
Comprehensive Virome Profiling of Apple Mosaic Disease-Affected Trees in Iran Using RT-PCR and Next-Generation Sequencing

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
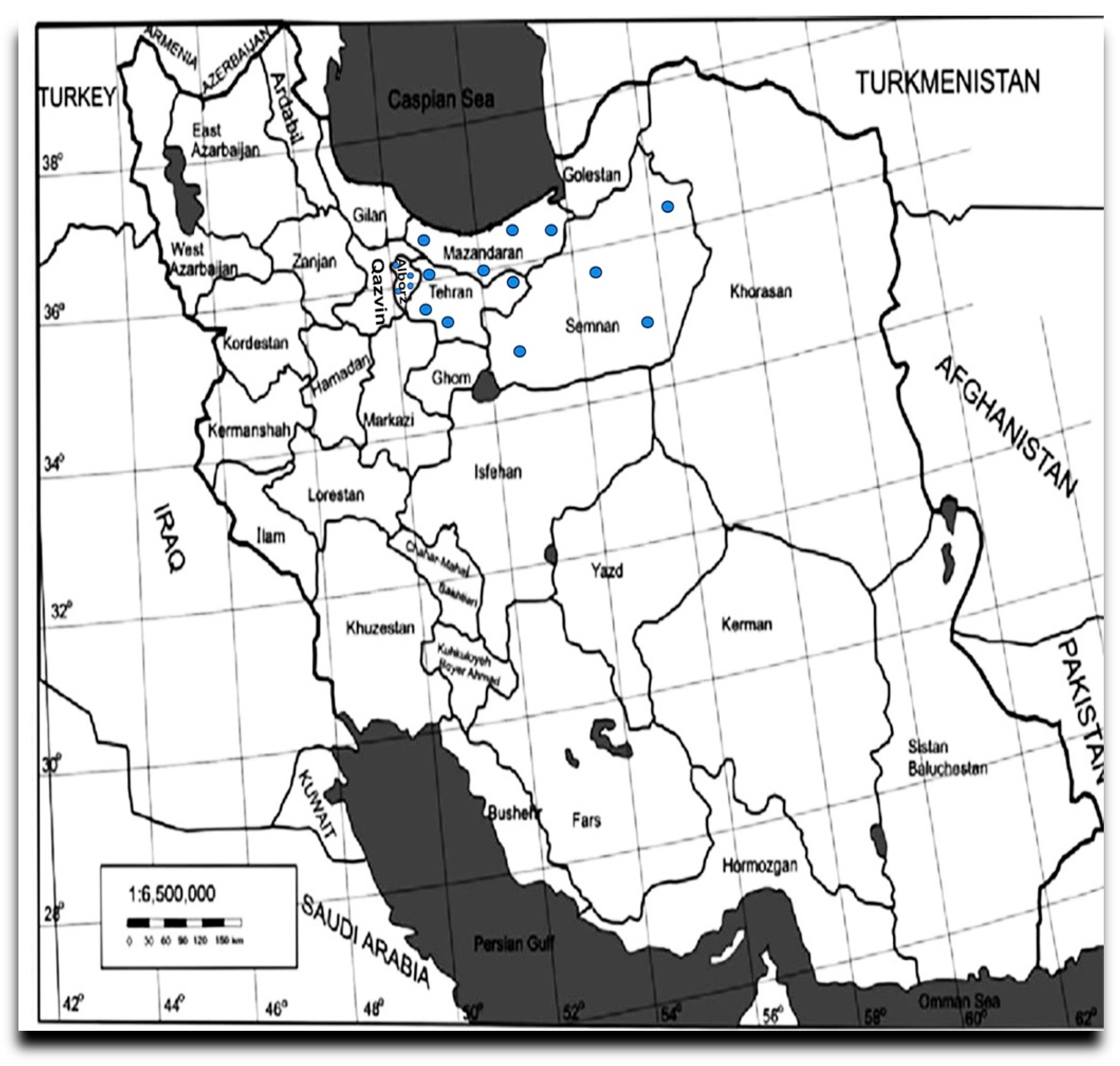
2.1. Plant Material and Sample Preparation
2.2. RT-PCR for Viruses and Viroids Detection
2.3. Next-Generation Sequencing (NGS)
2.4. Bioinformatic Analysis
2.5. Phylogenetic Analysis
3. Results
3.1. Detection and Prevalence of Apple Viruses and Viroids Using RT-PCR Assays
