

Trichomonas vaginalis Virus: Current Insights and Emerging Perspectives

{kind=link}
Abstract
1. Introduction
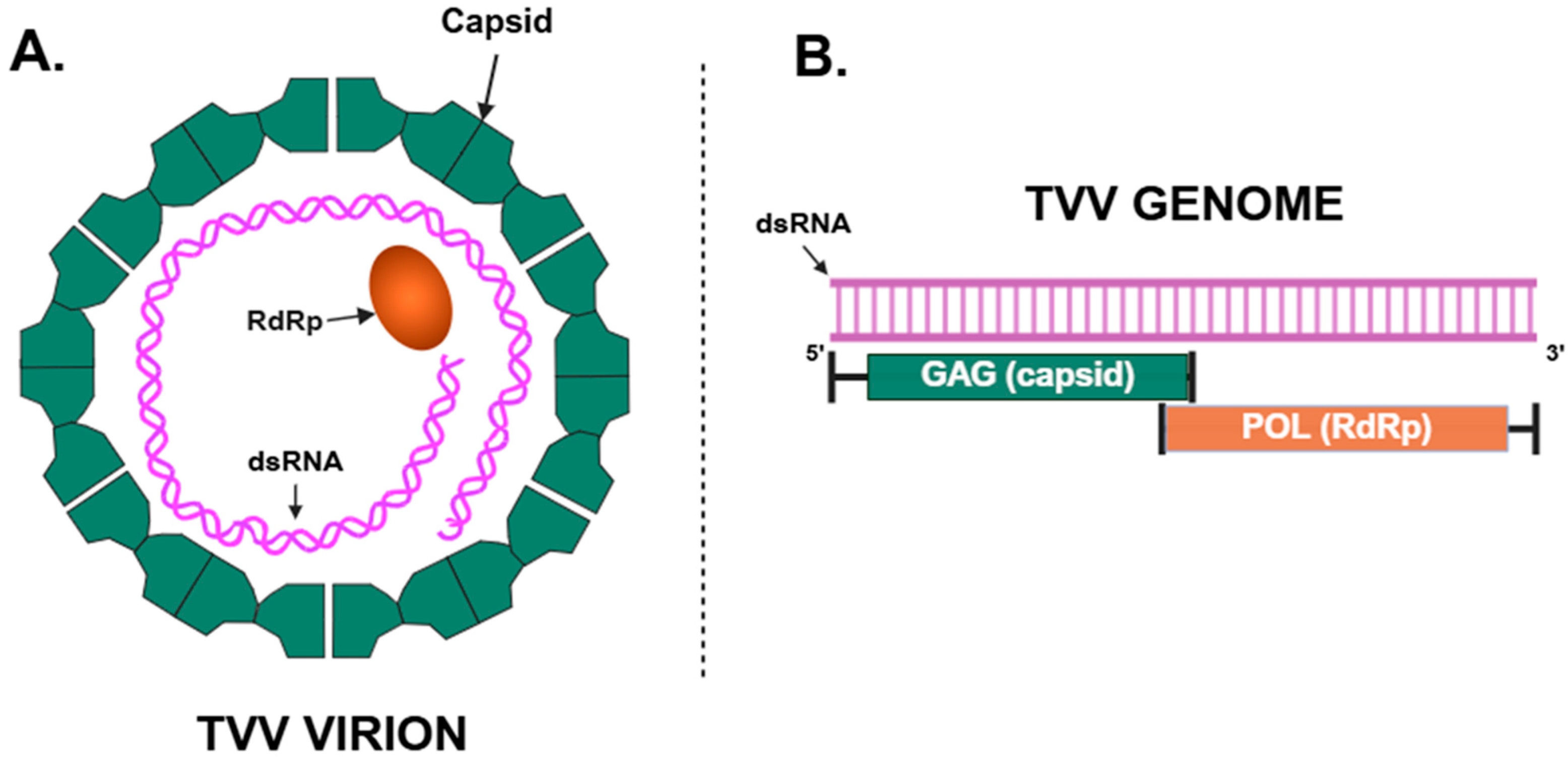
2. Genetic Structure and Transmission of TVV
3. Impact of TVV on T. vaginalis Protein Expression
3.1. Cysteine Proteases
3.2. Surface Antigens
3.3. Other Proteins
4. 5-Nitroimidazole Drug Susceptibility
5. Clinical Significance
6. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Rowley, J.; Vander Hoorn, S.; Korenromp, E.; Low, N.; Unemo, M.; Abu-Raddad, L.J.; Chico, R.M.; Smolak, A.; Newman, L.; Gottlieb, S.; et al. Chlamydia, gonorrhoea, trichomoniasis and syphilis: Global prevalence and incidence estimates, 2016. Bull. World Health Organ. 2019, 97, 548–562. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Liu, L.; Liu, K.; Qin, X.; Wu, J.; Jiang, L.; Shang, L. Global burden of trichomoniasis: Current status, trends, and projections (1990–2021). Front. Public. Health 2025, 13, 1530227. [Google Scholar] [CrossRef] [PubMed]
- Patel, E.U.; Gaydos, C.A.; Packman, Z.R.; Quinn, T.C.; Tobian, A.A.R. Prevalence and correlates of Trichomonas vaginalis infection among men and women in the United States. Clin. Infect. Dis. 2018, 67, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Van Gerwen, O.T.; Craig-Kuhn, M.C.; Jones, A.T.; Schroeder, J.A.; Deaver, J.; Buekens, P.; Kissinger, P.J.; Muzny, C.A. Trichomoniasis and adverse birth outcomes: A systematic review and meta-analysis. BJOG Int. J. Obstet. Gynaecol. 2021, 128, 1907–1915. [Google Scholar] [CrossRef]
- Masha, S.C.; Cools, P.; Sanders, E.J.; Vaneechoutte, M.; Crucitti, T. Trichomonas vaginalis and HIV infection acquisition: A systematic review and meta-analysis. Sex. Transm. Infect. 2019, 95, 36–42. [Google Scholar] [CrossRef]
- Allsworth, J.E.; Ratner, J.A.; Peipert, J.F. Trichomoniasis and other sexually transmitted infections: Results from the 2001–2004 National Health and Nutrition Examination Surveys. Sex. Transm. Dis. 2009, 36, 738–744. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Y.; Lu, H.; Li, D.; Zhang, R.; Xie, X.; Guo, L.; Hao, L.; Tian, X.; Yang, Z.; et al. A systematic review of the correlation between Trichomonas vaginalis infection and infertility. Acta Trop. 2022, 236, 106693. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D.; Li, Y.; Zhang, R.; Xie, X.; Yao, Y.; Zhao, L.; Tian, X.; Yang, Z.; Wang, S.; et al. The correlation between Trichomonas vaginalis infection and reproductive system cancer: A systematic review and meta-analysis. Infect. Agent. Cancer 2023, 18, 15. [Google Scholar] [CrossRef]
