
- Yasunaga, J.; Sakai, T.; Nosaka, K.; Etoh, K.; Tamiya, S.; Koga, S.; Mita, S.; Uchino, M.; Mitsuya, H.; Matsuoka, M. Impaired production of naive T lymphocytes in human T-cell leukemia virus type I-infected individuals: Its implications in the immunodeficient state. Blood 2001, 97, 3177–3183. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Matsuoka, H.; Yamashita, K.; Maeda, K.; Kawano, K.; Uno, H.; Tsubouchi, H. CD45RO expression on peripheral lymphocytes as a prognostic marker for adult T-cell leukemia. Leuk. Lymphoma. 1998, 28, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Moro, H.; Iwai, K.; Mori, N.; Watanabe, M.; Fukushi, M.; Oie, M.; Arai, M.; Tanaka, Y.; Miyawaki, T.; Gejyo, F.; et al. Interleukin-2-dependent but not independent T-cell lines infected with human T-cell leukemia virus type 1 selectively express CD45RO, a marker for persistent infection in vivo. Virus Genes 2001, 23, 263–271. [Google Scholar] [CrossRef]
- Schmitt, I.; Rosin, O.; Rohwer, P.; Gossen, M.; Grassmann, R. Stimulation of cyclin-dependent kinase activity and G1- to S-phase transition in human lymphocytes by the human T-cell leukemia/lymphotropic virus type 1 Tax protein. J. Virol. 1998, 72, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Waldele, K.; Silbermann, K.; Schneider, G.; Ruckes, T.; Cullen, B.; Grassmann, R. Requirement of the human T-cell leukemia virus (HTLV-1) tax- stimulated hiap-1 gene for the survival of transformed lymphocytes. Blood 2006, 107, 4491–4499. [Google Scholar] [CrossRef] [PubMed]
- Kress, A.K.; Schneider, G.; Pichler, K.; Kalmer, M.; Fleckenstein, B.; Grassmann, R. Elevated cyclic AMP levels in T lymphocytes transformed by human T-cell lymphotropic virus type 1. J. Virol. 2010, 84, 8732–8742. [Google Scholar] [CrossRef] [PubMed]
- Dezzutti, C.S.; Rudolph, D.L.; Lal, R.B. Infection with human T-lymphotropic virus types I and II results in alterations of cellular receptors, including the up-modulation of T-cell counterreceptors CD40, CD54, and CD80 (B7–1). Clin. Diagn. Lab. Immunol. 1995, 2, 349–355. [Google Scholar] [CrossRef]
- Lal, R.B.; Rudolph, D.L.; Dezzutti, C.S.; Linsley, P.S.; Prince, H.E. Costimulatory effects of T cell proliferation during infection with human T lymphotropic virus types I and II are mediated through CD80 and CD86 ligands. J. Immunol. 1996, 157, 1288–1296. [Google Scholar] [CrossRef]
- Nurieva, R.I.; Liu, X.; Dong, C. Yin-Yang of costimulation: Crucial controls of immune tolerance and function. Immunol. Rev. 2009, 229, 88–100. [Google Scholar] [CrossRef]
- Lal, R.B.; Rudolph, D.L.; Dezzutti, C.S.; Linsley, P.S.; Prince, H.E. Costimulatory effects of T cell proliferation during infection with human T lymphotropic virus types I and II are mediated through CD80 and CD86 ligands. J. Immunol. 1996, 157, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, K.; Harashima, N.; Hanabuchi, S.; Masuda, M.; Utsunomiya, A.; Tanosaki, R.; Tomonaga, M.; Ohashi, T.; Hasegawa, A.; Masuda, T.; et al. Potential immunogenicity of adult T cell leukemia cells in vivo. Int. J. Cancer 2005, 114, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Harhaj, N.S.; Janic, B.; Ramos, J.C.; Harrington, W.J., Jr.; Harhaj, E.W. Deregulated expression of CD40 ligand in HTLV-I infection: Distinct mechanisms of downregulation in HTLV-I-transformed cell lines and ATL patients. Virology 2007, 362, 99–108. [Google Scholar] [CrossRef]