3.2. NGS-Identified Viruses and Viroids in Apple and Their Prevalence Validated by RT-PCR
3.3. Regional and Varietal Distribution of Virus and Viroid Infections
3.4. Full Genome Sequencing of Detected Viruses and Viroids
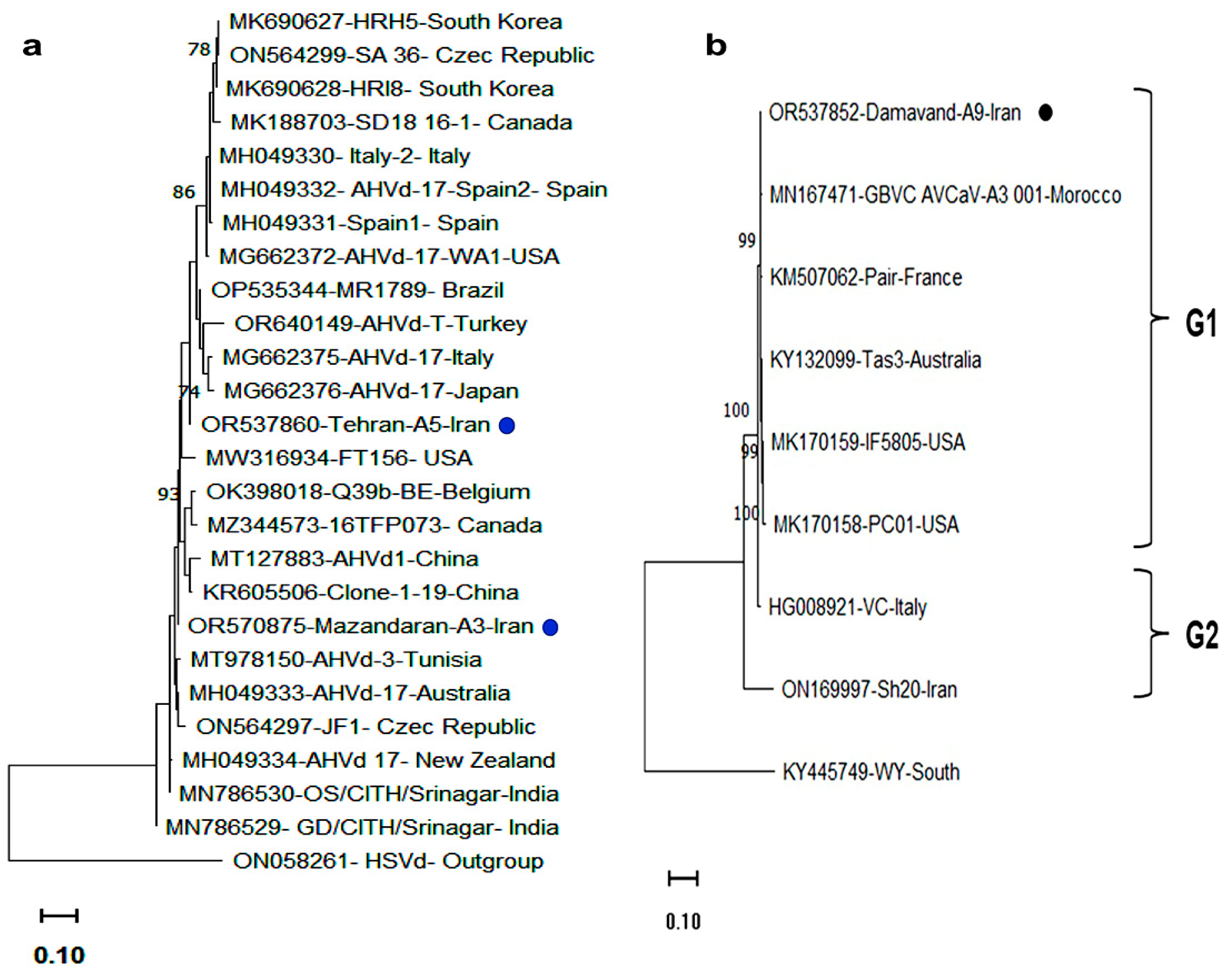
3.5. Phylogenetic and Sequence Variability Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. World Food and Agriculture Statistical Yearbook; FAO: Rome, Italy, 2020; Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 9 July 2020).