- Muzny, C.A.; George, S.; Kissinger, P.J.; Van Gerwen, O.T. Trichomoniasis and other sexually transmitted parasitic diseases in women. Clin. Obstet. Gynecol. 2025, 68, 194–205. [Google Scholar] [CrossRef]
- Workowski, K.A.; Bachmann, L.H.; Chan, P.A.; Johnston, C.M.; Muzny, C.A.; Park, I.; Reno, H.; Zenilman, J.M.; Bolan, G.A. Sexually transmitted infections treatment guidelines, 2021. MMWR Recomm. Rep. 2021, 70, 1–187. [Google Scholar]
- Mbizvo, E.M.; Msuya, S.E.; Stray-Pedersen, B.; Sundby, J.; Chirenje, Z.M.; Hussain, A. Determinants of reproductive tract infections among asymptomatic women in Harare, Zimbabwe. Cent. Afr. J. Med. 2001, 47, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Wiringa, A.E.; Ness, R.B.; Darville, T.; Beigi, R.H.; Haggerty, C.L. Trichomonas vaginalis, endometritis and sequelae among women with clinically suspected pelvic inflammatory disease. Sex. Transm. Infect. 2020, 96, 436–438. [Google Scholar] [CrossRef]
- Van Gerwen, O.T.; Camino, A.F.; Sharma, J.; Kissinger, P.J.; Muzny, C.A. Epidemiology, natural history, diagnosis, and treatment of Trichomonas vaginalis in Men. Clin. Infect. Dis. 2021, 73, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Van Gerwen, O.T.; Opsteen, S.A.; Graves, K.J.; Muzny, C.A. Trichomoniasis. Infect. Dis. Clin. N. Am. 2023, 37, 245–265. [Google Scholar] [CrossRef]
- Kissinger, P.; Mena, L.; Levison, J.; Clark, R.A.; Gatski, M.; Henderson, H.; Schmidt, N.; Rosenthal, S.L.; Myers, L.; Martin, D.H. A randomized treatment trial: Single versus 7-day dose of metronidazole for the treatment of Trichomonas vaginalis among HIV-infected women. J. Acquir. Immune Defic. Syndr. 2010, 55, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Kissinger, P.; Muzny, C.A.; Mena, L.A.; Lillis, R.A.; Schwebke, J.R.; Beauchamps, L.; Taylor, S.N.; Schmidt, N.; Myers, L.; Augostini, P.; et al. Single-dose versus 7-day-dose metronidazole for the treatment of trichomoniasis in women: An open-label, randomised controlled trial. Lancet Infect. Dis. 2018, 18, 1251–1259. [Google Scholar] [CrossRef]
