Move or Die: the Fate of the Tax Oncoprotein of HTLV-1

Abstract
1. Introduction
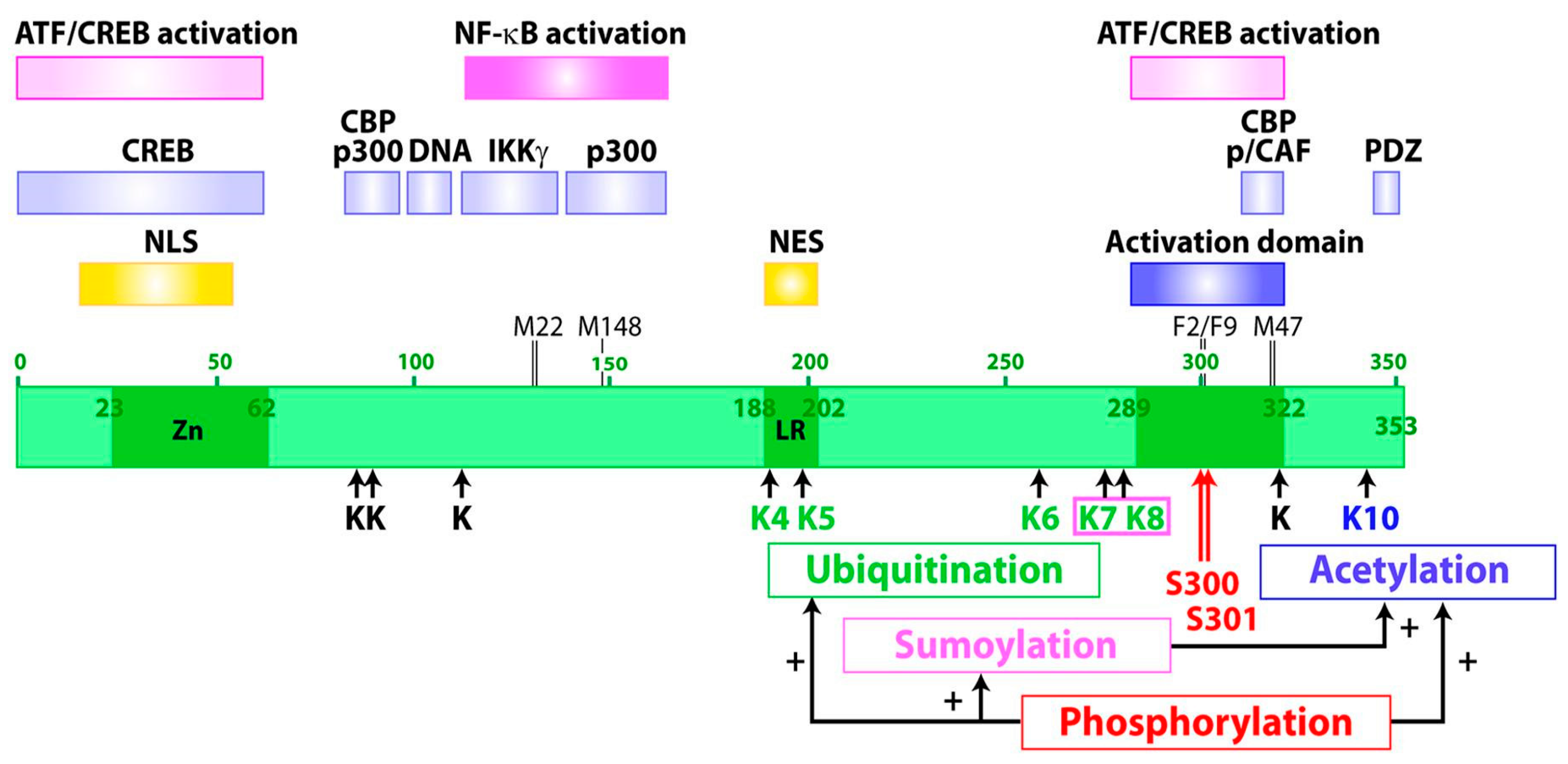
2. Tax Intracellular Distribution and Functional Structure
3. Tax Posttranslational Modifications
3.1. Ubiquitination of Tax
3.2. Sumoylation of Tax
3.3. Proline Isomerization of Tax
3.4. Acetylation of Tax
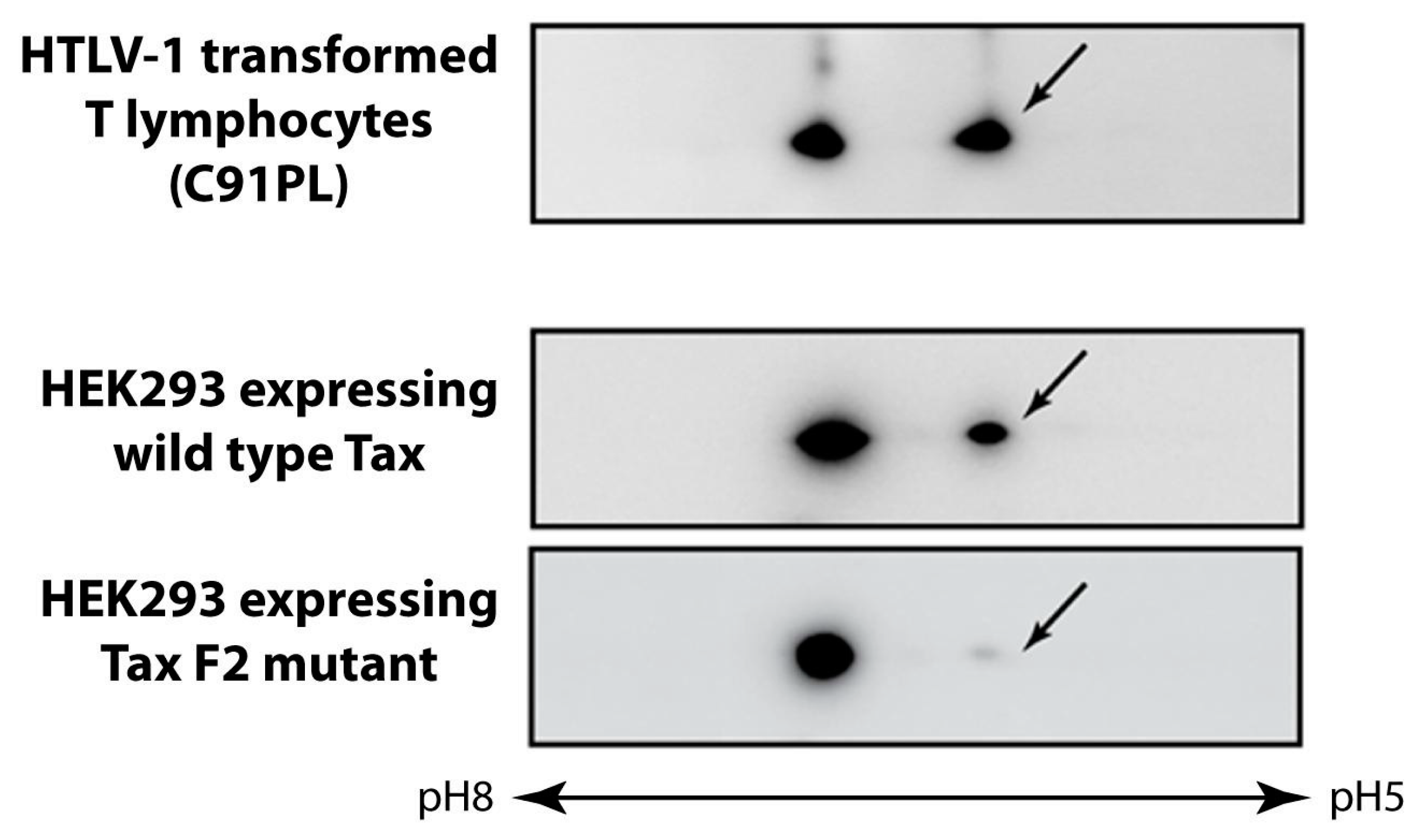
3.5. Phosphorylation of Tax
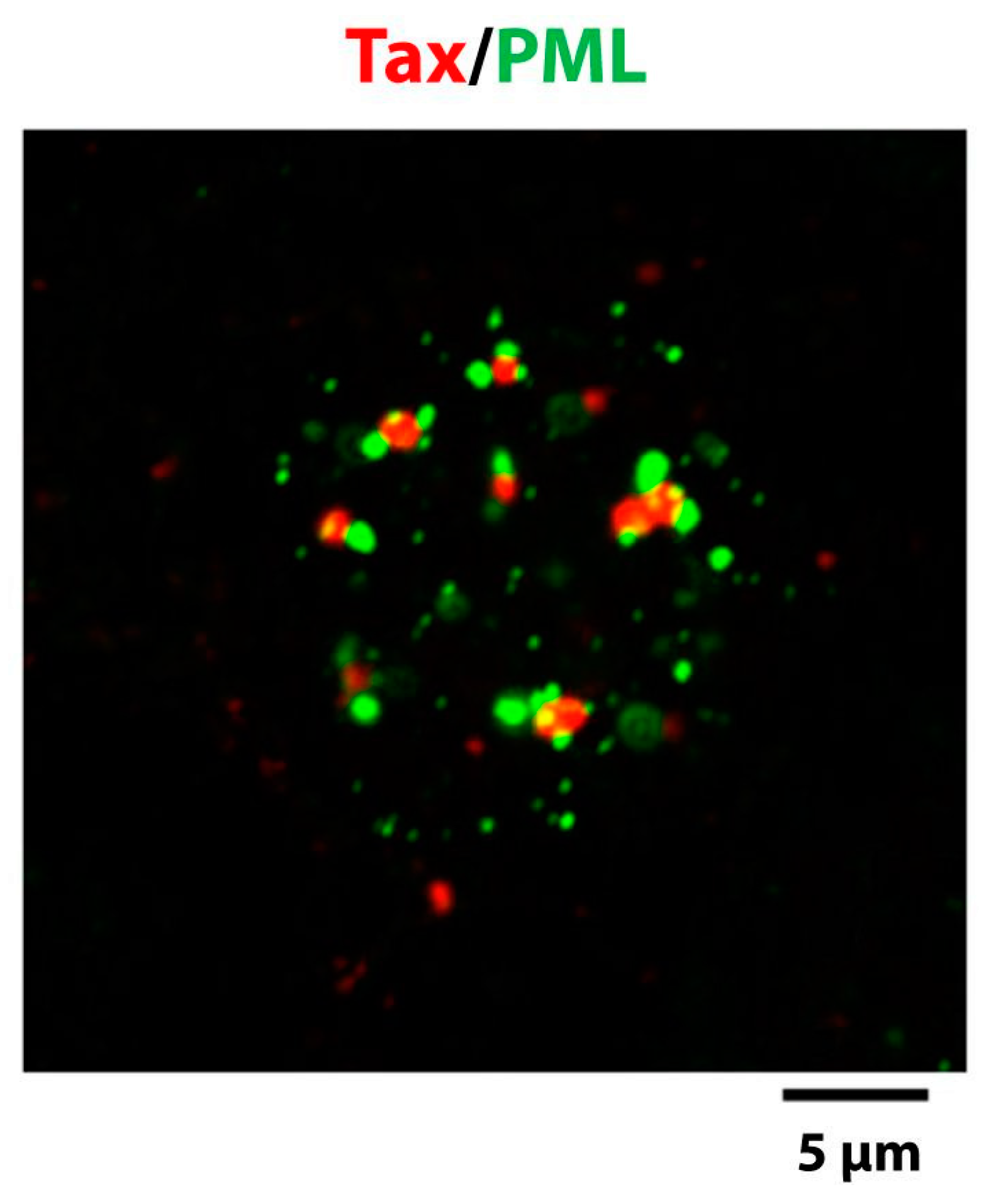
4. Distribution of Tax in the Nucleus
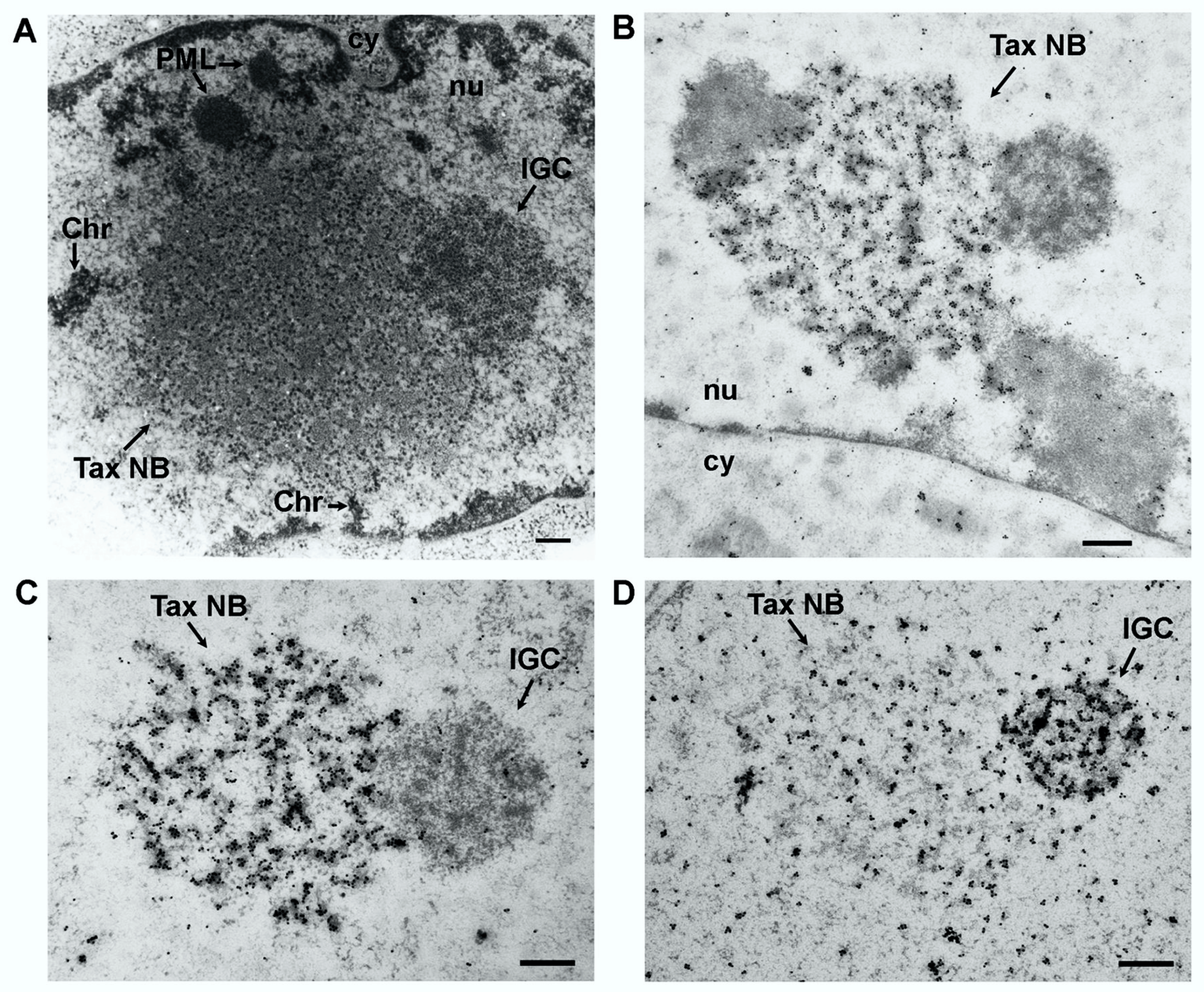
4.1. Structure of the Tax Nuclear Bodies
4.2. Composition of Tax Nuclear Bodies
4.3. Morphogenesis of the Tax Nuclear Bodies
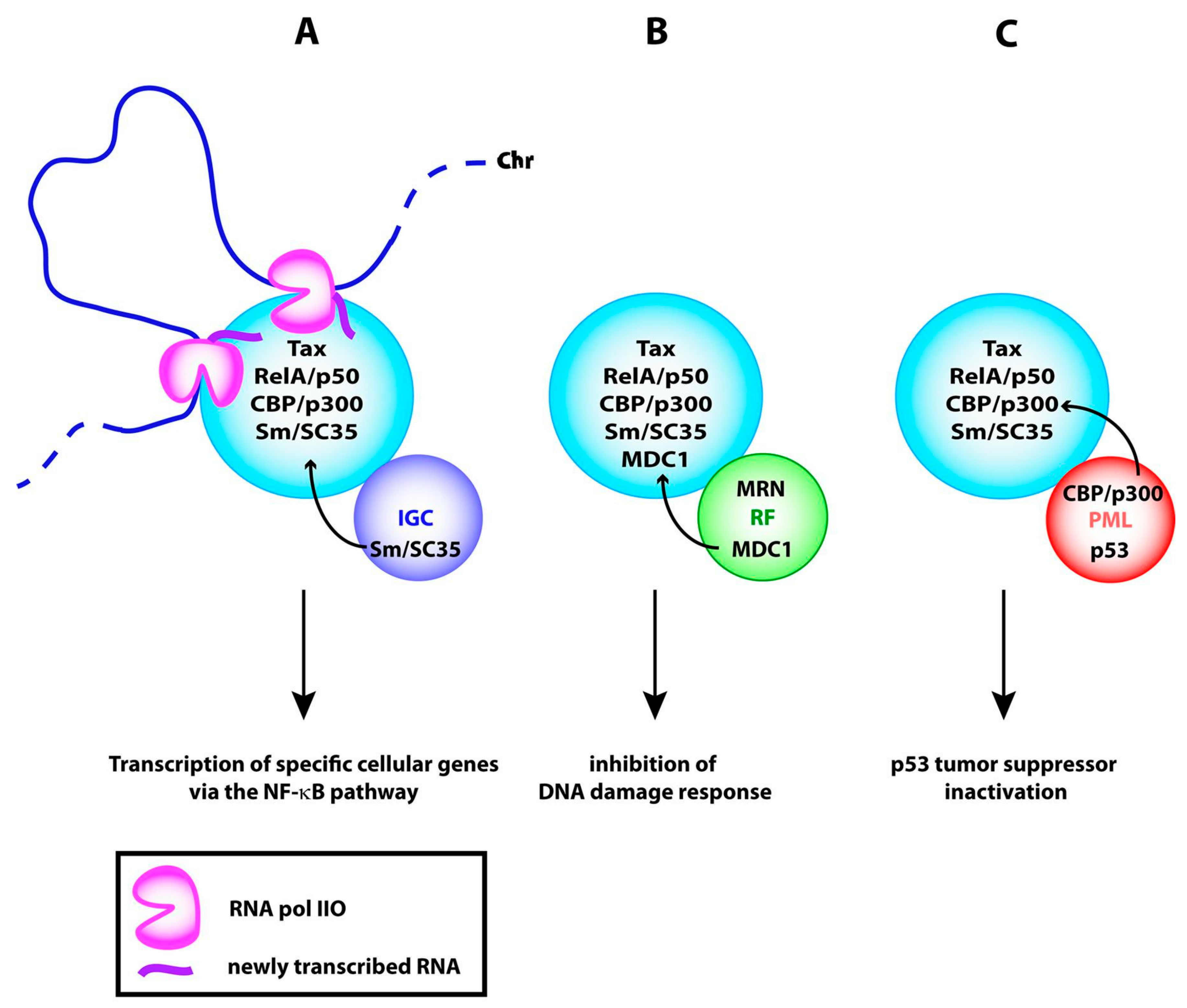
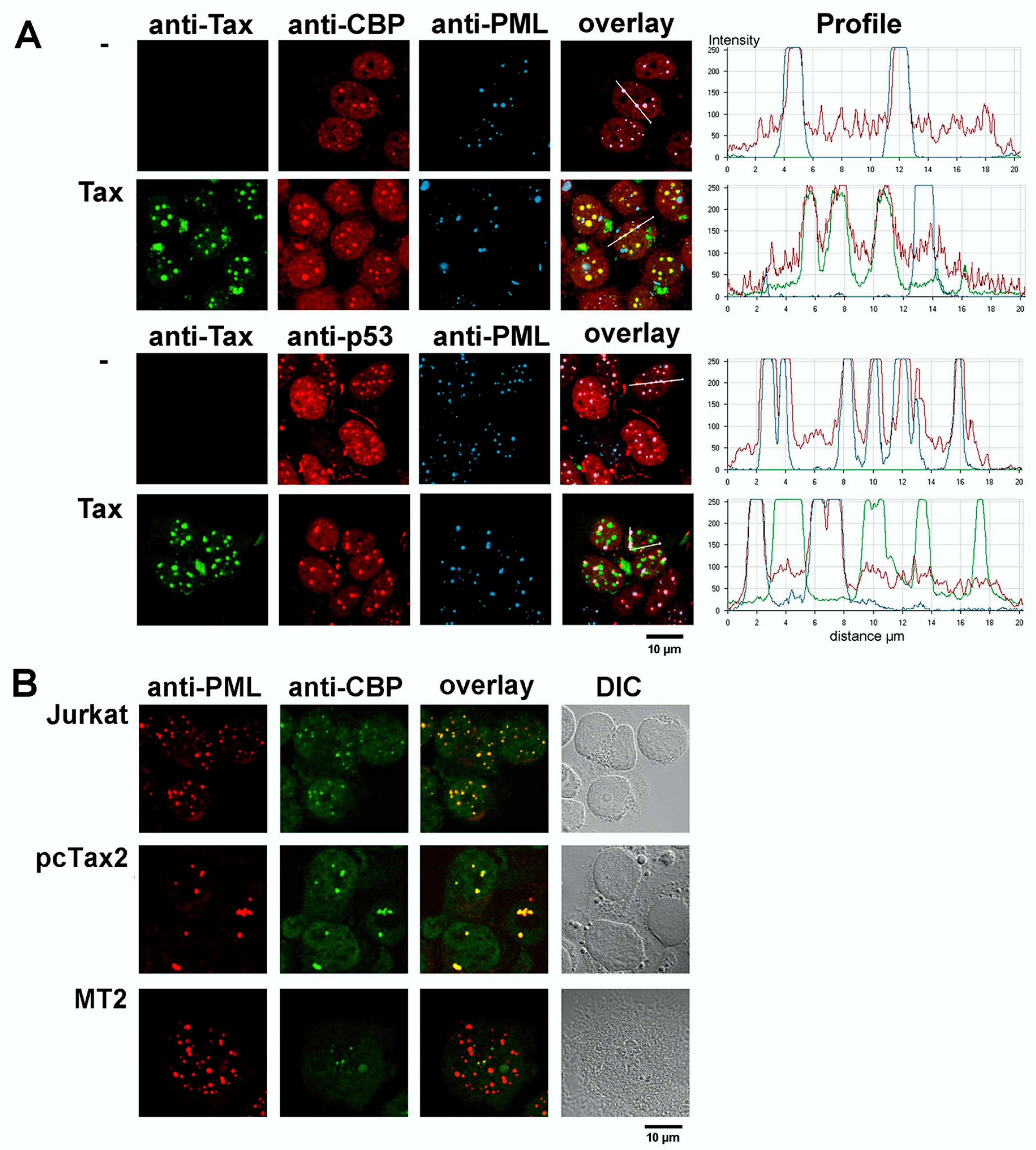
4.4. Function of the Tax Nuclear Bodies
5. Distribution of Tax in the Cytoplasm
6. Migration of Tax in and out of the Nucleus
7. Tax Intracellular Trafficking and Activation of the NF-κB Pathway
8. Conclusions
- Is SUMO/SIM interaction the basis for recruitment of Tax partners to the Tax NBs?
- Do the HTLV-1 provirus and specific cellular genes contact the surface of the Tax NBs in the course of viral and cellular transcription?
- What is the role of IKKγ/NEMO in the Tax NBs?
- How does the crosstalk between Tax NBs and PML NBs impact on prevention of apoptosis by Tax?
- Does calreticulin participate in Tax and IKKγ/NEMO export from the nucleus?
- How does proline isomerization by Pin1 and polyubiquitination by PDLIM2 affect Tax transcriptional and transforming activities?
- Which specific events occur in the cytoplasmic foci that contain Tax, TAB2, TAX1-BP1, IKKγ/NEMO and calreticulin?
- How does assembly of Tax/IKK complexes in the Golgi-associated structures lead to IKK activation?
- Is Tax accumulation in the Golgi involved in its transport to the nucleus?
- Must Tax be demodified in the process of nuclear entry?
- Which kinases phosphorylate Tax in the cytoplasm and in the nucleus?
- A question of central importance is whether Tax-induced genotoxic stress creates a signal in the nucleus that triggers activation of the NF-κB pathway.
Acknowledgements
References and Notes
- Boxus, M.; Twizere, J.C.; Legros, S.; Dewulf, J.F.; Kettmann, R.; Willems, L. The HTLV-1 Tax Interactome. Retrovirology 2008, 5, 76. [Google Scholar] [CrossRef] [PubMed]
- Ng, P.W.; Iha, H.; Iwanaga, Y.; Bittner, M.; Chen, Y.; Jiang, Y.; Gooden, G.; Trent, J.M.; Meltzer, P.; Jeang, K.T.; Zeichner, S.L. Genome-Wide Expression Changes Induced by HTLV-1 Tax: Evidence for MLK-3 Mixed Lineage Kinase Involvement in Tax-Mediated NF-KappaB Activation. Oncogene 2001, 20, 4484–4496. [Google Scholar] [CrossRef] [PubMed]
- Brady, J.; Jeang, K.T.; Duvall, J.; Khoury, G. Identification of P40x-Responsive Regulatory Sequences Within the Human T-Cell Leukemia Virus Type I Long Terminal Repeat. J. Virol. 1987, 61, 2175–2181. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, J.; Seiki, M.; Sato, M.; Yoshida, M. A Transcriptional Enhancer Sequence of HTLV-I Is Responsible for Trans-Activation Mediated by P40 Chi HTLV-I. EMBO J. 1986, 5, 713–718. [Google Scholar] [CrossRef]
- Xu, Y.L.; Adya, N.; Siores, E.; Gao, Q.S.; Giam, C.Z. Cellular Factors Involved in Transcription and Tax-Mediated Trans-Activation Directed by the TGACGT Motifs in Human T-Cell Leukemia Virus Type I Promoter. J. Biol. Chem. 1990, 265, 20285–20292. [Google Scholar] [CrossRef]
- Zhao, L.J.; Giam, C.Z. Human T-Cell Lymphotropic Virus Type I (HTLV-I) Transcriptional Activator, Tax, Enhances CREB Binding to HTLV-I 21-Base-Pair Repeats by Protein-Protein Interaction. Proc. Natl. Acad. Sci. U. S. A. 1992, 89, 7070–7074. [Google Scholar] [CrossRef]
- Tie, F.; Adya, N.; Greene, W.C.; Giam, C.Z. Interaction of the Human T-Lymphotropic Virus Type 1 Tax Dimer With CREB and the Viral 21-Base-Pair Repeat. J. Virol. 1996, 70, 8368–8374. [Google Scholar] [CrossRef]
- Yin, M.J.; Gaynor, R.B. Complex Formation Between CREB and Tax Enhances the Binding Affinity of CREB for the Human T-Cell Leukemia Virus Type 1 21-Base-Pair Repeats. Mol. Cell. Biol. 1996, 16, 3156–3168. [Google Scholar] [CrossRef][Green Version]
- Wagner, S.; Green, M.R. HTLV-I Tax Protein Stimulation of DNA Binding of BZIP Proteins by Enhancing Dimerization. Science 1993, 262, 395–399. [Google Scholar] [CrossRef]
- Kwok, R.P.; Laurance, M.E.; Lundblad, J.R.; Goldman, P.S.; Shih, H.; Connor, L.M.; Marriott, S.J.; Goodman, R.H. Control of CAMP-Regulated Enhancers by the Viral Transactivator Tax Through CREB and the Co-Activator CBP. Nature 1996, 380, 642–646. [Google Scholar] [CrossRef]
- Giebler, H.A.; Loring, J.E.; van Orden, K.; Colgin, M.A.; Garrus, J.E.; Escudero, K.W.; Brauweiler, A.; Nyborg, J.K. Anchoring of CREB Binding Protein to the Human T-Cell Leukemia Virus Type 1 Promoter: a Molecular Mechanism of Tax Transactivation. Mol. Cell. Biol. 1997, 17, 5156–5164. [Google Scholar] [CrossRef] [PubMed]
- Harrod, R.; Tang, Y.; Nicot, C.; Lu, H.S.; Vassilev, A.; Nakatani, Y.; Giam, C.Z. An Exposed KID-Like Domain in Human T-Cell Lymphotropic Virus Type 1 Tax Is Responsible for the Recruitment of Coactivators CBP/P300. Mol. Cell. Biol. 1998, 18, 5052–5061. [Google Scholar] [CrossRef] [PubMed]