- Nabi, S.U.; Baranwal, V.K.; Yadav, M.K.; Rao, G.P. Association of apple necrotic mosaic virus (ApNMV) with mosaic disease in commercially grown cultivars of apple (Malus domestica Borkh.) in India. Biotech 2020, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.A.C. Global plant virus disease pandemics and epidemics. Plants 2021, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Dong, Y.; Zhang, Z.; Fan, X.; Ren, F. Elimination of apple necrosis mosaic virus from potted apple plants by thermotherapy combined with shoot-tip grafting. Sci. Hortic. 2019, 252, 310–315. [Google Scholar] [CrossRef]
- Umer, M.; Liu, J.; You, H.; Xu, C.; Dong, K.; Luo, N.; Kong, L.; Li, X.; Hong, N.; Wang, G.; et al. Genomic, morphological and biological traits of the viruses infecting major fruit trees. Viruses 2019, 11, 515. [Google Scholar] [CrossRef]
- Wright, A.A.; Cross, A.R.; Harper, S.J.A. Bushel of viruses: Identification of seventeen novel putative viruses by RNA-seq in six apple trees. PLoS ONE 2020, 15, e0227669. [Google Scholar] [CrossRef]
- Chen, S.; Ye, T.; Hao, L.; Chen, H.; Wang, S.; Fan, Z.; Guo, L.; Zhou, T. Infection of apple by Apple stem grooving virus leads to extensive alterations in gene expression patterns but no disease symptoms. PLoS ONE 2014, 9, e95239. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization (EPPO). Pathogen-Tested Material of Malus, Pyrus and Cydonia. OEPP/EPPO Bull. 1999, 29, 239–252. [Google Scholar] [CrossRef]
- Beaver-Kanuya, E.; Szostek, S.A.; Harper, S.J. Development of real-time RT-PCR assays for two viruses infecting pome fruit. J. Virol. Methods 2019, 266, 25–29. [Google Scholar] [CrossRef]
- Maliogka, V.I.; Minafra, A.; Saldarelli, P.; Ruiz-García, A.B.; Glasa, M.; Katis, N.; Olmos, A. Recent advances on detection and characterization of fruit tree viruses using high-throughput sequencing technologies. Viruses 2018, 10, 436. [Google Scholar] [CrossRef]
- Santala, J.; Valkonen, J.P.T. Sensitivity of small RNA-based detection of plant viruses. Front. Microbiol. 2018, 9, 939. [Google Scholar] [CrossRef]
- Roossinck, M.J. Deep sequencing for discovery and evolutionary analysis of plant viruses. Virus Res. 2017, 239, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Canales, C.; Morán, F.; Olmos, A.; Ruiz-García, A.B. First detection and molecular characterization of Apple stem grooving virus, Apple chlorotic leaf spot virus, and Apple hammerhead viroid in loquat in Spain. Plants 2021, 10, 2293. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Hao, W.; Storoschuk, G.; MacDonald, J.; Sanfaçon, H. Characterizing the virome of apple orchards affected by rapid decline in the Okanagan and Similkameen valleys of British Columbia (Canada). Pathogens 2022, 11, 1231. [Google Scholar] [CrossRef] [PubMed]
