

4-Hydroxychalcone Inhibits Human Coronavirus HCoV-OC43 by Targeting EGFR/AKT/ERK1/2 Signaling Pathway

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals, Cells, and Viruses
2.2. Cytotoxicity and Cytopathic Effect (CPE) Inhibition Assays
2.3. Mouse Model of HCoV-OC43 Infection
2.4. Time of Addition and Plaque Reduction Assay
2.5. RNA Isolation and Quantitative Reverse Transcription PCR (RT-qPCR)
2.6. Western Blot Analysis
2.7. Transcriptome and Connectivity Map (cMap) Analysis
2.8. Cellular Thermal Shift Assay (CETSA)
2.9. Molecular Docking
2.10. Statistical Analysis
3. Results
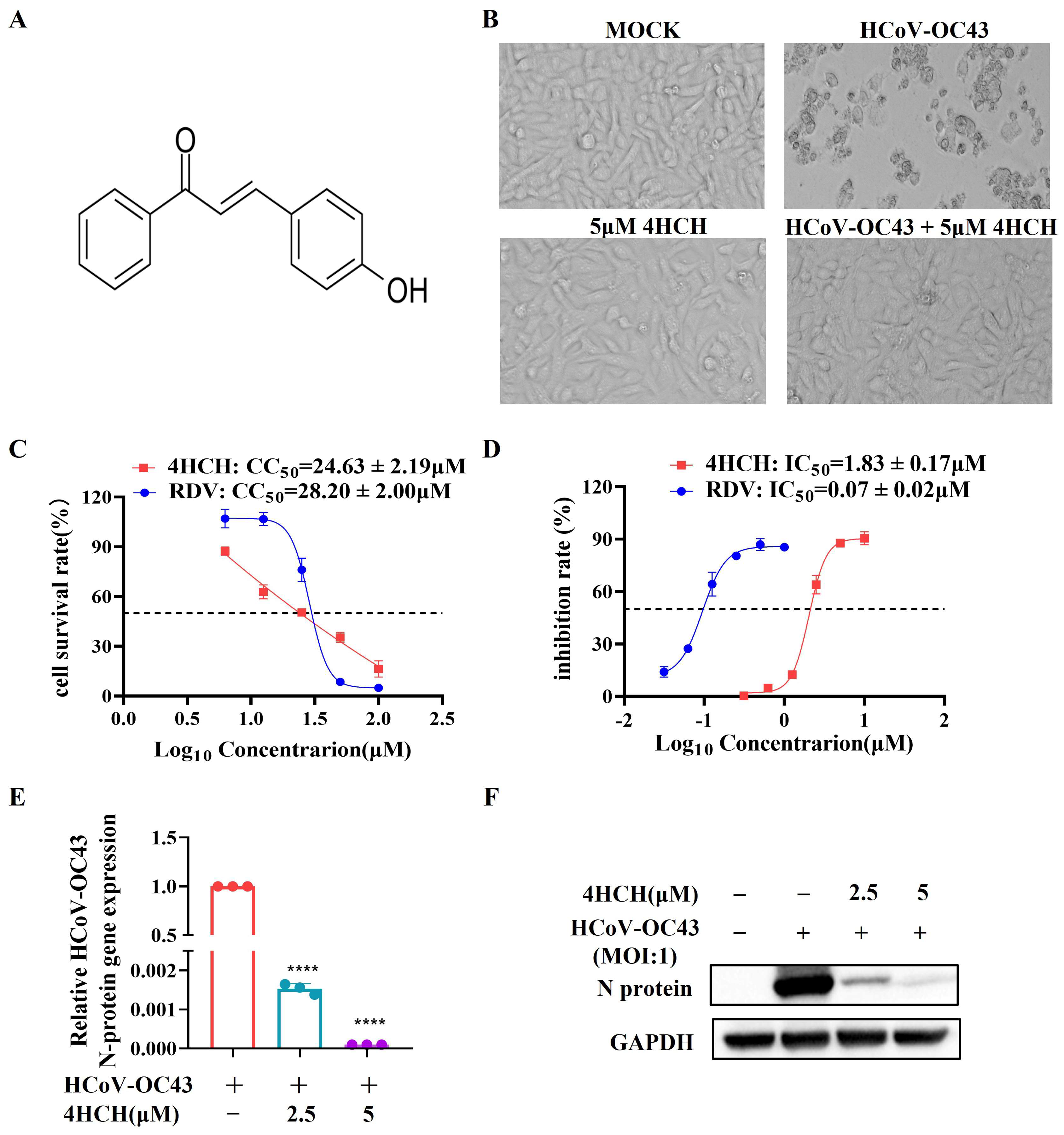
3.1. 4HCH Exhibits Potent Antiviral Activity Against HCoV-OC43 In Vitro
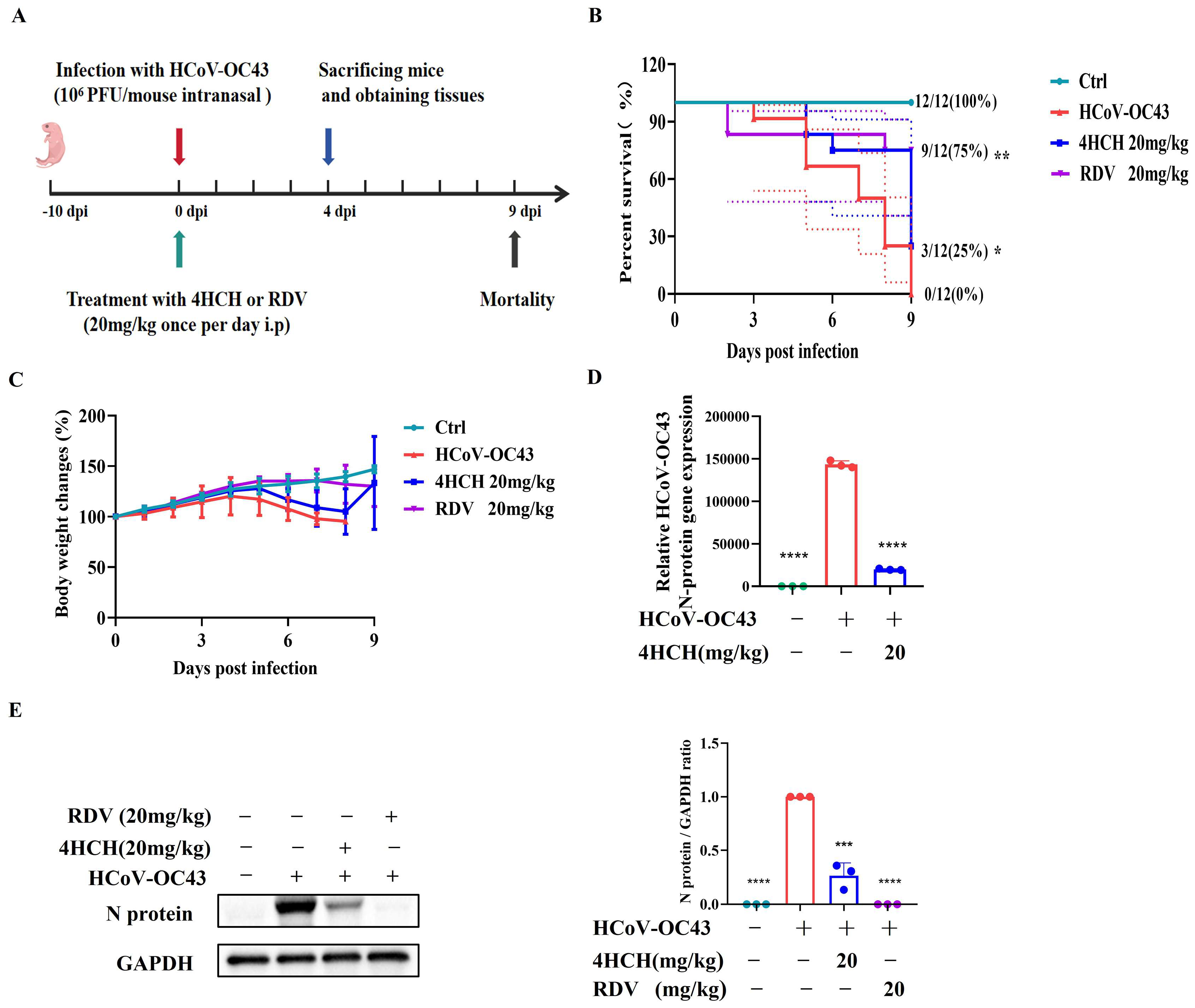
3.2. 4HCH Demonstrates Antiviral Efficacy in a Suckling Mouse Model
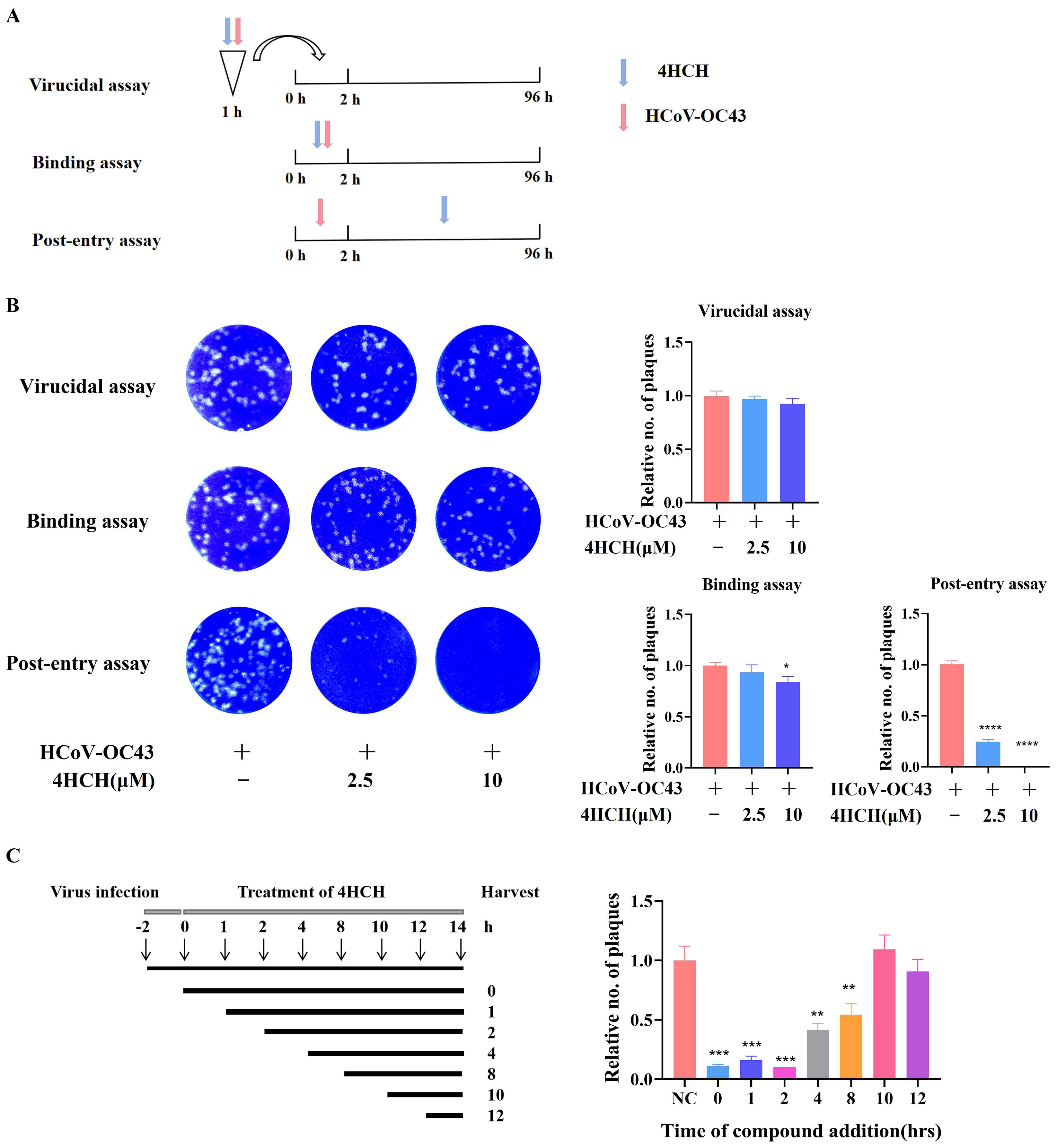
3.3. 4HCH Targets the Post-Entry Phase of HCoV-OC43 Infection
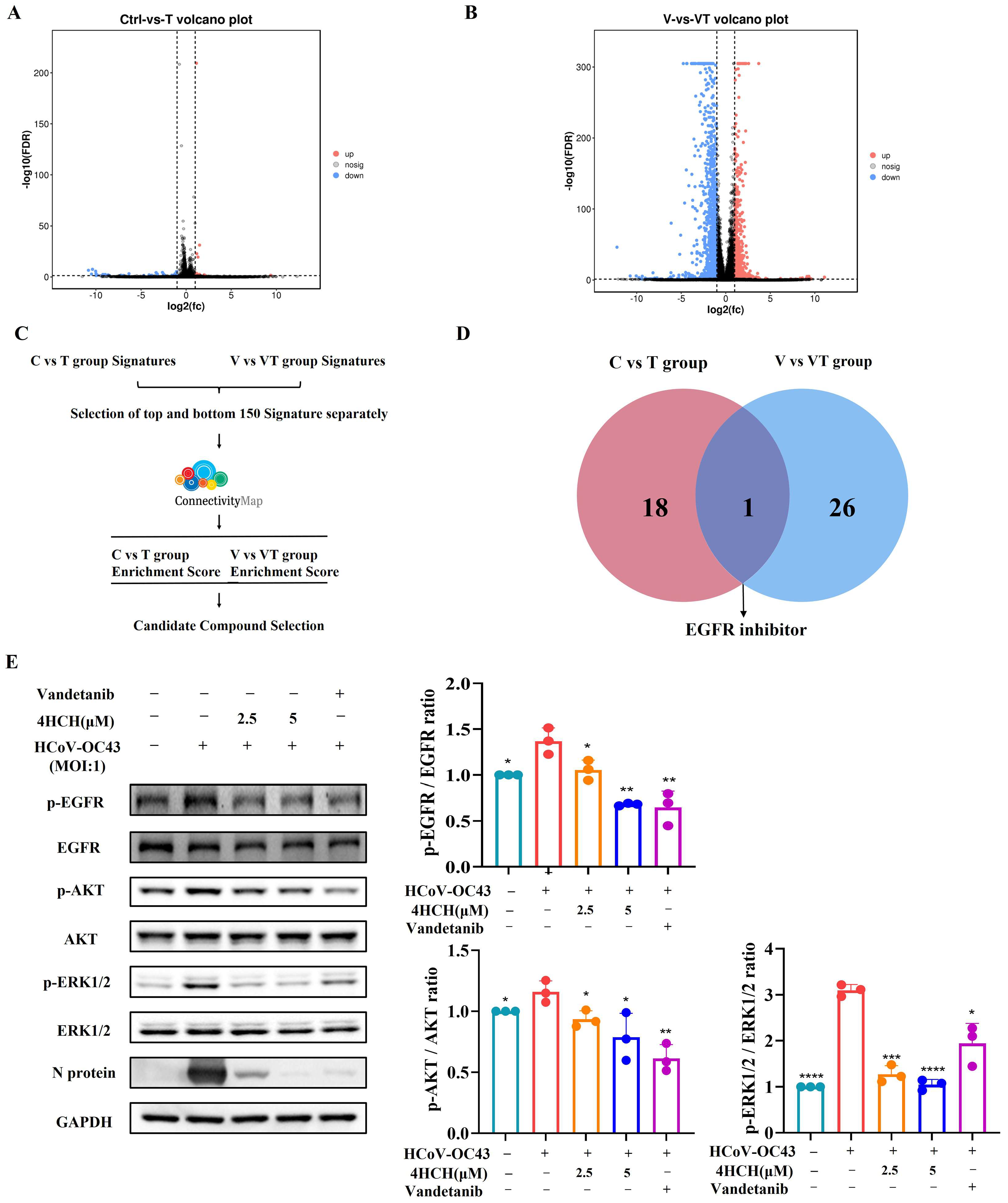
3.4. 4HCH Inhibits the EGFR/AKT/ERK1/2 Signaling Pathway
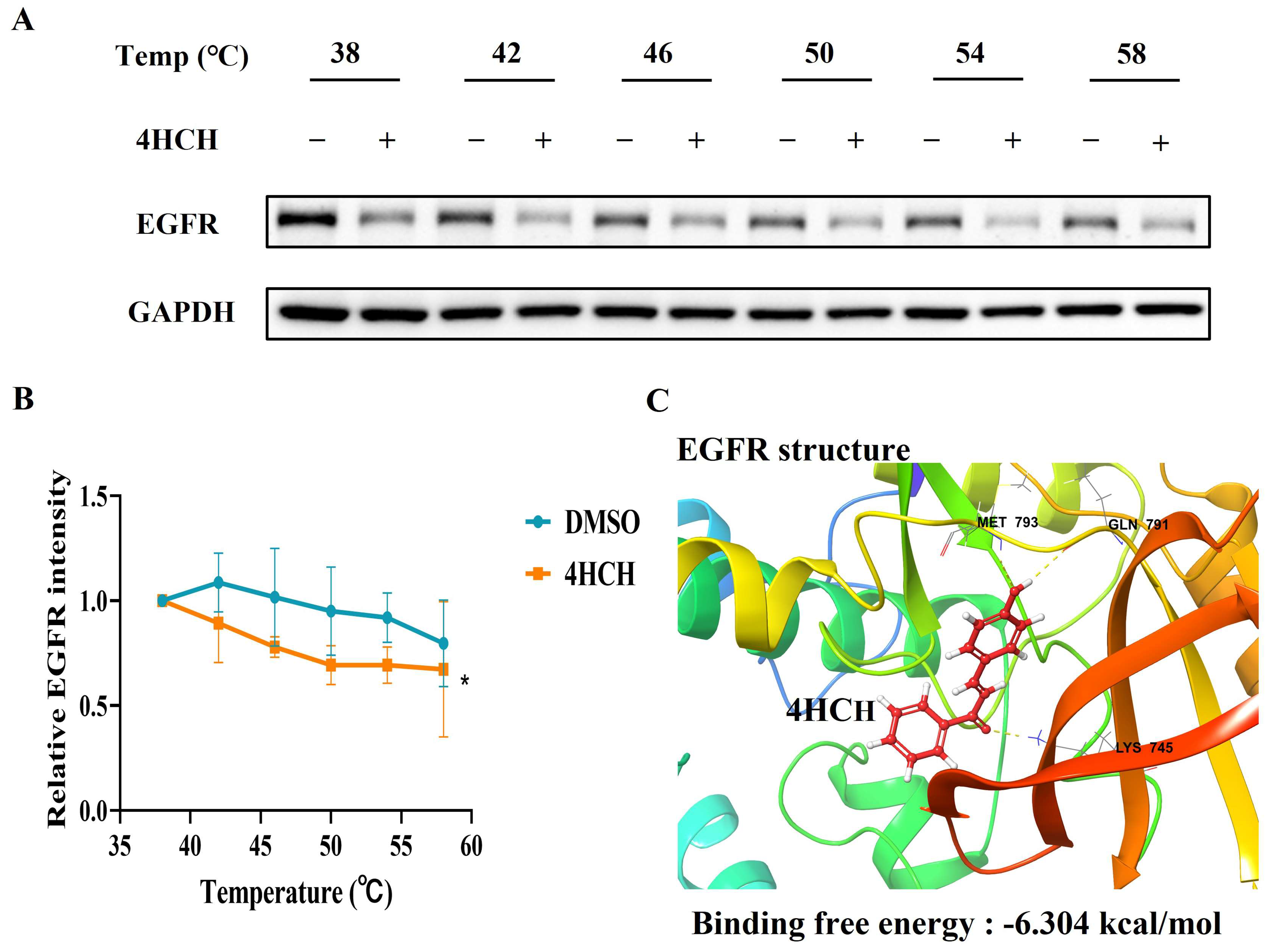
3.5. 4HCH Likely Interacts with EGFR
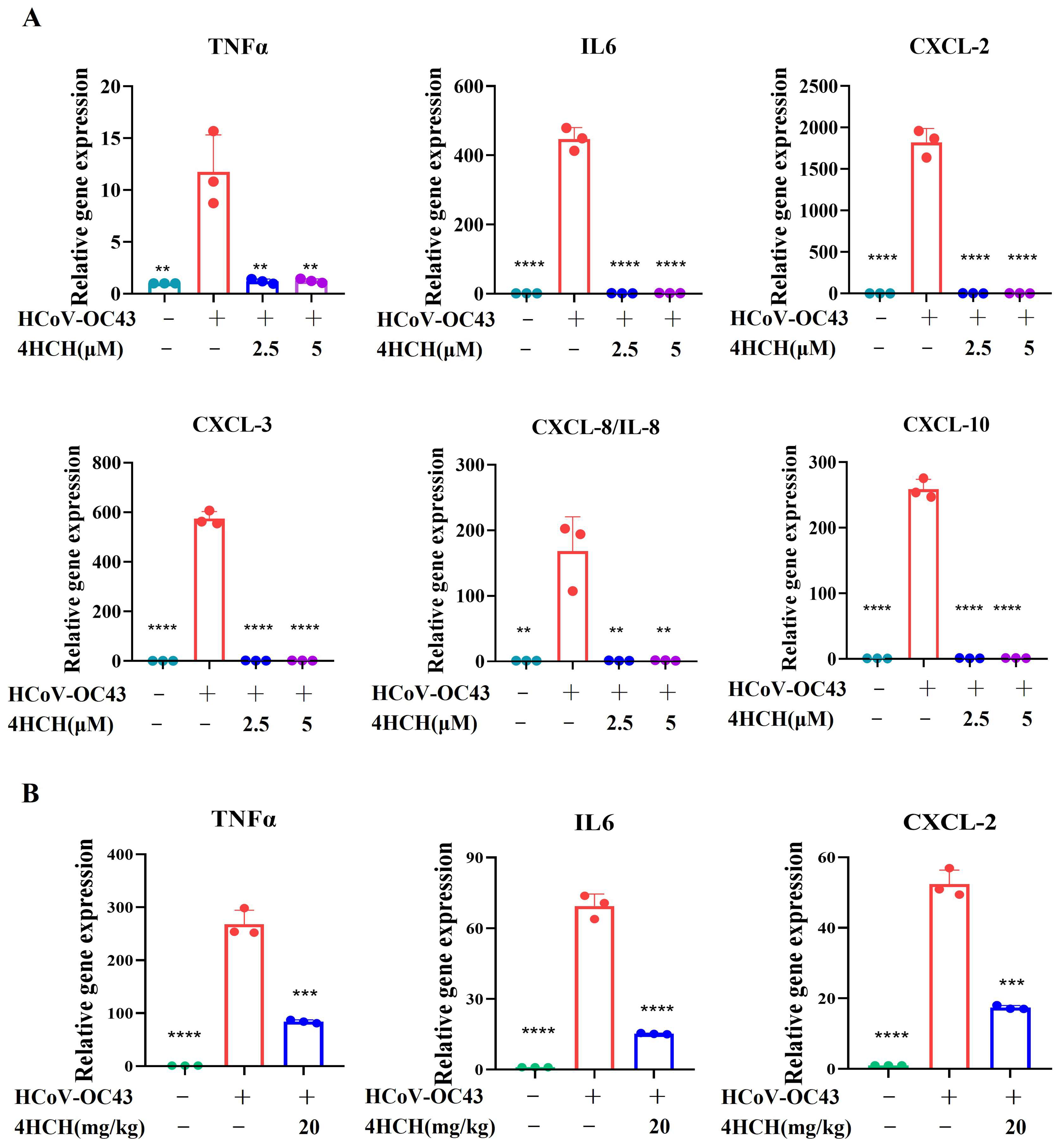
3.6. 4HCH Suppresses Cytokine and Chemokine Expression Induced by HCoV-OC43 Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4-HCH | 4-hydroxychalcone |
| AKT | protein kinase B |
| ARDS | acute respiratory distress syndrome |
| CC50 | 50% cytotoxic concentration |
| CETSA | cellular thermal shift assay |
| cMAP | connectivity map |
| CPE | cytopathic effect |
| COVID-19 | coronavirus disease 2019 |
| EGFR | epidermal growth factor receptor |
| ERK1/2 | extracellular regulated protein kinases 1/2 |
| HCoV-OC43 | human coronavirus OC43 |
| HCoVs | human coronaviruses |