- Watts, T.H. TNF/TNFR family members in costimulation of T cell responses. Annu. Rev. Immunol. 2005, 23, 23–68. [Google Scholar] [CrossRef] [PubMed]
- Rivera, I.; Harhaj, E.W.; Sun, S.C. Involvement of NF-AT in type I human T-cell leukemia virus Tax-mediated Fas ligand promoter transactivation. J. Biol. Chem. 1998, 273, 22382–22388. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zachar, V.; Zdravkovic, M.; Guo, M.; Ebbesen, P.; Liu, X. Role of the Fas/Fas ligand pathway in apoptotic cell death induced by the human T cell lymphotropic virus type I Tax transactivator. J. Gen. Virol. 1997, 78, 3277–3285. [Google Scholar] [CrossRef]
- Hanon, E.; Hall, S.; Taylor, G.P.; Saito, M.; Davis, R.; Tanaka, Y.; Usuku, K.; Osame, M.; Weber, J.N.; Bangham, C.R. Abundant tax protein expression in CD4+ T cells infected with human T-cell lymphotropic virus type I (HTLV-I) is prevented by cytotoxic T lymphocytes. Blood 2000, 95, 1386–1392. [Google Scholar] [CrossRef]
- Ohshima, K.; Haraoka, S.; Sugihara, M.; Suzumiya, J.; Kawasaki, C.; Kanda, M.; Kikuchi, M. Amplification and expression of a decoy receptor for fas ligand (DcR3) in virus (EBV or HTLV-I) associated lymphomas. Cancer Lett. 2000, 160, 89–97. [Google Scholar] [CrossRef]
- Croft, M. The role of TNF superfamily members in T-cell function and diseases. Nat. Rev. Immunol. 2009, 9, 271–285. [Google Scholar] [CrossRef]
- Salek-Ardakani, S.; Croft, M. Regulation of CD4 T cell memory by OX40 (CD134). Vaccine 2006, 24, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Miura, S.; Ohtani, K.; Numata, N.; Niki, M.; Ohbo, K.; Ina, Y.; Gojobori, T.; Tanaka, Y.; Tozawa, H.; Nakamura, M. Molecular cloning and characterization of a novel glycoprotein, gp34, that is specifically induced by the human T-cell leukemia virus type I transactivator p40tax. Mol. Cell Biol. 1991, 11, 1313–1325. [Google Scholar] [PubMed]
- Ohtani, K.; Tsujimoto, A.; Tsukahara, T.; Numata, N.; Miura, S.; Sugamura, K.; Nakamura, M. Molecular mechanisms of promoter regulation of the gp34 gene that is trans-activated by an oncoprotein Tax of human T cell leukemia virus type I. J. Biol. Chem. 1998, 273, 14119–14129. [Google Scholar] [CrossRef] [PubMed]
- Baum, P.R.; Gayle, R.B., III; Ramsdell, F.; Srinivasan, S.; Sorensen, R.A.; Watson, M.L.; Seldin, M.F.; Baker, E.; Sutherland, G.R.; Clifford, K.N. Molecular characterization of murine and human OX40/OX40 ligand systems: identification of a human OX40 ligand as the HTLV-1-regulated protein gp34. EMBO J. 1994, 13, 3992–4001. [Google Scholar] [CrossRef]
- Godfrey, W.R.; Fagnoni, F.F.; Harara, M.A.; Buck, D.; Engleman, E.G. Identification of a human OX-40 ligand, a costimulator of CD4+ T cells with homology to tumor necrosis factor. J. Exp. Med. 1994, 180, 757–762. [Google Scholar] [CrossRef]
- Pankow, R.; Durkop, H.; Latza, U.; Krause, H.; Kunzendorf, U.; Pohl, T.; Bulfone-Paus, S. The HTLV-I tax protein transcriptionally modulates OX40 antigen expression. J. Immunol. 2000, 165, 263–270. [Google Scholar] [CrossRef]