- Várallyay, E.; Přibylová, J.; Galbacs, Z.N.; Jahan, A.; Varga, T.; Špak, J.; Lenz, O.; Fránová, J.; Sedlák, J.; Koloniuk, I. Detection of apple hammerhead viroid, apple luteovirus 1 and citrus concave gum-associated virus in apple propagation materials and orchards in the Czech Republic and Hungary. Viruses 2022, 14, 2347. [Google Scholar] [CrossRef]
- Moini, A.A. Identification of Tomato ringspot virus (ToRSV) on apple in Iran. Aust. Plant Dis. Notes 2010, 5, 105. [Google Scholar] [CrossRef]
- Abtahi, F.; Shams-Bakhsh, M.; Safaie, N.; Autonell, C.R.; Ratti, C. Occurrence, distribution, and molecular characterization of apple stem pitting virus in Iran. J. Agric. Sci. Technol. 2017, 19, 217–230. [Google Scholar]
- Abtahi, F.; Shams-Bakhsh, M.; Safaie, N.; Azizi, A.; Autonell, C.R.; Ratti, C. Incidence and genetic diversity of apple chlorotic leaf spot virus in Iran. J. Plant Pathol. 2019, 101, 513–519. [Google Scholar] [CrossRef]
- Keshavarz, T.; Hajnajari, H. Evaluation of apple cultivars to four pome fruit viruses in Iranian national collection of Kamalshahr horticulture research station, Karaj. Arch. Phytopathol. Plant Prot. 2021, 54, 918–932. [Google Scholar] [CrossRef]
- Amirnia, F.; Hajizadeh, M. First report of apple hammerhead viroid infecting apple in Iran. J. Plant. Pathol. 2025, 107, 791–792. [Google Scholar] [CrossRef]
- Shokri, S.; Shujaei, K.; Gibbs, A.J.; Hajizadeh, M. Evolution and biogeography of apple stem grooving virus. Virol. J. 2023, 20, 105. [Google Scholar] [CrossRef]
- Foissac, X.; Svanella-Dumas, L.; Dulucq, M.J.; Candresse, T.; Gentit, P. Polyvalent detection of fruit tree Ribo-, Capillo- and Foveavirus by nested RT-PCR using degenerated and inosine containing primers (DOPRT-PCR). Acta Hortic. 2001, 550, 37–44. [Google Scholar] [CrossRef]
- Daccord, N.; Celton, J.M.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 2017, 49, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Roumi, V.; Gazel, M.; Caglayan, K. Occurrence of hop stunt viroid in apple orchards of the northwest Iran. J. Crop Prot. 2020, 9, 301–306. [Google Scholar]
- Elbeaino, T.; Giampetruzzi, A.; De Stradis, A.; Digiaro, M. Deep-sequencing analysis of an apricot tree with vein clearing symptoms reveals the presence of a novel Betaflexivirus. Virus Res. 2014, 181, 1–5. [Google Scholar] [CrossRef]
- Martelli, G.P.; Adams, M.J.; Kreuze, J.F.; Dolja, V.V. Family flexiviridae: A case study in virion and genome plasticity. Annu. Rev. Phytopathol. 2007, 45, 73–100. [Google Scholar] [CrossRef]
- Marais, A.; Faure, C.; Mustafayev, E.; Candresse, T. Characterization of New Isolates of Apricot vein clearing-associated virus and of a New Prunus-Infecting Virus: Evidence for Recombination as a Driving Force in Betaflexiviridae Evolution. PLoS ONE 2015, 10, e0129469. [Google Scholar] [CrossRef] [PubMed]