| IC50 | 50% inhibitory concentration |
| MAPK | mitogen-activated protein kinase |
| MERS-CoV | Middle East Respiratory Syndrome coronavirus |
| MHV-A59 | mouse hepatitis virus A-59 |
| MOI | multiplicity of infection |
| PFU | plaque forming units |
| RD | rhabdomyosarcoma |
| RDV | Remdesivir |
| RT-qPCR | quantitative real-time PCR |
| SARS-CoV | severe acute respiratory syndrome coronavirus |
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus 2 |
References
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Liu, Z.; Chen, D. Human coronaviruses: Origin, host and receptor. J. Clin. Virol. 2022, 155, 105246. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.S.; Liu, D.X. Human Coronavirus: Host-Pathogen Interaction. Annu. Rev. Microbiol. 2019, 73, 529–557. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Horby, P.W.; Hayden, F.G.; Gao, G.F. A novel coronavirus outbreak of global health concern. Lancet 2020, 395, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Gaunt, E.R.; Hardie, A.; Claas, E.C.J.; Simmonds, P.; Templeton, K.E. Epidemiology and clinical presentations of the four human coronaviruses 229E, HKU1, NL63, and OC43 detected over 3 years using a novel multi plex real-time PCR method. J. Clin. Microbiol. 2010, 48, 2940–2947. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.; Edner, N.; Albert, J.; Ternhag, A. Fatal encephalitis associated with coronavirus OC43 in an immunocompromised child. Infect. Dis. 2020, 52, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Parums, D.V. Editorial: The XBB.1.5 (‘Kraken’) Subvariant of Omicron SARS-CoV-2 and its Rapid Global Spread. Med. Sci. Monit. 2023, 29, e939580. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Long, Y.; Wang, F.; Li, C.; Liu, W. Characterization of SARS-CoV-2 recombinants and emerging Omicron sublineages. Int. J. Med. Sci. 2023, 20, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Ao, D.; He, X.; Liu, J.; Xu, L. Strategies for the development and approval of COVID-19 vaccines and therapeutics in the post-pandemic period. Signal Transduct. Target. Ther. 2020, 8, 466. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.I.; Lee, C. Human Coronavirus OC43 as a Low-Risk Model to Study COVID-19. Viruses 2023, 15, 578. [Google Scholar] [CrossRef] [PubMed]
- Sopjani, M.; Falco, F.; Impellitteri, F.; Guarrasi, V.; Nguyen Thi, X.; Dërmaku-Sopjani, M.; Faggio, C. Flavonoids derived from medicinal plants as a COVID-19 treatment. Phytother. Res. 2004, 38, 1589–1609. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-Y.; Ma, Y.-X.; Liu, Y.; Peng, X.-J.; Chen, X.-Z. A Comprehensive Review of Natural Flavonoids with Anti-SARS-CoV-2 Activity. Molecules 2024, 28, 2735. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Vargas, J.; Shapira, T.; Olmstead, A.D.; Villanueva, I.; Thompson, C.A.H.; Ennis, S.; Gao, G.; De Guzman, J.; Williams, D.E.; Wang, M.; et al. Discovery of lead natural products for developing pan-SARS-CoV-2 therapeutics. Antivir. Res. 2023, 209, 105484. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, S.A.M.; Yosri, N.; El-Mallah, M.F.; Ghonaim, R.; Guo, Z.; Musharraf, S.G.; Du, M.; Khatib, A.; Xiao, J.; Saeed, A.; et al. Screening for natural and derived bio-active compounds in preclinical and clinical studies: One of the frontlines of fighting the coronaviruses pandemic. Phytomedicine 2021, 85, 153311. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z. Chalcone: A Privileged Structure in Medicinal Chemistry. Chem. Rev. 2021, 117, 7762–7810. [Google Scholar] [CrossRef] [PubMed]