- Imura, A.; Hori, T.; Imada, K.; Kawamata, S.; Tanaka, Y.; Imamura, S.; Uchiyama, T. OX40 expressed on fresh leukemic cells from adult T-cell leukemia patients mediates cell adhesion to vascular endothelial cells: Implication for the possible involvement of OX40 in leukemic cell infiltration. Blood 1997, 89, 2951–2958. [Google Scholar] [CrossRef]
- Nocentini, G.; Riccardi, C. GITR: A multifaceted regulator of immunity belonging to the tumor necrosis factor receptor superfamily. Eur. J. Immunol. 2005, 35, 1016–1022. [Google Scholar] [CrossRef]
- Bal, H.P.; Cheng, J.; Murakami, A.; Tallarico, A.S.; Wang, W.; Zhou, D.; Vasicek, T.J.; Marasco, W.A. GITR overexpression on CD4+CD25+ HTLV-1 transformed cells: Detection by massively parallel signature sequencing. Biochem. Biophys. Res. Commun. 2005, 332, 569–584. [Google Scholar] [CrossRef]
- Kohno, T.; Yamada, Y.; Akamatsu, N.; Kamihira, S.; Imaizumi, Y.; Tomonaga, M.; Matsuyama, T. Possible origin of adult T-cell leukemia/lymphoma cells from human T lymphotropic virus type-1-infected regulatory T cells. Cancer Sci. 2005, 96, 527–533. [Google Scholar] [CrossRef]
- Harhaj, E.W.; Harhaj, N.S.; Grant, C.; Mostoller, K.; Alefantis, T.; Sun, S.C.; Wigdahl, B. Human T cell leukemia virus type I Tax activates CD40 gene expression via the NF-kappa B pathway. Virology 2005, 333, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Thelen, M.; Stein, J.V. How chemokines invite leukocytes to dance. Nat. Immunol. 2008, 9, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, H.; Nomura, T.; Kohno, M.; Tateishi, N.; Suzuki, Y.; Maeda, N.; Fujisawa, R.; Yoshie, O.; Fujita, S. Increased chemokine receptor CCR7/EBI1 expression enhances the infiltration of lymphoid organs by adult T-cell leukemia cells. Blood 2000, 95, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Nagakubo, D.; Jin, Z.; Hieshima, K.; Nakayama, T.; Shirakawa, A.K.; Tanaka, Y.; Hasegawa, H.; Hayashi, T.; Tsukasaki, K.; Yamada, Y.; Yoshie, O. Expression of CCR9 in HTLV-1+ T cells and ATL cells expressing Tax. Int. J. Cancer 2007, 120, 1591–1597. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.A.; Meingassner, J.G.; Lipp, M.; Moore, H.D.; Rot, A. CCR7 is required for the in vivo function of CD4+ CD25+ regulatory T cells. J. Exp. Med. 2007, 204, 735–745. [Google Scholar] [CrossRef]
- Burgstahler, R.; Kempkes, B.; Steube, K.; Lipp, M. Expression of the chemokine receptor BLR2/EBI1 is specifically transactivated by Epstein-Barr virus nuclear antigen 2. Biochem. Biophys. Res. Commun. 1995, 215, 737–743. [Google Scholar] [CrossRef]
- Harasawa, H.; Yamada, Y.; Hieshima, K.; Jin, Z.; Nakayama, T.; Yoshie, O.; Shimizu, K.; Hasegawa, H.; Hayashi, T.; Imaizumi, Y.; et al. Survey of chemokine receptor expression reveals frequent co-expression of skin-homing CCR4 and CCR10 in adult T-cell leukemia/lymphoma. Leuk. Lymphoma 2006, 47, 2163–2173. [Google Scholar] [CrossRef]
- Yoshie, O.; Fujisawa, R.; Nakayama, T.; Harasawa, H.; Tago, H.; Izawa, D.; Hieshima, K.; Tatsumi, Y.; Matsushima, K.; Hasegawa, H.; et al. Frequent expression of CCR4 in adult T-cell leukemia and human T-cell leukemia virus type 1-transformed T cells. Blood 2002, 99, 1505–1511. [Google Scholar] [CrossRef]