- Bex, F.; Yin, M.J.; Burny, A.; Gaynor, R.B. Differential Transcriptional Activation by Human T-Cell Leukemia Virus Type 1 Tax Mutants Is Mediated by Distinct Interactions With CREB Binding Protein and P300. Mol. Cell. Biol. 1998, 18, 2392–2405. [Google Scholar] [CrossRef] [PubMed]
- Lemasson, I.; Polakowski, N.J.; Laybourn, P.J.; Nyborg, J.K. Transcription Factor Binding and Histone Modifications on the Integrated Proviral Promoter in Human T-Cell Leukemia Virus-I-Infected T-Cells. J. Biol. Chem. 2002, 277, 49459–49465. [Google Scholar] [CrossRef]
- Gatza, M.L.; Watt, J.C.; Marriott, S.J. Cellular Transformation by the HTLV-I Tax Protein, a Jack-of-All-Trades. Oncogene 2003, 22, 5141–5149. [Google Scholar] [CrossRef] [PubMed]
- Giam, C.Z.; Jeang, K.T. HTLV-1 Tax and Adult T-Cell Leukemia. Front. Biosci. 2007, 12, 1496–1507. [Google Scholar] [CrossRef] [PubMed]
- Haller, K.; Wu, Y.; Derow, E.; Schmitt, I.; Jeang, K.T.; Grassmann, R. Physical Interaction of Human T-Cell Leukemia Virus Type 1 Tax With Cyclin-Dependent Kinase 4 Stimulates the Phosphorylation of Retinoblastoma Protein. Mol. Cell. Biol. 2002, 22, 3327–3338. [Google Scholar] [CrossRef]
- Liang, M.H.; Geisbert, T.; Yao, Y.; Hinrichs, S.H.; Giam, C.Z. Human T-Lymphotropic Virus Type 1 Oncoprotein Tax Promotes S-Phase Entry but Blocks Mitosis. J. Virol. 2002, 76, 4022–4033. [Google Scholar] [CrossRef]
- Liu, M.; Yang, L.; Zhang, L.; Liu, B.; Merling, R.; Xia, Z.; Giam, C.Z. Human T-Cell Leukemia Virus Type 1 Infection Leads to Arrest in the G1 Phase of the Cell Cycle. J. Virol. 2008, 82, 8442–8455. [Google Scholar] [CrossRef]
- Kuo, Y.L.; Giam, C.Z. Activation of the Anaphase Promoting Complex by HTLV-1 Tax Leads to Senescence. EMBO J. 2006, 25, 1741–1752. [Google Scholar] [CrossRef]
- Lemoine, F.J.; Kao, S.Y.; Marriott, S.J. Suppression of DNA Repair by HTLV Type 1 Tax Correlates With Tax Trans-Activation of Proliferating Cell Nuclear Antigen Gene Expression. AIDS Res. Hum. Retrovir. 2000, 16, 1623–1627. [Google Scholar] [CrossRef] [PubMed]
- Hiscott, J.; Petropoulos, L.; Lacoste, J. Molecular Interactions Between HTLV-1 Tax Protein and the NF-Kappa B/Kappa B Transcription Complex. Virology 1995, 214, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.L.; Shin, Y.A.; Yang, J.M.; DiDonato, J.A.; Ballard, D.W. IKKgamma Mediates the Interaction of Cellular IkappaB Kinases With the Tax Transforming Protein of Human T Cell Leukemia Virus Type 1. J. Biol. Chem. 1999, 274, 15297–15300. [Google Scholar] [CrossRef]
- Harhaj, E.W.; Sun, S.C. IKKgamma Serves As a Docking Subunit of the IkappaB Kinase (IKK) and Mediates Interaction of IKK with the Human T-Cell Leukemia Virus Tax Protein. J. Biol. Chem. 1999, 274, 22911–22914. [Google Scholar] [CrossRef] [PubMed]
- Geleziunas, R.; Ferrell, S.; Lin, X.; Mu, Y.; Cunningham, E.T., Jr.; Grant, M.; Connelly, M.A.; Hambor, J.E.; Marcu, K.B.; Greene, W.C. Human T-Cell Leukemia Virus Type 1 Tax Induction of NF-KappaB Involves Activation of the IkappaB Kinase Alpha (IKKalpha) and IKKbeta Cellular Kinases. Mol. Cell. Biol. 1998, 18, 5157–5165. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.J.; Christerson, L.B.; Yamamoto, Y.; Kwak, Y.T.; Xu, S.; Mercurio, F.; Barbosa, M.; Cobb, M.H.; Gaynor, R.B. HTLV-I Tax Protein Binds to MEKK1 to Stimulate IkappaB Kinase Activity and NF-KappaB Activation. Cell 1998, 93, 875–884. [Google Scholar] [CrossRef]
- Sun, S.C.; Harhaj, E.W.; Xiao, G.; Good, L. Activation of I-KappaB Kinase by the HTLV Type 1 Tax Protein: Mechanistic Insights into the Adaptor Function of IKKgamma. AIDS Res. Hum. Retrovir. 2000, 16, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Gaynor, R.B. Mechanisms of NF-KappaB Activation by the HTLV Type 1 Tax Protein. AIDS Res. Hum. Retrovir. 2000, 16, 1583–1590. [Google Scholar] [CrossRef]
- Smith, M.R.; Greene, W.C. Characterization of a Novel Nuclear Localization Signal in the HTLV-I Tax Transactivator Protein. Virology 1992, 187, 316–320. [Google Scholar] [CrossRef]
- Semmes, O.J.; Jeang, K.T. HTLV-I Tax Is a Zinc-Binding Protein: Role of Zinc in Tax Structure and Function. Virology 1992, 188, 754–764. [Google Scholar] [CrossRef]
- Alefantis, T.; Barmak, K.; Harhaj, E.W.; Grant, C.; Wigdahl, B. Characterization of a Nuclear Export Signal Within the Human T Cell Leukemia Virus Type I Transactivator Protein Tax. J. Biol. Chem. 2003, 278, 21814–21822. [Google Scholar] [CrossRef]
- Lamsoul, I.; Lodewick, J.; Lebrun, S.; Brasseur, R.; Burny, A.; Gaynor, R.B.; Bex, F. Exclusive Ubiquitination and Sumoylation on Overlapping Lysine Residues Mediate NF-KappaB Activation by the Human T-Cell Leukemia Virus Tax Oncoprotein. Mol. Cell. Biol. 2005, 25, 10391–10406. [Google Scholar] [CrossRef] [PubMed]
- Semmes, O.J.; Jeang, K.T. Localization of Human T-Cell Leukemia Virus Type 1 Tax to Subnuclear Compartments That Overlap With Interchromatin Speckles. J. Virol. 1996, 70, 6347–6357. [Google Scholar] [CrossRef]