- Saha, C.; Naskar, R.; Chakraborty, S. Antiviral Flavonoids: A Natural Scaffold with Prospects as Phytomedicines against SARS-CoV2. Mini Rev. Med. Chem. 2017, 24, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Mezgebe, K.; Melaku, Y.; Mulugeta, E. Synthesis and Pharmacological Activities of Chalcone and Its Derivatives Bearing N-Heterocyclic Scaffolds: A Review. ACS Omega 2024, 8, 19194–19211. [Google Scholar] [CrossRef] [PubMed]
- Elkhalifa, D.; Al-Hashimi, I.; Al Moustafa, A.-E.; Khalil, A. A comprehensive review on the antiviral activities of chalcones. J. Drug Target. 2022, 29, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, H.; Wang, M.; Sun, D.; Lu, J.; Zhu, T.; Chen, H.; Chen, L.; Liu, S. Discovery of Chalcone Derivatives as Bifunctional Molecules with Anti- SARS-CoV-2 and Anti-inflammatory Activities. J. Nat. Prod. 2021, 87, 2680–2694. [Google Scholar] [CrossRef] [PubMed]
- Herzog, A.-M.; Göbel, K.; Marongiu, L.; Ruetalo, N.; Alonso, M.C.; Leischner, C.; Busch, C.; Burkard, M.; Lauer, U.M.; Geurink, P.P.; et al. Compounds derived from Humulus lupulus inhibit SARS-CoV-2 papain-like protease and virus replication. Phytomedicine 2024, 123, 155176. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.-H.; Min, J.S.; Kwon, S. Cardamonin as a p38 MAPK Signaling Pathway Activator Inhibits Human Coronavirus OC43 Infection in Human Lung Cells. Nutrients 2024, 15, 1335. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, M.; Qin, H.; Lin, H.; An, X.; Shi, Z.; Song, L.; Yang, X.; Fan, H.; Tong, Y. Artemether, Artesunate, Arteannuin B, Echinatin, Licochalcone B and An drographolide Effectively Inhibit SARS-CoV-2 and Related Viruses In Vitro. Front. Cell. Infect. Microbiol. 2023, 11, 680127. [Google Scholar]