- Ishida, T.; Utsunomiya, A.; Iida, S.; Inagaki, H.; Takatsuka, Y.; Kusumoto, S.; Takeuchi, G.; Shimizu, S.; Ito, M.; Komatsu, H.; Wakita, A.; Eimoto, T.; Matsushima, K.; Ueda, R. Clinical significance of CCR4 expression in adult T-cell leukemia/lymphoma: Its close association with skin involvement and unfavorable outcome. Clin. Cancer Res. 2003, 9, 3625–3634. [Google Scholar]
- Ishida, T.; Iida, S.; Akatsuka, Y.; Ishii, T.; Miyazaki, M.; Komatsu, H.; Inagaki, H.; Okada, N.; Fujita, T.; Shitara, K.; et al. The CC chemokine receptor 4 as a novel specific molecular target for immunotherapy in adult T-Cell leukemia/lymphoma. Clin. Cancer Res. 2004, 10, 7529–7539. [Google Scholar] [CrossRef]
- Yamamoto, K.; Utsunomiya, A.; Tobinai, K.; Tsukasaki, K.; Uike, N.; Uozumi, K.; Yamaguchi, K.; Yamada, Y.; Hanada, S.; Tamura, K.; et al. Phase I study of KW-0761, a defucosylated humanized anti-CCR4 antibody, in relapsed patients with adult T-cell leukemia-lymphoma and peripheral T-cell lymphoma. J. Clin. Oncol. 2010, 28, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Hieshima, K.; Nagakubo, D.; Nakayama, T.; Shirakawa, A.K.; Jin, Z.; Yoshie, O. Tax-inducible production of CC chemokine ligand 22 by human T cell leukemia virus type 1 (HTLV-1)-infected T cells promotes preferential transmission of HTLV-1 to CCR4-expressing CD4+ T cells. J. Immunol. 2008, 180, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Twizere, J.C.; Springael, J.Y.; Boxus, M.; Burny, A.; Dequiedt, F.; Dewulf, J.F.; Duchateau, J.; Portetelle, D.; Urbain, P.; Van Lint, C.; Green, P.L.; Mahieux, R.; Parmentier, M.; Willems, L.; Kettmann, R. Human T-cell leukemia virus type-1 Tax oncoprotein regulates G-protein signaling. Blood 2007, 109, 1051–1060. [Google Scholar] [CrossRef]
- Kawaguchi, A.; Orba, Y.; Kimura, T.; Iha, H.; Ogata, M.; Tsuji, T.; Ainai, A.; Sata, T.; Okamoto, T.; Hall, W.W.; Sawa, H.; Hasegawa, H. Inhibition of the SDF-1alpha-CXCR4 axis by the CXCR4 antagonist AMD3100 suppresses the migration of cultured cells from ATL patients and murine lymphoblastoid cells from HTLV-I Tax transgenic mice. Blood 2009, 114, 2961–2968. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi, M.; Asao, H.; Hara, T.; Higuchi, M.; Fujii, M.; Nakamura, M. Transcriptional activation of the interleukin-21 gene and its receptor gene by human T-cell leukemia virus type 1 Tax in human T-cells. J. Biol. Chem. 2009, 284, 25501–25511. [Google Scholar] [CrossRef] [PubMed]
- Baba, H.; Yamada, Y.; Mori, N.; Hayashibara, T.; Harasawa, H.; Tsuruda, K.; Sugahara, K.; Soda, H.; Takasaki, Y.; Tawara, M.; et al. Multiple gammac-receptor expression in adult T-cell leukemia. Eur. J. Haematol. 2002, 68, 362–369. [Google Scholar] [CrossRef]
- Ballard, D.W.; Bohnlein, E.; Lowenthal, J.W.; Wano, Y.; Franza, B.R.; Greene, W.C. HTLV-I tax induces cellular proteins that activate the kappa B element in the IL-2 receptor alpha gene. Science 1988, 241, 1652–1655. [Google Scholar] [CrossRef]
- Ruben, S.; Poteat, H.; Tan, T.H.; Kawakami, K.; Roeder, R.; Haseltine, W.; Rosen, C.A. Cellular transcription factors and regulation of IL-2 receptor gene expression by HTLV-I tax gene product. Science 1988, 241, 89–92. [Google Scholar] [CrossRef]