- Bex, F.; McDowall, A.; Burny, A.; Gaynor, R. The Human T-Cell Leukemia Virus Type 1 Transactivator Protein Tax Colocalizes in Unique Nuclear Structures With NF-KappaB Proteins. J. Virol. 1997, 71, 3484–3497. [Google Scholar] [CrossRef]
- Yin, M.J.; Paulssen, E.J.; Seeler, J.S.; Gaynor, R.B. Protein Domains Involved in Both in Vivo and in Vitro Interactions Between Human T-Cell Leukemia Virus Type I Tax and CREB. J. Virol. 1995, 69, 3420–3432. [Google Scholar] [CrossRef] [PubMed]
- Goren, I.; Semmes, O.J.; Jeang, K.T.; Moelling, K. The Amino Terminus of Tax Is Required for Interaction With the Cyclic AMP Response Element Binding Protein. J. Virol. 1995, 69, 5806–5811. [Google Scholar] [CrossRef] [PubMed]
- Georges, S.A.; Giebler, H.A.; Cole, P.A.; Luger, K.; Laybourn, P.J.; Nyborg, J.K. Tax Recruitment of CBP/P300, Via the KIX Domain, Reveals a Potent Requirement for Acetyltransferase Activity That Is Chromatin Dependent and Histone Tail Independent. Mol. Cell. Biol. 2003, 23, 3392–3404. [Google Scholar] [CrossRef]
- Kimzey, A.L.; Dynan, W.S. Identification of a Human T-Cell Leukemia Virus Type I Tax Peptide in Contact With DNA. J. Biol. Chem. 1999, 274, 34226–34232. [Google Scholar] [CrossRef]
- Semmes, O.J.; Jeang, K.T. Definition of a Minimal Activation Domain in Human T-Cell Leukemia Virus Type I Tax. J. Virol. 1995, 69, 1827–1833. [Google Scholar] [CrossRef]
- Jiang, H.; Lu, H.; Schiltz, R.L.; Pise-Masison, C.A.; Ogryzko, V.V.; Nakatani, Y.; Brady, J.N. PCAF Interacts With Tax and Stimulates Tax Transactivation in a Histone Acetyltransferase-Independent Manner. Mol. Cell. Biol. 1999, 19, 8136–8145. [Google Scholar] [CrossRef]
- Smith, M.R.; Greene, W.C. Identification of HTLV-I Tax Trans-Activator Mutants Exhibiting Novel Transcriptional Phenotypes. Genes Dev. 1990, 4, 1875–1885. [Google Scholar] [CrossRef] [PubMed]
- Lenzmeier, B.A.; Giebler, H.A.; Nyborg, J.K. Human T-Cell Leukemia Virus Type 1 Tax Requires Direct Access to DNA for Recruitment of CREB Binding Protein to the Viral Promoter. Mol. Cell. Biol. 1998, 18, 721–731. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xiao, G.; Harhaj, E.W.; Sun, S.C. Domain-Specific Interaction With the I Kappa B Kinase (IKK) Regulatory Subunit IKK Gamma Is an Essential Step in Tax-Mediated Activation of IKK. J. Biol. Chem. 2000, 275, 34060–34067. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, S.; Inoue, H.; Sakurai, M.; Sugiyama, T.; Hazama, M.; Yamada, T.; Hatanaka, M. Constitutive Activation of NF-Kappa B Is Essential for Transformation of Rat Fibroblasts by the Human T-Cell Leukemia Virus Type I Tax Protein. EMBO J. 1996, 15, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Fryrear, K.A.; Durkin, S.S.; Gupta, S.K.; Tiedebohl, J.B.; Semmes, O.J. Dimerization and a Novel Tax Speckled Structure Localization Signal Are Required for Tax Nuclear Localization. J. Virol. 2009, 83, 5339–5352. [Google Scholar] [CrossRef]
- Basbous, J.; Bazarbachi, A.; Granier, C.; Devaux, C.; Mesnard, J.M. The Central Region of Human T-Cell Leukemia Virus Type 1 Tax Protein Contains Distinct Domains Involved in Subunit Dimerization. J. Virol. 2003, 77, 13028–13035. [Google Scholar] [CrossRef]
- Jin, D.Y.; Jeang, K.T. HTLV-I Tax Self-Association in Optimal Trans-Activation Function. Nucleic Acids Res. 1997, 25, 379–387. [Google Scholar] [CrossRef]
- Rousset, R.; Fabre, S.; Desbois, C.; Bantignies, F.; Jalinot, P. The C-Terminus of the HTLV-1 Tax Oncoprotein Mediates Interaction With the PDZ Domain of Cellular Proteins. Oncogene 1998, 16, 643–654. [Google Scholar] [CrossRef]
- Peloponese, J.M., Jr.; Iha, H.; Yedavalli, V.R.; Miyazato, A.; Li, Y.; Haller, K.; Benkirane, M.; Jeang, K.T. Ubiquitination of Human T-Cell Leukemia Virus Type 1 Tax Modulates Its Activity. J. Virol. 2004, 78, 11686–11695. [Google Scholar] [CrossRef]
- Chiari, E.; Lamsoul, I.; Lodewick, J.; Chopin, C.; Bex, F.; Pique, C. Stable Ubiquitination of Human T-Cell Leukemia Virus Type 1 Tax Is Required for Proteasome Binding. J. Virol. 2004, 78, 11823–11832. [Google Scholar] [CrossRef]
- Nasr, R.; Chiari, E.; El Sabban, M.; Mahieux, R.; Kfoury, Y.; Abdulhay, M.; Yazbeck, V.; Hermine, O.; de The, H.; Pique, C.; Bazarbachi, A. Tax Ubiquitylation and Sumoylation Control Critical Cytoplasmic and Nuclear Steps of NF-Kappa B Activation. Blood 2006, 107, 4021–4029. [Google Scholar] [CrossRef] [PubMed]
- Hemelaar, J.; Bex, F.; Booth, B.; Cerundolo, V.; McMichael, A.; Daenke, S. Human T-Cell Leukemia Virus Type 1 Tax Protein Binds to Assembled Nuclear Proteasomes and Enhances Their Proteolytic Activity. J. Virol. 2001, 75, 11106–11115. [Google Scholar] [CrossRef] [PubMed]
- Shembade, N.; Harhaj, N.S.; Yamamoto, M.; Akira, S.; Harhaj, E.W. The Human T-Cell Leukemia Virus Type 1 Tax Oncoprotein Requires the Ubiquitin-Conjugating Enzyme Ubc13 for NF-KappaB Activation. J. Virol. 2007, 81, 13735–13742. [Google Scholar] [CrossRef]