- Longdon, B.; Brockhurst, M.A.; Russell, C.A.; Welch, J.J.; Jiggins, F.M. The evolution and genetics of virus host shifts. PLOS Pathogens. 2014, 10, e1004395. [Google Scholar] [CrossRef]
- Minutolo, M.; Cinque, M.; Chiumenti, M.; Di Serio, F.; Alioto, D.; Navarro, B. Identification and characterization of Citrus concave gum-associated virus infecting citrus and apple trees by serological, molecular and high-throughput sequencing approaches. Plants 2021, 10, 2390. [Google Scholar] [CrossRef]
- Paudel, D.B.; Sanfaçon, H. Exploring the diversity of mechanisms associated with plant tolerance to virus infection. Front. Plant Sci. 2018, 9, 1575. [Google Scholar] [CrossRef]
- Wunsch, A.; Hoff, B.; Sazo, M.M.; van Zoeren, J.; Lamour, K.H.; Hurtado-Gonzales, O.P.; Fuchs, M. Viruses of Apple Are Seedborne but Likely Not Vertically Transmitted. Viruses 2024, 16, 95. [Google Scholar] [CrossRef] [PubMed]
- Malandraki, I.; Beris, D.; Isaioglou, I.; Olmos, A.; Varveri, C.; Vassilakos, N. Simultaneous detection of three pome fruit tree viruses by one-step multiplex quantitative RT-PCR. PLoS ONE 2017, 12, e0180877. [Google Scholar] [CrossRef] [PubMed]
- Vamenani, R.; Rahimian, H.; Alavi, S.; Borhani, B. A viroid resembling hop stunt viroid in infected apple trees with apple scar skin disease in the northeast of Iran. Iran. J. Virol. 2015, 9, 25–28. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Name a | Genome Segment (RNA) | Genome Size (kb) | Sequence Similarity (%) | Accession No. Received | UTR-5′ | ORF1 b | ORF2 | ORF3 | ORF4 | ORF5 | UTR-3′ | Reference Accession |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ACLSV | 1 | 7555 | 88 | OR537850 | 1–151 | 152–5809 (Replicase) | 5718–7100 (MP) | 6784–7365 (CP) | 7366–7550 | M58152 | ||
| AGCaV | 1 | 9266 | 90 | OR537851 | 1–60 | 61–6612 (Replicase) | 6697–7350 (TGB1) | 7352–7714 (TGB2) | 7623–7835 (TGB3) | 7924–9135 (CP) | 9136–9266 | HE963831 |
| AVCaV | 1 | 8358 | 98 | OR537852 | 1–78 | 79–6144 (Replicase) | 6144–7526 (MP) | 7057–7722 (CP) | 7790–8209 (Nucleic acid binding) | 8209–8358 | KM507062 | |
| ASPV | 1 | 9258 | 86 | OR537853 | 1–55 | 56–6603 (Replicase) | 6702–7373 (TGB1) | 7375–7737 (TGB2) | 7646–7873 (TGB3) | 7946–9136 (CP) | 9137–9258 | MZ148076 |
| ASGV | 1 | 6486 | 98 | OR537854 | 1–28 | 29–6346 (Replicae and CP) | 4780–5742 (MP) | 6354–6486 | MZ126527 | |||
| CCGaV | 1 | 6658 | 99 | OR537855 | 1–70 | 71–6625 (RdRp) | 6625–6658 | MZ926713 | ||||
| 2 | 2706 | 99 | OR537856 | 1–52 | 53–1276 (MP) | 1580–2632 (NP) | 2632–2706 | MZ926714 | ||||
| ApMV | 1 | 3440 | 89 | OR537857 | 1–62 | 63–3206 (RdRp) | 3207–3440 | HE574162 | ||||
| 2 | 2979 | 90 | OR537858 | 1–79 | 80–2707 (RdRp) | 2708–2979 | HE574163 | |||||