- Hoyos, B.; Ballard, D.W.; Bohnlein, E.; Siekevitz, M.; Greene, W.C. Kappa B-specific DNA binding proteins: Role in the regulation of human interleukin-2 gene expression. Science 1989, 244, 457–460. [Google Scholar] [CrossRef]
- McGuire, K.L.; Curtiss, V.E.; Larson, E.L.; Haseltine, W.A. Influence of human T-cell leukemia virus type I tax and rex on interleukin-2 gene expression. J. Virol. 1993, 67, 1590–1599. [Google Scholar] [CrossRef]
- Good, L.; Maggirwar, S.B.; Sun, S.C. Activation of the IL-2 gene promoter by HTLV-I tax involves induction of NF-AT complexes bound to the CD28-responsive element. EMBO J. 1996, 15, 3744–3750. [Google Scholar] [CrossRef]
- Arya, S.K.; Wong-Staal, F.; Gallo, R.C. T-cell growth factor gene: Lack of expression in human T-cell leukemia-lymphoma virus-infected cells. Science 1984, 223, 1086–1087. [Google Scholar] [CrossRef] [PubMed]
- Dezzutti, C.S.; Rudolph, D.L.; Roberts, C.R.; Lal, R.B. Characterization of human T-lymphotropic virus type I- and II-infected T-cell lines: Antigenic, phenotypic, and genotypic analysis. Virus Res. 1993, 29, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Azimi, N.; Brown, K.; Bamford, R.N.; Tagaya, Y.; Siebenlist, U.; Waldmann, T.A. Human T cell lymphotropic virus type I Tax protein trans-activates interleukin 15 gene transcription through an NF-kappaB site. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 2452–2457. [Google Scholar] [CrossRef] [PubMed]
- Mariner, J.M.; Lantz, V.; Waldmann, T.A.; Azimi, N. Human T cell lymphotropic virus type I Tax activates IL-15R alpha gene expression through an NF-kappa B site. J. Immunol. 2001, 166, 2602–2609. [Google Scholar] [CrossRef] [PubMed]
- Azimi, N.; Jacobson, S.; Leist, T.; Waldmann, T.A. Involvement of IL-15 in the pathogenesis of human T lymphotropic virus type I-associated myelopathy/tropical spastic paraparesis: Implications for therapy with a monoclonal antibody directed to the IL-2/15R beta receptor. J. Immunol. 1999, 163, 4064–4072. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Petrus, M.; Bryant, B.R.; Phuc, N., V; Stamer, M.; Goldman, C.K.; Bamford, R.; Morris, J.C.; Janik, J.E.; Waldmann, T.A. Induction of the IL-9 gene by HTLV-I Tax stimulates the spontaneous proliferation of primary adult T-cell leukemia cells by a paracrine mechanism. Blood 2008, 111, 5163–5172. [Google Scholar] [CrossRef]
- Ju, W.; Zhang, M.; Jiang, J.K.; Thomas, C.J.; Oh, U.; Bryant, B.R.; Chen, J.; Sato, N.; Tagaya, Y.; Morris, J.C.; Janik, J.E.; Jacobson, S.; Waldmann, T.A. CP-690, 550, a therapeutic agent, inhibits cytokine-mediated Jak3 activation and proliferation of T cells from patients with ATL and HAM/TSP. Blood 2011, 117, 1938–1946. [Google Scholar] [CrossRef]
- Chung, H.K.; Young, H.A.; Goon, P.K.; Heidecker, G.; Princler, G.L.; Shimozato, O.; Taylor, G.P.; Bangham, C.R.; Derse, D. Activation of interleukin-13 expression in T cells from HTLV-1-infected individuals and in chronically infected cell lines. Blood 2003, 102, 4130–4136. [Google Scholar] [CrossRef]
- Waldele, K.; Schneider, G.; Ruckes, T.; Grassmann, R. Interleukin-13 overexpression by tax transactivation: A potential autocrine stimulus in human T-cell leukemia virus-infected lymphocytes. J. Virol. 2004, 78, 6081–6090. [Google Scholar] [CrossRef]