- Shen, L.; Niu, J.; Wang, C.; Huang, B.; Wang, W.; Zhu, N.; Deng, Y.; Wang, H.; Ye, F.; Cen, S.; et al. High-Throughput Screening and Identification of Potent Broad-Spectrum Inhibitors of Coronaviruses. J. Virol. 2021, 93, e00023-19. [Google Scholar] [CrossRef] [PubMed]
- Varinska, L.; van Wijhe, M.; Belleri, M.; Mitola, S.; Perjesi, P.; Presta, M.; Koolwijk, P.; Ivanova, L.; Mojzis, J. Anti-angiogenic activity of the flavonoid precursor 4-hydroxychalcone. Eur. J. Pharmacol. 2019, 691, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Qu, Q.; Dai, B.; Yang, B.; Li, X.; Liu, Y.; Zhang, F. 4-Hydroxychalcone attenuates hyperaldosteronism, inflammation, and ren al injury in cryptochrome-null mice. BioMed Res. Int. 2012, 2014, 603415. [Google Scholar]
- Han, X.; Zhu, Q.-Q.; Li, Z.; He, J.-K.; Sun, Y.; Zhong, Q.-H.; Tang, S.-X.; Zhang, Y.-L. 4-Hydroxychalcone attenuates AngII-induced cardiac remodeling and dysfunction via regulating PI3K/AKT pathway. Hypertens. Res. 2014, 48, 1054–1067. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.-C.; Wang, Y.; Liu, Y.-P.; Zhang, R.-Q.; Li, X.; Su, W.-H.; Long, F.; Luo, X.-D.; Peng, T. Inhibition of enterovirus 71 replication by chrysosplenetin and penduletin. Eur. J. Pharm. Sci. 2011, 44, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Narayan, R.; Corsello, S.M.; Peck, D.D.; Natoli, T.E.; Lu, X.; Gould, J.; Davis, J.F.; Tubelli, A.A.; Asiedu, J.K.; et al. A Next Generation Connectivity Map: L1000 Platform and the First 1,000,000 Profiles. Cell 2017, 171, 1437–1452.e17. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Lee, W.; Ku, K.B.; Yoon, G.Y.; Moon, H.W.; Kim, C.; Kim, M.H.; Yi, Y.S.; Jun, S.; Kim, B.T.; et al. SARS-CoV-2 aberrantly elevates mitochondrial bioenergetics to induce robust virus propagation. Signal Transduct. Target. Ther. 2024, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, X.; Peng, C.; Tan, C.; Sun, H.; Liu, H.; Zhang, Y.; Wu, P.; Cui, C.; Liu, C.; et al. The phytochemical hyperforin triggers thermogenesis in adipose tissue via a Dlat-AMPK signaling axis to curb obesity. Cell Metab. 2021, 33, 565–580.e7. [Google Scholar] [CrossRef] [PubMed]
- Ueki, I.F.; Min-Oo, G.; Kalinowski, A.; Ballon-Landa, E.; Lanier, L.L.; Nadel, J.A.; Koff, J.L. Respiratory virus-induced EGFR activation suppresses IRF1-dependent interferon λ and antiviral defense in airway epithelium. J. Exp. Med. 2013, 210, 1929–1936. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Niu, S.; Wang, S.; Shi, X.; Feng, S.; Du, L.; Zhang, X.; Ma, Z.; Yu, B.; Liu, H. Discovery of WS-157 as a highly potent, selective and orally active EG. Acta Pharm. Sin. B 2019, 9, 1193–1203. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Jing, J.; Du, L.; Zhang, X.; Ren, Y.; Yang, H.; Yu, B.; Liu, H. Discovery of YS-363 as a highly potent, selective, and orally efficacious EGFR inhibitor. Biomed. Pharmacother. 2023, 167, 115491. [Google Scholar] [CrossRef] [PubMed]
- Jafari, R.; Almqvist, H.; Axelsson, H.; Ignatushchenko, M.; Lundbäck, T.; Nordlund, P.; Martinez Molina, D. The cellular thermal shift assay for evaluating drug target interactions in cells. Nat. Protoc. 2014, 9, 2100–2122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ye, C.; Liu, J.; Xu, W.; Wu, C.; Yu, Q.; Xu, X.; Zeng, X.; Jin, H.; Wu, Y.; et al. JaponiconeA induces apoptosis of bortezomib-sensitive and -resistant myeloma cells in vitro and in vivo by targeting IKK. Cancer Biol. Med. 2021, 19, 651–668. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Li, R.; Pan, W.; Huang, W.; Liu, B.; Xie, Y.; Wang, Z.; Li, C.; Jiang, H.; Huang, J.; et al. Phillyrin (KD-1) exerts anti-viral and anti-inflammatory activities against novel coronavirus (SARS-CoV-2) and human coronavirus 229E (HCoV-229E) by suppressing the nuclear factor kappa B (NF-κB) signaling path way. Phytomedicine 2020, 78, 153296. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.; Fang, Y.; Shen, Z.; Shao, Y.; Ma, Q.; Yang, Z.; Zhao, J.; Li, H.; Li, R.; Dong, S.; et al. Broad antiviral and anti-inflammatory activity of Qingwenjiere mixture against SARS-CoV-2 and other human coronavirus infections. Phytomedicine 2021, 93, 153808. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, T.; Frieman, M.B. The role of epidermal growth factor receptor (EGFR) signaling in SARS coronavirus-induced pulmonary fibrosis. Antivir. Res. 2017, 143, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Channappanavar, R.; Perlman, S. Pathogenic human coronavirus infections: Causes and consequences of cytokine storm and immunopathology. Semin. Immunopathol. 2017, 39, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.E.; Chambers, R.C. The mercurial nature of neutrophils: Still an enigma in ARDS? Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L217–L230. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′ Sequence) | Reverse (5′-3′ Sequence) |
|---|---|---|
| HCoV-OC43 N | AGCAACCAGGCTGATGTCAATACC | AGCAGACCTTCCTGAGCCTTCAAT |
| TNFα | GAACCCCGAGTGACAAGCCT | TATCTCTCAGCTCCACGCCAT |
| IL6 | GCAATAACCACCCCTGACCCAA | GCTACATTTGCCGAAGAGCC |