- Huang, J.; Ren, T.; Guan, H.; Jiang, Y.; Cheng, H. HTLV-1 Tax Is a Critical Lipid Raft Modulator That Hijacks IkappaB Kinases to the Microdomains for Persistent Activation of NF-KappaB. J. Biol. Chem. 2009, 284, 6208–6217. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Fu, J.; Qu, Z.; Li, S.; Tanaka, T.; Grusby, M. J.; Xiao, G. PDLIM2 Suppresses Human T-Cell Leukemia Virus Type I Tax-Mediated Tumorigenesis by Targeting Tax into the Nuclear Matrix for Proteasomal Degradation. Blood 2009, 113, 4370–4380. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Grusby, M.J.; Kaisho, T. PDLIM2-Mediated Termination of Transcription Factor NF-KappaB Activation by Intranuclear Sequestration and Degradation of the P65 Subunit. Nat. Immunol. 2007, 8, 584–591. [Google Scholar] [CrossRef]
- Fu, J.; Yan, P.; Li, S.; Qu, Z.; Xiao, G. Molecular Determinants of PDLIM2 in Suppressing HTLV-I Tax-Mediated Tumorigenesis. Oncogene 2010, 29, 6499–6507. [Google Scholar] [CrossRef]
- Gatza, M.L.; Dayaram, T.; Marriott, S.J. Ubiquitination of HTLV-I Tax in Response to DNA Damage Regulates Nuclear Complex Formation and Nuclear Export. Retrovirology 2007, 4, 95. [Google Scholar] [CrossRef]
- Kfoury, Y.; Nasr, R.; Favre-Bonvin, A.; El Sabban, M.; Renault, N.; Giron, M.L.; Setterblad, N.; Hajj, H.E.; Chiari, E.; Mikati, A.G.; Hermine, O.; Saib, A.; de The, H.; Pique, C.; Bazarbachi, A. Ubiquitylated Tax Targets and Binds the IKK Signalosome at the Centrosome. Oncogene 2008, 27, 1665–1676. [Google Scholar] [CrossRef]
- Peloponese, J.M.; Jeang, K.; Kinjo, T.; Junichiro, Y. PIN1 Interacts and Modulates Tax's Activation of NF-KB. AIDS Res. Hum. Retrovir. 2009, 25, 1291. [Google Scholar]
- Jeong, S.J.; Ryo, A.; Yamamoto, N. The Prolyl Isomerase Pin1 Stabilizes the Human T-Cell Leukemia Virus Type 1 (HTLV-1) Tax Oncoprotein and Promotes Malignant Transformation. Biochem. Biophys. Res. Comm. 2009, 381, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Ryo, A.; Suizu, F.; Yoshida, Y.; Perrem, K.; Liou, Y.C.; Wulf, G.; Rottapel, R.; Yamaoka, S.; Lu, K.P. Regulation of NF-KappaB Signaling by Pin1-Dependent Prolyl Isomerization and Ubiquitin-Mediated Proteolysis of P65/RelA. Mol. Cell 2003, 12, 1413–1426. [Google Scholar] [CrossRef] [PubMed]
- Wulf, G.M.; Liou, Y.C.; Ryo, A.; Lee, S.W.; Lu, K.P. Role of Pin1 in the Regulation of P53 Stability and P21 Transactivation, and Cell Cycle Checkpoints in Response to DNA Damage. J. Biol. Chem. 2002, 277, 47976–47979. [Google Scholar] [CrossRef] [PubMed]
- Deronne, B.; Sampaio, C.; Roger, P.; Bex, F. Involvement of proline-directed kinase CDK4 and peptidyl propyl isomerase Pin1 in HTLV-1 Tax transcriptional activity. Université Libre de Bruxelles. Unpublished work. 2011. [Google Scholar]
- Lodewick, J.; Lamsoul, I.; Polania, A.; Lebrun, S.; Burny, A.; Ratner, L.; Bex, F. Acetylation of the Human T-Cell Leukemia Virus Type 1 Tax Oncoprotein by P300 Promotes Activation of the NF-Kappa B Pathway. Virology 2009, 386, 68–78. [Google Scholar] [CrossRef]
- Lodewick, J.; Coulonval, K.; Roger, P.; Bex, F. Two-dimensional gel electrophoresis reveals phosphorylated and acetylated forms of the HTLV-1 Tax oncoprotein in HTLV-1 infected T lympbocytes. 2011; to be submitted for publication. [Google Scholar]
- Bex, F.; Murphy, K.; Wattiez, R.; Burny, A.; Gaynor, R.B. Phosphorylation of the Human T-Cell Leukemia Virus Type 1 Transactivator Tax on Adjacent Serine Residues Is Critical for Tax Activation. J. Virol. 1999, 73, 738–745. [Google Scholar] [CrossRef]
- Durkin, S.S.; Ward, M.D.; Fryrear, K.A.; Semmes, O.J. Site-Specific Phosphorylation Differentiates Active From Inactive Forms of the Human T-Cell Leukemia Virus Type 1 Tax Oncoprotein. J. Biol. Chem. 2006, 281, 31705–31712. [Google Scholar] [CrossRef]
- Bidoia, C.; Mazzorana, M.; Pagano, M.A.; Arrigoni, G.; Meggio, F.; Pinna, L.A.; Bertazzoni, U. The Pleiotropic Protein Kinase CK2 Phosphorylates HTLV-1 Tax Protein in Vitro, Targeting Its PDZ-Binding Motif. Virus Genes 2010, 41, 149–157. [Google Scholar] [CrossRef]
- Baydoun, H.; Duc-Dodon, M.; Lebrun, S.; Gazzolo, L.; Bex, F. Regulation of the Human T-Cell Leukemia Virus Gene Expression Depends on the Localization of Regulatory Proteins Tax, Rex and P30II in Specific Nuclear Subdomains. Gene 2007, 386, 191–201. [Google Scholar] [CrossRef]
- Huang, S.; Spector, D.L. Nascent Pre-MRNA Transcripts Are Associated With Nuclear Regions Enriched in Splicing Factors. Genes Dev. 1991, 5, 2288–2302. [Google Scholar] [CrossRef]
- Misteli, T.; Caceres, J.F.; Clement, J.Q.; Krainer, A.R.; Wilkinson, M.F.; Spector, D.L. Serine Phosphorylation of SR Proteins Is Required for Their Recruitment to Sites of Transcription in vivo. J. Cell Biol. 1998, 143, 297–307. [Google Scholar] [CrossRef]
- Spector, D.L.; Lamond, A.I. Nuclear Speckles. Cold Spring Harb. Perspect. Biol. 2011, 3, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Ariumi, Y.; Ego, T.; Kaida, A.; Matsumoto, M.; Pandolfi, P.P.; Shimotohno, K. Distinct Nuclear Body Components, PML and SMRT, Regulate the Trans-Acting Function of HTLV-1 Tax Oncoprotein. Oncogene 2003, 22, 1611–1619. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lallemand-Breitenbach, V.; de The, H. PML Nuclear Bodies. Cold Spring Harb. Perspect. Biol. 2010, 2, a000661. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Matunis, M.J.; Dejean, A. Conjugation With the Ubiquitin-Related Modifier SUMO-1 Regulates the Partitioning of PML Within the Nucleus. EMBO J. 1998, 17, 61–70. [Google Scholar] [CrossRef]