- Silbermann, K.; Schneider, G.; Grassmann, R. Stimulation of interleukin-13 expression by human T-cell leukemia virus type 1 oncoprotein Tax via a dually active promoter element responsive to NF-kappaB and NFAT. J. Gen. Virol. 2008, 89, 2788–2798. [Google Scholar] [CrossRef]
- Tanaka, Y.; Fukudome, K.; Hayashi, M.; Takagi, S.; Yoshie, O. Induction of ICAM-1 and LFA-3 by Tax1 of human T-cell leukemia virus type 1 and mechanism of down-regulation of ICAM-1 or LFA-1 in adult-T-cell-leukemia cell lines. Int. J. Cancer. 1995, 60, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Murakami, S.; Oda, S.; Eto, S. Human T-cell leukemia virus type I tax induces intracellular adhesion molecule-1 expression in T cells. Blood 1994, 84, 350–351. [Google Scholar] [CrossRef] [PubMed]
- Valentin, H.; Lemasson, I.; Hamaia, S.; Casse, H.; Konig, S.; Devaux, C.; Gazzolo, L. Transcriptional activation of the vascular cell adhesion molecule-1 gene in T lymphocytes expressing human T-cell leukemia virus type 1 Tax protein. J. Virol. 1997, 71, 8522–8530. [Google Scholar] [CrossRef] [PubMed]
- Valentin, H.; Lemasson, I.; Hamaia, S.; Casse, H.; Konig, S.; Devaux, C.; Gazzolo, L. Transcriptional activation of the vascular cell adhesion molecule-1 gene in T lymphocytes expressing human T-cell leukemia virus type 1 Tax protein. J. Virol. 1997, 71, 8522–8530. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Fukudome, K.; Hayashi, M.; Takagi, S.; Yoshie, O. Induction of ICAM-1 and LFA-3 by Tax1 of human T-cell leukemia virus type 1 and mechanism of down-regulation of ICAM-1 or LFA-1 in adult-T-cell-leukemia cell lines. Int. J. Cancer 1995, 60, 554–561. [Google Scholar] [CrossRef]
- Wake, A.; Tanaka, Y.; Nakatsuka, K.; Misago, M.; Oda, S.; Morimoto, I.; Eto, S. Calcium-dependent homotypic adhesion through leukocyte function-associated antigen-1/intracellular adhesion molecule-1 induces interleukin-1 and parathyroid hormone-related protein production on adult T-cell leukemia cells in vitro. Blood 1995, 86, 2257–2267. [Google Scholar] [CrossRef]
- Wucherpfennig, K.W.; Hollsberg, P.; Richardson, J.H.; Benjamin, D.; Hafler, D.A. T-cell activation by autologous human T-cell leukemia virus type I-infected T-cell clones. Proc. Natl. Acad. Sci. U. S. A. 1992, 89, 2110–2114. [Google Scholar] [CrossRef]
- Nejmeddine, M.; Barnard, A.L.; Tanaka, Y.; Taylor, G.P.; Bangham, C.R. Human T-lymphotropic virus, type 1, tax protein triggers microtubule reorientation in the virological synapse. J. Biol. Chem. 2005, 280, 29653–29660. [Google Scholar] [CrossRef]
- Nejmeddine, M.; Bangham, C.R.M. The HTLV-1 Virological Synapse. Viruses 2010, 2, 1427–1447. [Google Scholar] [CrossRef]
- Banerjee, P.; Feuer, G.; Barker, E. Human T-cell leukemia virus type 1 (HTLV-1) p12I down-modulates ICAM-1 and -2 and reduces adherence of natural killer cells, thereby protecting HTLV-1-infected primary CD4+ T cells from autologous natural killer cell-mediated cytotoxicity despite the reduction of major histocompatibility complex class I molecules on infected cells. J. Virol. 2007, 81, 9707–9717. [Google Scholar] [PubMed]