| CXCL-2 | CCGAAGTCATAGCCACACTCA | TGGATTTGCCATTTTTCAGCATCT |
| CXCL-3 | TGAATGTAAGGTCCCCCGGA | CACCCTGCAGGAAGTGTCAA |
| CXCL-8 | CAGTTTTGCCAAGGAGTGCT | ACTTCTCCACAACCCTCTGC |
| CXCL-10 | TGAATCCAGAATCGAAGGCCA | TGCATCGATTTTGCTCCCCT |
| β-Actin | CTCACCATGGATGATGATATCGC | ATAGGAATCCTTCTGACCCATGC |
| TNFα (Mouse) | AGGGTCTGGGCCATAGAACT | CCACCACGCTCTTCTGTCTAC |
| IL6 (Mouse) | ATCCAGTTGCCTTCTTGGGACTGA | TAAGCCTCCGACTTGTGAAGTGGT |
| CXCL-2 (Mouse) | CACTCTCAAGGGCGGTCAA | AGGCACATCAGGTACGATCCA |
| GAPDH (Mouse) | CCTCGTCCCGTAGACAAAATG | TGAGGTCAATGAAGGGGTCGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Li, J.; Luo, Q.; Dai, Y.; Luo, X.; Xu, J.; Ye, W.; Zhou, X.; Diao, J.; Ren, Z.; et al. 4-Hydroxychalcone Inhibits Human Coronavirus HCoV-OC43 by Targeting EGFR/AKT/ERK1/2 Signaling Pathway. Viruses 2025, 17, 1028. https://doi.org/10.3390/v17081028
Huang Y, Li J, Luo Q, Dai Y, Luo X, Xu J, Ye W, Zhou X, Diao J, Ren Z, et al. 4-Hydroxychalcone Inhibits Human Coronavirus HCoV-OC43 by Targeting EGFR/AKT/ERK1/2 Signaling Pathway. Viruses. 2025; 17(8):1028. https://doi.org/10.3390/v17081028
Chicago/Turabian StyleHuang, Yuanyuan, Jieyu Li, Qiting Luo, Yuexiang Dai, Xinyi Luo, Jiapeng Xu, Wei Ye, Xinrui Zhou, Jiayi Diao, Zhe Ren, and et al. 2025. "4-Hydroxychalcone Inhibits Human Coronavirus HCoV-OC43 by Targeting EGFR/AKT/ERK1/2 Signaling Pathway" Viruses 17, no. 8: 1028. https://doi.org/10.3390/v17081028
APA StyleHuang, Y., Li, J., Luo, Q., Dai, Y., Luo, X., Xu, J., Ye, W., Zhou, X., Diao, J., Ren, Z., Liu, G., He, Z., Wang, Z., Wang, Y., & Zhu, Q. (2025). 4-Hydroxychalcone Inhibits Human Coronavirus HCoV-OC43 by Targeting EGFR/AKT/ERK1/2 Signaling Pathway. Viruses, 17(8), 1028. https://doi.org/10.3390/v17081028