- Seeler, J.S.; Dejean, A. SUMO: of Branched Proteins and Nuclear Bodies. Oncogene 2001, 20, 7243–7249. [Google Scholar] [CrossRef]
- de, l., V; Frobius, K.; Moreno, R.; Calzado, M.A.; Geng, H.; Schmitz, M.L. Control of Nuclear HIPK2 Localization and Function by a SUMO Interaction Motif. Biochim. Biophys. Acta 2011, 1813, 283–297. [Google Scholar]
- Sung, K.S.; Lee, Y.A.; Kim, E.T.; Lee, S.R.; Ahn, J.H.; Choi, C.Y. Role of the SUMO-Interacting Motif in HIPK2 Targeting to the PML Nuclear Bodies and Regulation of P53. Exp. Cell Res. 2011, 317, 1060–1070. [Google Scholar] [CrossRef]
- Weidtkamp-Peters, S.; Lenser, T.; Negorev, D.; Gerstner, N.; Hofmann, T.G.; Schwanitz, G.; Hoischen, C.; Maul, G.; Dittrich, P.; Hemmerich, P. Dynamics of Component Exchange at PML Nuclear Bodies. J. Cell Sci. 2008, 121, 2731–2743. [Google Scholar] [CrossRef]
- Avesani, F.; Romanelli, M.G.; Turci, M.; di Gennaro, G.; Sampaio, C.; Bidoia, C.; Bertazzoni, U.; Bex, F. Association of HTLV Tax Proteins With TAK1-Binding Protein 2 and RelA in Calreticulin-Containing Cytoplasmic Structures Participates in Tax-Mediated NF-KappaB Activation. Virology 2010, 408, 39–48. [Google Scholar] [CrossRef]
- Kfoury, Y.; Setterblad, N.; El Sabban, M.; Zamborlini, A.; Dassouki, Z.; El Hajj, H.; Hermine, O.; Pique, C.; de The, H.; Saib, A.; Bazarbachi, A. Tax Ubiquitylation and SUMOylation Control the Dynamic Shuttling of Tax and NEMO Between Ubc9 Nuclear Bodies and the Centrosome. Blood 2011, 117, 190–199. [Google Scholar] [CrossRef]
- Suzuki, T.; Hirai, H.; Yoshida, M. Tax Protein of HTLV-1 Interacts With the Rel Homology Domain of NF-Kappa B P65 and C-Rel Proteins Bound to the NF-Kappa B Binding Site and Activates Transcription. Oncogene 1994, 9, 3099–3105. [Google Scholar] [PubMed]
- Suzuki, T.; Hirai, H.; Fujisawa, J.; Fujita, T.; Yoshida, M.A. Trans-Activator Tax of Human T-Cell Leukemia Virus Type 1 Binds to NF-Kappa B P50 and Serum Response Factor (SRF) and Associates with Enhancer DNAs of the NF-Kappa B Site and CArG Box. Oncogene 1993, 8, 2391–2397. [Google Scholar] [PubMed]
- Harhaj, E.W.; Good, L.; Xiao, G.; Uhlik, M.; Cvijic, M. E.; Rivera-Walsh, I.; Sun, S.C. Somatic Mutagenesis Studies of NF-Kappa B Signaling in Human T Cells: Evidence for an Essential Role of IKK Gamma in NF-Kappa B Activation by T-Cell Costimulatory Signals and HTLV-I Tax Protein. Oncogene 2000, 19, 1448–1456. [Google Scholar] [CrossRef] [PubMed]
- Krappmann, D.; Hatada, E.N.; Tegethoff, S.; Li, J.; Klippel, A.; Giese, K.; Baeuerle, P.A.; Scheidereit, C. The I Kappa B Kinase (IKK) Complex Is Tripartite and Contains IKK Gamma but Not IKAP As a Regular Component. J. Biol. Chem. 2000, 275, 29779–29787. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, G.J.; Zhang, Z.Q.; Jin, D.Y. Stimulation of IKK-Gamma Oligomerization by the Human T-Cell Leukemia Virus Oncoprotein Tax. FEBS Lett. 2002, 531, 494–498. [Google Scholar] [CrossRef]
- Yan, J.P.; Garrus, J.E.; Giebler, H.A.; Stargell, L.A.; Nyborg, J.K. Molecular Interactions Between the Coactivator CBP and the Human T-Cell Leukemia Virus Tax Protein. J. Mol. Biol. 1998, 281, 395–400. [Google Scholar] [CrossRef][Green Version]
- Geiger, T.R.; Sharma, N.; Kim, Y.M.; Nyborg, J.K. The Human T-Cell Leukemia Virus Type 1 Tax Protein Confers CBP/P300 Recruitment and Transcriptional Activation Properties to Phosphorylated CREB. Mol. Cell. Biol. 2008, 28, 1383–1392. [Google Scholar] [CrossRef]
- Nizet, S. The Tax oncoprotein of HTLV-1 assembles unique nuclear structures involved in transcriptional activation. PHD Thesis, Université Libre de Bruxelles, 2001. [Google Scholar]
- Belgnaoui, S.M.; Fryrear, K.A.; Nyalwidhe, J.O.; Guo, X.; Semmes, O.J. The Viral Oncoprotein Tax Sequesters DNA Damage Response Factors by Tethering MDC1 to Chromatin. J. Biol. Chem. 2010, 285, 32897–32905. [Google Scholar] [CrossRef]
- Haoudi, A.; Daniels, R.C.; Wong, E.; Kupfer, G.; Semmes, O.J. Human T-Cell Leukemia Virus-I Tax Oncoprotein Functionally Targets a Subnuclear Complex Involved in Cellular DNA Damage-Response. J. Biol. Chem. 2003, 278, 37736–37744. [Google Scholar] [CrossRef]
- Gupta, S.K.; Guo, X.; Durkin, S.S.; Fryrear, K.F.; Ward, M.D.; Semmes, O.J. Human T-Cell Leukemia Virus Type 1 Tax Oncoprotein Prevents DNA Damage-Induced Chromatin Egress of Hyperphosphorylated Chk2. J. Biol. Chem. 2007, 282, 29431–29440. [Google Scholar] [CrossRef]
- Kinjo, T.; Ham-Terhune, J.; Peloponese, J.M., Jr.; Jeang, K.T. Induction of Reactive Oxygen Species by Human T-Cell Leukemia Virus Type 1 Tax Correlates With DNA Damage and Expression of Cellular Senescence Marker. J. Virol. 2010, 84, 5431–5437. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liang, M.H.; Kuo, Y.L.; Liao, W.; Boros, I.; Kleinberger, T.; Blancato, J.; Giam, C.Z. Human T-Lymphotropic Virus Type 1 Oncoprotein Tax Promotes Unscheduled Degradation of Pds1p/Securin and Clb2p/Cyclin B1 and Causes Chromosomal Instability. Mol. Cell. Biol. 2003, 23, 5269–5281. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mihaylova, V.T.; Green, A.M.; Khurgel, M.; Semmes, O.J.; Kupfer, G.M. Human T-Cell Leukemia Virus I Tax Protein Sensitizes P53-Mutant Cells to DNA Damage. Cancer Res. 2008, 68, 4843–4852. [Google Scholar] [CrossRef] [PubMed]