- Kim, S.J.; Nair, A.M.; Fernandez, S.; Mathes, L.; Lairmore, M.D. Enhancement of LFA-1-mediated T cell adhesion by human T lymphotropic virus type 1 p12I1. J. Immunol. 2006, 176, 5463–5470. [Google Scholar] [CrossRef] [PubMed]
- Van Prooyen, N.; Gold, H.; Andresen, V.; Schwartz, O.; Jones, K.; Ruscetti, F.; Lockett, S.; Gudla, P.; Venzon, D.; Franchini, G. Human T-cell leukemia virus type 1 p8 protein increases cellular conduits and virus transmission. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 20738–20743. [Google Scholar] [CrossRef]
- Malbec, M.; Roesch, F.; Schwartz, O. A New Role for the HTLV-1 p8 Protein: Increasing Intercellular Conduits and Viral Cell-to-Cell Transmission. Viruses 2011, 3, 254–259. [Google Scholar] [CrossRef]
- Sasaki, H.; Nishikata, I.; Shiraga, T.; Akamatsu, E.; Fukami, T.; Hidaka, T.; Kubuki, Y.; Okayama, A.; Hamada, K.; Okabe, H.; et al. Overexpression of a cell adhesion molecule, TSLC1, as a possible molecular marker for acute-type adult T-cell leukemia. Blood 2005, 105, 1204–1213. [Google Scholar] [CrossRef] [PubMed]
- Ghez, D.; Lepelletier, Y.; Jones, K.S.; Pique, C.; Hermine, O. Current concepts regarding the HTLV-1 receptor complex. Retrovirology 2010, 7, 99. [Google Scholar] [CrossRef]
- Matsuoka, M. Human T-cell leukemia virus type I (HTLV-I) infection and the onset of adult T-cell leukemia (ATL). Retrovirology 2005, 2, 27. [Google Scholar] [CrossRef]
- Jacobson, S.; Shida, H.; McFarlin, D.E.; Fauci, A.S.; Koenig, S. Circulating CD8+ cytotoxic T lymphocytes specific for HTLV-I pX in patients with HTLV-I associated neurological disease. Nature 1990, 348, 245–248. [Google Scholar] [CrossRef]
- Kannagi, M.; Harada, S.; Maruyama, I.; Inoko, H.; Igarashi, H.; Kuwashima, G.; Sato, S.; Morita, M.; Kidokoro, M.; Sugimoto, M. Predominant recognition of human T cell leukemia virus type I (HTLV-I) pX gene products by human CD8+ cytotoxic T cells directed against HTLV-I-infected cells. Int. Immunol. 1991, 3, 761–767. [Google Scholar] [CrossRef]
- Nakayama, T.; Hieshima, K.; Arao, T.; Jin, Z.; Nagakubo, D.; Shirakawa, A.K.; Yamada, Y.; Fujii, M.; Oiso, N.; Kawada, A.; et al. Aberrant expression of Fra-2 promotes CCR4 expression and cell proliferation in adult T-cell leukemia. Oncogene 2008, 27, 3221–3232. [Google Scholar] [CrossRef]
- Taylor, J.M.; Nicot, C. HTLV-1 and apoptosis: Role in cellular transformation and recent advances in therapeutic approaches. Apoptosis 2008, 13, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Callens, C.; Moura, I.C.; Lepelletier, Y.; Coulon, S.; Renand, A.; Dussiot, M.; Ghez, D.; Benhamou, M.; Monteiro, R.C.; Bazarbachi, A.; et al. Recent advances in adult T-cell leukemia therapy: focus on a new anti-transferrin receptor monoclonal antibody. Leukemia 2008, 22, 42–48. [Google Scholar] [CrossRef] [PubMed]
- FCS Express, version 3; De Novo Software: Los Angeles, CA, USA.
Appendix

{kind=link}
| Antigen | Conjugate | Isotype | Clone | Origin | Company |
|---|---|---|---|---|---|
| CCR7 | FITC | IgG2a | 150503 | mouse | R&D systems, Wiesbaden, Germany |
| CD25 (IL2RA) | PE | IgG1 | M-A251 | mouse | BD, San Jose, CA, USA |
| CD27 (TNFRSF7) | FITC | IgG1 | O323 | mouse | NatuTec, Frankfurt/M., Germany |
| CD45RO (PTPRC) | FITC | IgG2a | UCHL1 | mouse | NatuTec |
| CD69 | PE | IgG1 | L78 | mouse | BD |