| 3 | 2056 | 90 | OR537859 | 1–168 | 169–1029 (MP) | 1126–1794 (CP) | 1795–2056 | HE574164 | ||||
| AHVd | 1 | 435 | 92 | OR537860 | - | - | - | - | - | - | - | KR605506 |
| HSVd | 1 | 300 | 98 | OR537861 | - | - | - | - | - | - | - | MF576422 |
| Province | Tested Samples a (n) | AGCaV (n/%) | ACLS (n/%) | AVCaV (n/%) | ASGV (n/%) | ASPV (n/%) | CCGaV (n/%) | ApMV (n/%) | HSVd (n/%) | AHVd (n/%) | Mixed Infections b (n/%) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mazandaran | 56 | 39/69.6 | 32/57.1 | 10/17.8 | 10/17.8 | 11/19.6 | 4/7.1 | 3/5.3 | 18/32.1 | 11/19.6 | 30/53.5 |
| Semnan | 52 | 26/50.0 | 11/21.1 | 7/13.4 | 12/23.0 | 8/15.3 | 3/5.7 | 1/1.9 | 8/15.3 | 1/1.9 | 25/48.0 |
| Alborz | 32 | 9/28.1 | 21/65.6 | 15/46.8 | 4/12.5 | 3/9.3 | 1/3.1 | 2/6.2 | 13/40.6 | 8/25.0 | 23/72.0 |
| Tehran | 30 | 7/23.3 | 14/46.6 | 3/10.0 | 3/10.0 | 2/6.6 | 0/0.0 | 0/0.0 | 8/26.6 | 6/20.0 | 15/50.0 |
| Total | 170 | 81 | 78 | 35 | 29 | 24 | 8 | 6 | 47 | 26 | 114 |
| Mean % | — | 47.6 | 45.8 | 20.5 | 17.0 | 14.1 | 4.7 | 3.5 | 27.6 | 15.2 | 67.0 |
| Origin of the Samples | Cultivar | AGCaV | ACLSV | ASGV | ASPV | AVCaV | |||||
| No. Infected Samples | Symptoms | No. Infected Samples | Symptoms | No. Infected Samples | Symptoms | No. Infected Samples | Symptoms | No. Infected Samples | Symptoms | ||
| Mazandaran | Gala | 3 | Cl, Vc | 2 | NL, D | 1 | Vc, Cl | 3 | D, Vc | ND | - |
| Golden Delicious | 11 | D, Vc | 9 | NL, D | 3 | Vc, Cl | ND | - | 2 | Vc | |
| Red Delicious | 13 | D, Cl | 8 | CL, D | 4 | Vc, Cl | 1 | D, Cl | 5 | Vc, D | |
| Shafi Abadi | 7 | D, NL | 12 | D, NL | 3 | Vc, Cl | 5 | D, NL | 3 | Vc | |
| Starking | 2 | CL, Vc | 1 | Vc | ND | - | ND | - | ND | - | |
| Granny Smith | 3 | CL, Vc | ND | - | ND | - | 2 | D, Vc | ND | - | |
| Semnan | Gala | 1 | D, Vc | ND | - | ND | - | 3 | D, Vc | ND | - |
| Golden Delicious | 5 | M, Vc | 1 | CL | ND | - | 1 | CL, D, NL | 2 | Vc | |
| Red Delicious | 6 | D, NL | 3 | D, CL | 2 | Vc, NL | 2 | D, Vc | 4 | Vc, D | |
| Shafi Abadi | 10 | D, NL | 6 | D, Vc | 7 | Vc, Cl | 1 | - | 1 | Vc | |
| Starking | 2 | NL, D | ND | - | 2 | Vc, Cl | 1 | D, Vc | ND | - | |
| Granny Smith | 2 | CL | 1 | CL | 1 | Vc, Cl | ND | CL, D, NL | ND | - | |
| Alborz | Gala | ND | - | 3 | NL, D | ND | - | ND | - | ND | - |
| Golden Delicious | 2 | CL, Vc | 4 | D, Vc | 1 | Vc, Cl | ND | - | 6 | Vc | |
| Red Delicious | 1 | D, Cl | 5 | CL, D, NL | ND | - | 1 | D, CL | 6 | Vc, D | |
| Shafi Abadi | 3 | D, NL | 7 | D, Vc | 3 | Vc, Cl | 2 | D, CL | 3 | Vc | |
| Starking | 2 | D, Vc | ND | - | ND | - | ND | - | ND | - | |
| Granny Smith | 1 | Vc | 2 | CL, NL | ND | - | ND | - | ND | - | |
| Tehran | Gala | ND | - | ND | - | ND | - | ND | - | ND | - |