- Majone, F.; Luisetto, R.; Zamboni, D.; Iwanaga, Y.; Jeang, K.T. Ku Protein As a Potential Human T-Cell Leukemia Virus Type 1 (HTLV-1) Tax Target in Clastogenic Chromosomal Instability of Mammalian Cells. Retrovirology 2005, 2, 45. [Google Scholar] [CrossRef] [PubMed]
- Marriott, S.J.; Semmes, O.J. Impact of HTLV-I Tax on Cell Cycle Progression and the Cellular DNA Damage Repair Response. Oncogene 2005, 24, 5986–5995. [Google Scholar] [CrossRef]
- Chandhasin, C.; Ducu, R.I.; Berkovich, E.; Kastan, M.B.; Marriott, S.J. Human T-Cell Leukemia Virus Type 1 Tax Attenuates the ATM-Mediated Cellular DNA Damage Response. J. Virol. 2008, 82, 6952–6961. [Google Scholar] [CrossRef]
- Durkin, S.S.; Guo, X.; Fryrear, K.A.; Mihaylova, V.T.; Gupta, S.K.; Belgnaoui, S.M.; Haoudi, A.; Kupfer, G.M.; Semmes, O.J. HTLV-1 Tax Oncoprotein Subverts the Cellular DNA Damage Response Via Binding to DNA-Dependent Protein Kinase. J. Biol. Chem. 2008, 283, 36311–36320. [Google Scholar] [CrossRef]
- Hay, R.T. SUMO: A History of Modification. Mol. Cell 2005, 18, 1–12. [Google Scholar] [CrossRef]
- Lin, D.Y.; Huang, Y.S.; Jeng, J.C.; Kuo, H.Y.; Chang, C.C.; Chao, T.T.; Ho, C.C.; Chen, Y.C.; Lin, T.P.; Fang, H.I.; Hung, C.C.; Suen, C.S.; Hwang, M.J.; Chang, K.S.; Maul, G.G.; Shih, H.M. Role of SUMO-Interacting Motif in Daxx SUMO Modification, Subnuclear Localization, and Repression of Sumoylated Transcription Factors. Mol. Cell 2006, 24, 341–354. [Google Scholar] [CrossRef]
- Shen, T.H.; Lin, H.K.; Scaglioni, P.P.; Yung, T.M.; Pandolfi, P.P. The Mechanisms of PML-Nuclear Body Formation. Mol. Cell 2006, 24, 331–339. [Google Scholar] [CrossRef]
- Matera, A.G.; Izaguire-Sierra, M.; Praveen, K.; Rajendra, T.K. Nuclear Bodies: Random Aggregates of Sticky Proteins or Crucibles of Macromolecular Assembly? Dev. Cell 2009, 17, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Marenduzzo, D.; Faro-Trindade, I.; Cook, P.R. What Are the Molecular Ties That Maintain Genomic Loops? Trends Genet. 2007, 23, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Stein, G.S.; Stein, J.L.; van Wijnen, A.J.; Lian, J.B.; Zaidi, S.K.; Nickerson, J.A.; Montecino, M.A.; Young, D.W. An Architectural Genetic and Epigenetic Perspective. Integr. Biol. (Camb.) 2011, 3, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, F.M.; Kruhlak, M.J.; Box, A.K.; Hendzel, M.J.; Bazett-Jones, D.P. The Transcription Coactivator CBP Is a Dynamic Component of the Promyelocytic Leukemia Nuclear Body. J. Cell Biol. 2001, 152, 1099–1106. [Google Scholar] [CrossRef]
- D'Orazi, G.; Cecchinelli, B.; Bruno, T.; Manni, I.; Higashimoto, Y.; Saito, S.; Gostissa, M.; Coen, S.; Marchetti, A.; Del Sal, G.; Piaggio, G.; Fanciulli, M.; Appella, E.; Soddu, S. Homeodomain-Interacting Protein Kinase-2 Phosphorylates P53 at Ser 46 and Mediates Apoptosis. Nat. Cell Biol. 2002, 4, 11–19. [Google Scholar] [CrossRef]
- Louria-Hayon, I.; Grossman, T.; Sionov, R.V.; Alsheich, O.; Pandolfi, P.P.; Haupt, Y. PML Protects P53 From Mdm2-Mediated Inhibition and DegradationB. J. Biol. Chem. 2003, 278, 33134–33141. [Google Scholar] [CrossRef]
- Hofmann, T.G.; Moller, A.; Sirma, H.; Zentgraf, H.; Taya, Y.; Droge, W.; Will, H.; Schmitz, M.L. Regulation of P53 Activity by Its Interaction With Homeodomain-Interacting Protein Kinase-2. Nat. Cell Biol. 2002, 4, 1–10. [Google Scholar] [CrossRef]
- Krieghoff-Henning, E.; Hofmann, T.G. Role of Nuclear Bodies in Apoptosis Signalling. Biochim. Biophys. Acta 2008, 1783, 2185–2194. [Google Scholar] [CrossRef]
- Moller, A.; Sirma, H.; Hofmann, T.G.; Rueffer, S.; Klimczak, E.; Droge, W.; Will, H.; Schmitz, M.L. PML Is Required for Homeodomain-Interacting Protein Kinase 2 (HIPK2)-Mediated P53 Phosphorylation and Cell Cycle Arrest but Is Dispensable for the Formation of HIPK Domains. Cancer Res. 2003, 63, 4310–4314. [Google Scholar]
- Pearson, M.; Carbone, R.; Sebastiani, C.; Cioce, M.; Fagioli, M.; Saito, S.; Higashimoto, Y.; Appella, E.; Minucci, S.; Pandolfi, P.P.; Pelicci, P.G. PML Regulates P53 Acetylation and Premature Senescence Induced by Oncogenic Ras. Nature 2000, 406, 207–210. [Google Scholar] [CrossRef]
- Pearson, M.; Pelicci, P.G. PML Interaction with P53 and Its Role in Apoptosis and Replicative Senescence. Oncogene 2001, 20, 7250–7256. [Google Scholar] [CrossRef] [PubMed]
- Fogal, V.; Gostissa, M.; Sandy, P.; Zacchi, P.; Sternsdorf, T.; Jensen, K.; Pandolfi, P.P.; Will, H.; Schneider, C.; Del Sal, G. Regulation of P53 Activity in Nuclear Bodies by a Specific PML Isoform. EMBO J. 2000, 19, 6185–6195. [Google Scholar] [CrossRef] [PubMed]