| CD80 | FITC | IgG1 | MEM-233 | mouse | EuroBioSciences, Friesoythe, Germany |
| CD127 (IL7R) | PE | IgG1 | hIL-7R-M21 | mouse | BD |
| CD137 (TNFRSF9/ 4-1BB) | FITC | IgG1 | 4B4-1 | mouse | AbD Serotec, Oxford, UK |
| CD137L (TNFSF9/ 4-1BBL) | R-PE | IgG1 | C65-485 | mouse | BD |
| GITR (TNFRSF 18) | FITC | IgG1 | 110416 | mouse | R&D Systems |
| IgG1 isotype control | R-PE | IgG1 | MOPC-21 | mouse | BD |
| IgG1 isotype control | FITC | IgG1 | MOPC-21 | mouse | BD |
| IgG2a isotype control | FITC | IgG2a | mouse | NatuTec |
Analysis of Surface Expression by Flow Cytometry

| % Expressiona | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Origin | Name | IL2b | PTPRC CD45R0 | CD69 | CD80 B7-1 | TNFRSF18 GITR | TNFRSF7 CD27 | TNFRSF 4-1BB | TNFSF9 4-1BBL | CCR7 EBI1 | IL2RAc CD25 | IL7R CD127 |
| in vitro transformed | C91-PL | no | 0 +/− 0 | 0 +/− 0 | 82 +/− 17 | 41 +/− 3 | 0 +/−0 | 38 +/− 11 | 68 +/− 9 | 73 +/− 42 | 99 +/− 1 | 3 +/− 1 |
| MT-2 | no | 1 +/− 0 | 1 +/− 1 | 84 +/− 15 | 76 +/− 7 | 0 +/− 0 | 39 +/− 9 | 74 +/− 6 | 79 +/− 46 | 99 +/− 0 | 2 +/− 1 | |
| ATLL-derived | HuT-102 | no | 5 +/− 2 | 0 +/− 0 | 88 +/− 11 | 99 +/− 0 | 0 +/− 0 | 65 +/− 5 | 70 +/− 9 | 63 +/− 36 | 100 +/− 0 | 1 +/− 1 |
| ATL-3 | 40 | 96 +/− 1 | NDd | 74 +/− 8c | 72 +/− 9 | 0 +/− 0 | 21 +/− 5 | 19 +/− 6 | 21 +/− 12 | 99 +/− 0 | 2 +/− 2 | |
| Champ | 20 | 88 +/− 5c | 37 +/− 16 | 75 +/− 8c | 83 +/− 8 | 1 +/− 1 | 43 +/− 4 | 31 +/− 5 | 31 +/− 18 | 99 +/− 1 | 3 +/− 1 | |
| JuanaW | 20 | 89 +/− 5 | 45 +/− 13 | 60 +/− 1c | 92 +/− 3 | 0 +/− 0 | 18 +/− 6 | 24 +/− 8 | 55 +/− 32 | 98 +/− 0 | 2 +/− 2 | |
| PaBe | 20 | 96 +/− 3 | 5 +/− 1 | 66 +/− 13 | 39 +/− 0 | 0 +/− 0 | 7 +/− 3 | 21 +/− 3 | 19 +/− 11 | 100 +/− 1 | 3 +/− 2 | |
| StEd | 40 | 70 +/− 5 | 13 +/− 4 | 88 +/− 10 | 79 +/− 1 | 1 +/− 0 | 9 +/− 4 | 16 +/− 1 | 23 +/− 15 | 95 +/− 4 | 2 +/− 2 | |
| HAM/TSP-derived | Abgho | 40 | 95 +/− 1 | 31 +/− 15 | 67 +/− 10c | 71 +/− 10 | 1 +/− 0 | 26 +/− 8 | 18 +/− 4 | 27 +/− 2 | 95 +/− 1 | 4 +/− 2 |
| Eva | 20 | 80 +/− 3 | 28 +/− 9c | 75 +/− 5c | 76 +/− 6 | 1 +/− 0 | 39 +/− 4 | 24 +/− 3 | 38 +/− 2 | 96 +/− 2 | 7 +/− 2 | |
| Nilu | 20 | 72 +/− 3 | 9 +/− 2c | 45 +/− 8c | 41 +/− 4 | 0 +/− 0 | 6 +/− 1 | 12 +/− 3 | 18 +/− 5 | 92 +/− 0 | 1 +/− 0 | |
| Xpos | 20 | 62 +/− 14 | 38 +/− 26c | 60 +/− 5 | 91 +/− 3 | 1 +/− 1 | 31 +/− 3 | 16 +/− 2 | 60 +/− 12 | 99 +/− 1 | 5 +/− 0 | |
| Tax-transformed | Tesi | 40 | 57 +/− 4c | 1 +/− 1c | ND | 93 +/− 2c | 0 +/−0c | 13 +/− 7c | 15 +/− 1 | 9 +/− 1c | 96 +/− 1 | 2 +/− 1c |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kress, A.K.; Grassmann, R.; Fleckenstein, B. Cell Surface Markers in HTLV-1 Pathogenesis. Viruses 2011, 3, 1439-1459. https://doi.org/10.3390/v3081439
Kress AK, Grassmann R, Fleckenstein B. Cell Surface Markers in HTLV-1 Pathogenesis. Viruses. 2011; 3(8):1439-1459. https://doi.org/10.3390/v3081439
Chicago/Turabian StyleKress, Andrea K., Ralph Grassmann, and Bernhard Fleckenstein. 2011. "Cell Surface Markers in HTLV-1 Pathogenesis" Viruses 3, no. 8: 1439-1459. https://doi.org/10.3390/v3081439
APA StyleKress, A. K., Grassmann, R., & Fleckenstein, B. (2011). Cell Surface Markers in HTLV-1 Pathogenesis. Viruses, 3(8), 1439-1459. https://doi.org/10.3390/v3081439