| Golden Delicious | 1 | D, Vc | 2 | D, NL | 1 | Vc, Cl | 1 | Vc, D | ND | Vc | |
| Red Delicious | 2 | D, CL | 4 | D, Vc | ND | - | 1 | Vc, Cl | 1 | Vc | |
| Shafi Abadi | 4 | D, NL | 6 | CL, NL, Vc | 2 | Vc, Cl | ND | - | 2 | Vc, D | |
| Starking | ND | - | ND | - | ND | - | ND | - | ND | - | |
| Granny Smith | ND | - | 2 | Vc, CL | ND | - | ND | - | ND | - | |
| Origin Of the samples | Cultivar | ApMV | CCGaV | AHVd | HSVd | ||||||
| No. Infected samples | Symptoms | No. Infected samples | Symptoms | No. Infected samples | Symptoms | No. Infected samples | Symptoms | ||||
| Mazandaran | Gala | ND | - | ND | - | 3 | CL | 2 | NL | ||
| Golden Delicious | ND | - | 1 | D | 1 | D, CL | 5 | D, CL | |||
| Red Delicious | 1 | CL, NL | 2 | D, Vc | 2 | NL | 4 | Vc, CL | |||
| Shafi Abadi | 2 | D, CL | 1 | D | 3 | D, CL | 5 | D, CL | |||
| Starking | ND | - | ND | - | 1 | D, CL | 1 | D | |||
| Granny Smith | ND | - | ND | - | 1 | D, CL | 1 | D, CL | |||
| Semnan | Gala | ND | - | ND | - | ND | - | ND | - | ||
| Golden Delicious | 1 | Vc, CL | 3 | D, NL | ND | - | 3 | D, NL | |||
| Red Delicious | ND | - | ND | - | ND | - | 2 | D, NL | |||
| Shafi Abadi | ND | - | ND | - | 1 | D, NL | 1 | CL, D | |||
| Starking | ND | - | ND | - | ND | - | 1 | CL | |||
| Granny Smith | ND | - | ND | - | ND | - | 1 | CL | |||
| Alborz | Gala | ND | - | ND | - | 2 | D, Vc | ND | - | ||
| Golden Delicious | 1 | D, CL, NL | 1 | D, NL | 1 | D, CL | 5 | CL, Vc | |||
| Red Delicious | ND | - | ND | - | 2 | D, CL, NL | 3 | CL, D | |||
| Shafi Abadi | 1 | D, CL | ND | - | 2 | D | 4 | CL, D | |||
| Starking | ND | - | ND | - | ND | - | ND | - | |||
| Granny Smith | ND | - | ND | - | 1 | D | 1 | CL | |||
| Tehran | Gala | ND | - | ND | - | ND | - | 1 | CL | ||
| Golden Delicious | ND | - | ND | - | 1 | CL, NL | 1 | CL | |||
| Red Delicious | ND | - | ND | - | ND | - | 2 | CL, D | |||
| Shafi Abadi | ND | - | ND | - | 3 | D, CL | 3 | CL, D, CL | |||
| Starking | ND | - | ND | - | ND | - | CL | NL | |||
| Granny Smith | ND | - | ND | - | 2 | D, NL | 1 | CL | |||
| ASGV (aa) | ASGV (nt) | ACLSV (aa) | ACLSV (nt) | AGCaV (aa) | AGCaV (nt) | ASGV c Accession | ACLSV b Accession | AGCaV a Accession |
|---|---|---|---|---|---|---|---|---|
| 100 | 99 | 92 | 84 | 85 | 84 | OR537854 (Tehran-A4, Iran) | OR537850 (Damavand-A2, Iran) | OR537851 (Alborz-A5, Iran) |
| 100 | 99 | 92 | 85 | 94 | 92 | OR537865 (Damavand-A1, Iran) | OR537862 (Damavand-A, Iran) | OR570871 (AL22, Iran) |
| 100 | 99 | 94 | 88 | 94 | 92 | OR537866 (Alborz, Iran) | OR537863 (Semnan-A4, Iran) | OR570872 (MA10, Iran) |
| 100 | 99 | 91 | 84 | 93 | 91 | OR537867 (Mazandaran-A5, Iran) | OR537864 (Mazandaran-A5, Iran) | OR570873 (SEM19, Iran) |
| 100 | 99 | 90 | 86 | 89 | 94 | OQ263371 (K, Iran) | MK354028 (KS, Iran) | KT835289 (CYD, Italy) |