- Ferbeyre, G.; de Stanchina, E.; Querido, E.; Baptiste, N.; Prives, C.; Lowe, S.W. PML Is Induced by Oncogenic Ras and Promotes Premature Senescence. Genes Dev. 2000, 14, 2015–2027. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Salomoni, P.; Luo, J.; Shih, A.; Zhong, S.; Gu, W.; Paolo, P.P. The Function of PML in P53-Dependent Apoptosis. Nat. Cell Biol. 2000, 2, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Bischof, O.; Kirsh, O.; Pearson, M.; Itahana, K.; Pelicci, P.G.; Dejean, A. Deconstructing PML-Induced Premature Senescence. EMBO J. 2002, 21, 3358–3369. [Google Scholar] [CrossRef]
- Robek, M.D.; Ratner, L. Immortalization of CD4(+) and CD8(+) T Lymphocytes by Human T-Cell Leukemia Virus Type 1 Tax Mutants Expressed in a Functional Molecular Clone. J. Virol. 1999, 73, 4856–4865. [Google Scholar] [CrossRef] [PubMed]
- Nejmeddine, M.; Barnard, A.L.; Tanaka, Y.; Taylor, G.P.; Bangham, C.R. HTLV-1 Tax Protein Triggers Microtubule Reorientation in the Virological Synapse. J. Biol. Chem. 2005, 280, 29653–29660. [Google Scholar] [CrossRef]
- Harhaj, N.S.; Sun, S.C.; Harhaj, E.W. Activation of NF-Kappa B by the Human T Cell Leukemia Virus Type I Tax Oncoprotein Is Associated With Ubiquitin-Dependent Relocalization of I Kappa B Kinase. J. Biol. Chem. 2007, 282, 4185–4192. [Google Scholar] [CrossRef]
- Alefantis, T.; Flaig, K.E.; Wigdahl, B.; Jain, P. Interaction of HTLV-1 Tax Protein with Calreticulin: Implications for Tax Nuclear Export and Secretion. Biomed. Pharmacother. 2007, 61, 194–200. [Google Scholar] [CrossRef]
- Tsuji, T.; Sheehy, N.; Gautier, V.W.; Hayakawa, H.; Sawa, H.; Hall, W.W. The Nuclear Import of the Human T Lymphotropic Virus Type I (HTLV-1) Tax Protein Is Carrier- and Energy-Independent. J. Biol. Chem. 2007, 282, 13875–13883. [Google Scholar] [CrossRef]
- Gatza, M.L.; Marriott, S.J. Genotoxic Stress and Cellular Stress Alter the Subcellular Distribution of Human T-Cell Leukemia Virus Type 1 Tax Through a CRM1-Dependent Mechanism. J. Virol. 2006, 80, 6657–6668. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Groenendyk, J.; Szabo, E.; Gold, L.I.; Opas, M. Calreticulin, a Multi-Process Calcium-Buffering Chaperone of the Endoplasmic Reticulum. Biochem. J. 2009, 417, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Berchtold, C.M.; Wu, Z.H.; Huang, T.T.; Miyamoto, S. Calcium-Dependent Regulation of NEMO Nuclear Export in Response to Genotoxic Stimuli. Mol. Cell. Biol. 2007, 27, 497–509. [Google Scholar] [CrossRef]
- Holaska, J.M.; Black, B.E.; Rastinejad, F.; Paschal, B.M. Ca2+-Dependent Nuclear Export Mediated by Calreticulin. Mol. Cell. Biol. 2002, 22, 6286–6297. [Google Scholar] [CrossRef] [PubMed]
- Defranco, D.B. Nuclear Export: DNA-Binding Domains Find a Surprising Partner. Curr. Biol. 2001, 11, R1036–R1037. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Grespin, M.E.; Bonamy, G.M.; Roggero, V.R.; Cameron, N.G.; Adam, L.E.; Atchison, A.P.; Fratto, V.M.; Allison, L.A. Thyroid Hormone Receptor Alpha1 Follows a Cooperative CRM1/Calreticulin-Mediated Nuclear Export Pathway. J. Biol. Chem. 2008, 283, 25576–25588. [Google Scholar] [CrossRef]
- Jain, P.; Mostoller, K.; Flaig, K.E.; Ahuja, J.; Lepoutre, V.; Alefantis, T.; Khan, Z.K.; Wigdahl, B. Identification of Human T Cell Leukemia Virus Type 1 Tax Amino Acid Signals and Cellular Factors Involved in Secretion of the Viral Oncoprotein. J. Biol. Chem. 2007, 282, 34581–34593. [Google Scholar] [CrossRef]
- Shembade, N.; Harhaj, N.S.; Parvatiyar, K.; Copeland, N.G.; Jenkins, N.A.; Matesic, L.E.; Harhaj, E.W. The E3 Ligase Itch Negatively Regulates Inflammatory Signaling Pathways by Controlling the Function of the Ubiquitin-Editing Enzyme A20. Nat. Immunol. 2008, 9, 254–262. [Google Scholar] [CrossRef]
- Shembade, N.; Ma, A.; Harhaj, E.W. Inhibition of NF-KappaB Signaling by A20 Through Disruption of Ubiquitin Enzyme Complexes. Science 2010, 327, 1135–1139. [Google Scholar] [CrossRef]
- Huang, T.T.; Wuerzberger-Davis, S.M.; Wu, Z.H.; Miyamoto, S. Sequential Modification of NEMO/IKKgamma by SUMO-1 and Ubiquitin Mediates NF-KappaB Activation by Genotoxic Stress. Cell 2003, 115, 565–576. [Google Scholar] [CrossRef]
- Jin, H.S.; Lee, D.H.; Kim, D.H.; Chung, J.H.; Lee, S.J.; Lee, T.H. CIAP1, CIAP2, and XIAP Act Cooperatively Via Nonredundant Pathways to Regulate Genotoxic Stress-Induced Nuclear Factor-KappaB Activation. Cancer Res. 2009, 69, 1782–1791. [Google Scholar] [CrossRef]
- Wu, Z.H.; Wong, E.T.; Shi, Y.; Niu, J.; Chen, Z.; Miyamoto, S.; Tergaonkar, V. ATM- and NEMO-Dependent ELKS Ubiquitination Coordinates TAK1-Mediated IKK Activation in Response to Genotoxic Stress. Mol. Cell 2010, 40, 75–86. [Google Scholar] [CrossRef]
- Hinz, M.; Stilmann, M.; Arslan, S.C.; Khanna, K.K.; Dittmar, G.; Scheidereit, C. A Cytoplasmic ATM-TRAF6-CIAP1 Module Links Nuclear DNA Damage Signaling to Ubiquitin-Mediated NF-KappaB Activation. Mol. Cell 2010, 40, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Hadian, K.; Krappmann, D. Signals From the Nucleus: Activation of NF-{Kappa}B by Cytosolic ATM in the DNA Damage Response. Sci. Signal. 2011, 4, e2. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S. Nuclear Initiated NF-KappaB Signaling: NEMO and ATM Take Center Stage. Cell Res. 2011, 21, 116–130. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function/comment | Methods* | References | |