| 100 | 99 | 90 | 87 | 85 | 82 | OQ263370 (MN18, Iran) | MK354029 (MS, Iran) | MW842992 (Royal-Gala, Brazil) |
| 100 | 99 | 92 | 84 | 100 | 100 | OQ718412 (L57, Laos) | MK354022 (KSS, Iran) | HE963831 (Aurora-1, Canada) |
| 100 | 99 | 91 | 86 | MK354031 (KZ, Iran) | KM586376 (T12JA, Iran) | |||
| 100 | 99 | 91 | 84 | MK354036 (X4, Iran) | KM586369 (T5Te, Iran) | |||
| 97 | 93 | 91 | 86 | KR185346 (Cuiguan, China) | KM586363 (R2T6Na, Iran) | |||
| 100 | 99 | 89 | 87 | MZ127820 (ASGV-MK, India) | MH534892 (DN11, China) | |||
| 100 | 99 | 88 | 84 | OR640152 (ASGV_T, Turkey) | OR640150 (ACLSV-T, Turkey) | |||
| 95 | 90 | 94 | 87 | LC184612 (Nagami, Japan) | AM494513 (Salooni, India) | |||
| 97 | 93 | 94 | 86 | LC480456 (BH-Cnidium, South Korea) | KY579382 (16–133A-LT, Lithuania) | |||
| 100 | 99 | 95 | 87 | MW322833 (ASGV/FT146, USA) | KX668485 (M075, Brazil) | |||
| 100 | 99 | 95 | 88 | MW582790 (DSMZPV-0199, Germany) | KX224102 (ACLSV-Eth1, Ethiopia) | |||
| 100 | 99 | 95 | 88 | AF438409 (UV01, Brazil) | ON641858 (LV-54, Latvia) | |||
| 98 | 90 | 94 | 86 | LC645073 (ASGV-AK6, Japan) | LC006864 (PKO, South Korea) | |||
| 100 | 100 | 96 | 88 | MZ126527 (13TF138, Canada) | MZ126517 (13TF180B, Canada) | |||
| 94 | 87 | LC533838 (RRACV2, Japan) | ||||||
| 95 | 87 | MK086027 (ACLSV-K2, Kazakhstan) | ||||||
| 95 | 88 | OQ731902 (DSMZ PV-1000, Germany) | ||||||
| 92 | 90 | ON641871 (UKR5–36, Ukraine) | ||||||
| 93 | 86 | HQ398253 (SK-92-pl, Slovakia) | ||||||
| 84 | 80 | EU223295 (TaTao5, USA) | ||||||
| 100 | 100 | M58152 (P863, USA) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamedi, A.; Rakhshandehroo, F.; Safarnejad, M.R.; Jouzani, G.S.; Ben Slimen, A.; Elbeaino, T. Comprehensive Virome Profiling of Apple Mosaic Disease-Affected Trees in Iran Using RT-PCR and Next-Generation Sequencing. Viruses 2025, 17, 979. https://doi.org/10.3390/v17070979
Hamedi A, Rakhshandehroo F, Safarnejad MR, Jouzani GS, Ben Slimen A, Elbeaino T. Comprehensive Virome Profiling of Apple Mosaic Disease-Affected Trees in Iran Using RT-PCR and Next-Generation Sequencing. Viruses. 2025; 17(7):979. https://doi.org/10.3390/v17070979
Chicago/Turabian StyleHamedi, Anahita, Farshad Rakhshandehroo, Mohammad Reza Safarnejad, Gholamreza Salehi Jouzani, Amani Ben Slimen, and Toufic Elbeaino. 2025. "Comprehensive Virome Profiling of Apple Mosaic Disease-Affected Trees in Iran Using RT-PCR and Next-Generation Sequencing" Viruses 17, no. 7: 979. https://doi.org/10.3390/v17070979
APA StyleHamedi, A., Rakhshandehroo, F., Safarnejad, M. R., Jouzani, G. S., Ben Slimen, A., & Elbeaino, T. (2025). Comprehensive Virome Profiling of Apple Mosaic Disease-Affected Trees in Iran Using RT-PCR and Next-Generation Sequencing. Viruses, 17(7), 979. https://doi.org/10.3390/v17070979