| Components detected in the Tax NBs | |||
| Transcription | |||
| Tax | HTLV-1 oncoprotein | LM/EM | [33,34] |
| Pol IIO | Hyperphosphorylated form of RNA polymerase II | LM | [34,70] |
| RelA | NF-κB subunit | LM/coIP | [32,34,65,81,83] |
| p50 | NF-κB subunit | LM/coIP | [34,84] |
| IKKγ/NEMO | Regulatory subunit of IκB kinase/NF-κB Essential | LM/coIP | [23,27,32 |
| modulator | 43,81,82,85,86,87] | ||
| ATF1 | Activating Transcription Factor 1, member of CREB/ATF | LM | [13] |
| transcription factor family | |||
| CBP and p300 | Homologous transcriptional coactivators, acetyltransferases | LM/coIP | [10,13,65,88,89] |
| SMRT | Silencing mediator for retinoic acid and thyroid hormone | LM | [74] |
| receptors | |||
| Splicing | |||
| Sm | Core components of snRNP | LM/EM | [34,90] |
| SC-35 | Member of SR protein family | LM/coIP | [33,34] |
| DNA damage | |||
| response | |||
| DNA-PKcs | DNA-dependent protein kinase catalytic subunit | LM | [91] |
| BRCA1 | Breast cancer 2 | LM | [91] |
| MDC1 | Mediator of DNA damage checkpoint protein 1 | LM | [91] |
| ChK2 | Checkpoint kinase 2 | LM | [92] |
| 53BP1 | p53 binding protein 1 | LM | [93] |
| Protein modification | |||
| CBP and p300 | Homologous transcriptional coactivators, acetyltransferases | LM/coIP | [10,13,65,88,89], |
| Figure 6 | |||
| Ubc9 | SUMO E2 conjugating enzyme | LM | [82] |
| ChK2 | Checkpoint kinase 2 | LM | [93] |
| CDK8 | Cyclin-dependent kinase 8 | LM | [34] |
| SUMO-1 | Small ubiquitin-like modifier | LM | [32,51] |
| Nucleocytoplasmic | |||
| transport | |||
| CRM-1 | Chromosome region maintenance protein 1, nuclear export | LM | [70] |
| Calreticulin | Calcium-buffering chaperone, nuclear export of hormone | LM | [81] |
| receptors | |||
| Nucleotides | |||
| Nascent RNA | Nascent ribonucleic acid | LM | [70] |
| Components not detected in Tax NBs | |||
| PML | Promyelocytic leukemia protein: component of PML NBs | LM | [32,34,74], |
| Figure 3A, Figure 4, Figure 6 | |||
| Coilin | Component of Cajal bodies | LM | [34,90] |
| Nbs1 | Component of the repair foci | LM | [91] |
| Ubiquitin (Ub) | 76 amino acid peptide conjugated to lysines of target proteins, | LM | [32] |
| either as mono-Ub or via branching at lysine K48 or K63 of Ub | |||
| p53 | Tumor suppressor p53 | LM | Figure 6 |
| DNA | Desoxyribonucleic acid | LM/EM | [70,90] |
| Rex | HTLV-1 post-transcriptional regulator | LM | [70] |
| Function/comment | Methods* | References | |
| Golgi-associated lipid | |||
| raft microdomains | |||
| Tax | HTLV-1 oncoprotein | LM | [32,54,120,121] |
| IKKγ/ΝΕΜO | Regulatory subunit of IκB kinase/NF-κB Essential Modulator | LM/LRF | [32,54,121] |
| IKKα | Catalytic subunit α of IKK | LM/LRF | [54,121] |
| IKKβ | Catalytic subunit β of IKK | LM/LRF | [54,121] |
| Hsp90 | Heat shock protein 90, IKK-associated chaperone protein | LRF | [54] |
| TAK1 | TGFβ activating kinase, IKK-upstream kinase | LRF | [54] |
| GM130 | Golgi matrix protein 130, marker of the Golgi apparatus | LM | [54,120,121] |
| Giantin | Marker of the Golgi apparatus | LM | [120] |
| GM1, LAT, Calveolin | Markers of lipid rafts | LM/LRF | [54] |
| Ubiquitin (Ub) | 76 amino acid peptide conjugated to lysines of target proteins, | LM | [32] |
| either as mono-Ub or via branching at lysines K48 or K63 | |||
| of Ub | |||
| TAB2-containing | |||
| cytoplasmic foci | |||
| Tax | HTLV-1 oncoprotein | LM | [81] |
| RelA | NF-κB subunit | LM/coIP | [81] |
| Calreticulin | Calcium-buffering chaperone, nuclear export of hormone | LM/coIP | [81,122] |
| receptors | |||
| IKKγ/ΝΕΜO | Regulatory subunit of IκB kinase/NF-κB Essential Modulator | LM/coIP | [32,81] |
| IKKα | Catalytic subunit α of IKK | LM | [32] |
| IKKβ | Catalytic subunit β of IKK | LM | [32] |
| TAB2 | TGFβ activating kinase binding protein 2 | LM | [81] |
| TAX1-BP1 | TAX1 binding protein 1, component of the | LM | [81] |
| A20/Itch/TAX1-BP1 ubiquitin editing complexes | |||
| Centrosome | |||
| Tax | HTLV-1 oncoprotein | LM/coIP | [59] |
| γ-Tubulin | Marker of the centrosome | LM/coIP | [59] |
| IKKγ/ΝΕΜO | Regulatory subunit of IκB kinase/NF-κB Essential Modulator | LM/coIP | [59] |
| IKKα | Catalytic subunit α of IKK | coIP | [59] |
| IKKβ | Catalytic subunit β of IKK | coIP | [59] |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lodewick, J.; Lamsoul, I.; Bex, F. Move or Die: the Fate of the Tax Oncoprotein of HTLV-1. Viruses 2011, 3, 829-857. https://doi.org/10.3390/v3060829
Lodewick J, Lamsoul I, Bex F. Move or Die: the Fate of the Tax Oncoprotein of HTLV-1. Viruses. 2011; 3(6):829-857. https://doi.org/10.3390/v3060829
Chicago/Turabian StyleLodewick, Julie, Isabelle Lamsoul, and Françoise Bex. 2011. "Move or Die: the Fate of the Tax Oncoprotein of HTLV-1" Viruses 3, no. 6: 829-857. https://doi.org/10.3390/v3060829
APA StyleLodewick, J., Lamsoul, I., & Bex, F. (2011). Move or Die: the Fate of the Tax Oncoprotein of HTLV-1. Viruses, 3(6), 829-857. https://doi.org/10.3390/v3060829