- Muzny, C.A.; Schwebke, J.R.; Nyirjesy, P.; Kaufman, G.; Mena, L.A.; Lazenby, G.B.; Van Gerwen, O.T.; Graves, K.J.; Arbuckle, J.; Carter, B.A.; et al. Efficacy and safety of single oral dosing of secnidazole for trichomoniasis in women: Results of a phase 3, randomized, double-blind, placebo-controlled, delayed-treatment study. Clin. Infect. Dis. 2021, 73, e1282–e1289. [Google Scholar] [CrossRef]
- Muzny, C.A.; Van Gerwen, O.T.; Legendre, D. Secnidazole: A treatment for trichomoniasis in adolescents and adults. Expert Rev. Anti Infect. Ther. 2022, 20, 1067–1076. [Google Scholar] [CrossRef]
- Kirkcaldy, R.D.; Augostini, P.; Asbel, L.E.; Bernstein, K.T.; Kerani, R.P.; Mettenbrink, C.J.; Pathela, P.; Schwebke, J.R.; Secor, W.E.; Workowski, K.A.; et al. Trichomonas vaginalis antimicrobial drug resistance in 6 US cities, STD Surveillance Network, 2009–2010. Emerg. Infect. Dis. 2012, 18, 939–943. [Google Scholar] [CrossRef]
- Conrad, M.D.; Gorman, A.W.; Schillinger, J.A.; Fiori, P.L.; Arroyo, R.; Malla, N.; Dubey, M.L.; Gonzalez, J.; Blank, S.; Secor, W.E.; et al. Extensive genetic diversity, unique population structure and evidence of genetic exchange in the sexually transmitted parasite Trichomonas vaginalis. PLoS Negl. Trop. Dis. 2012, 6, e1573. [Google Scholar] [CrossRef]
- Wang, A.L.; Wang, C.C. A linear double-stranded RNA in Trichomonas vaginalis. J. Biol. Chem. 1985, 260, 3697–3702. [Google Scholar] [CrossRef]
- Wang, A.L.; Wang, C.C. The double-stranded RNA in Trichomonas vaginalis may originate from virus-like particles. Proc. Natl. Acad. Sci. USA 1986, 83, 7956–7960. [Google Scholar] [CrossRef]
- Graves, K.J.; Ghosh, A.P.; Kissinger, P.J.; Muzny, C.A. Trichomonas vaginalis virus: A review of the literature. Int. J. STD AIDS 2019, 30, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Parent, K.N.; Takagi, Y.; Cardone, G.; Olson, N.H.; Ericsson, M.; Yang, M.; Lee, Y.; Asara, J.M.; Fichorova, R.N.; Baker, T.S.; et al. Structure of a protozoan virus from the human genitourinary parasite Trichomonas vaginalis. mBio 2013, 4, e00056-13. [Google Scholar] [CrossRef] [PubMed]
- Fraga, J.; Rojas, L.; Sariego, I.; Fernandez-Calienes, A.; Nunez, F.A. Species typing of Cuban Trichomonas vaginalis virus by RT-PCR, and association of TVV-2 with high parasite adhesion levels and high pathogenicity in patients. Arch. Virol. 2012, 157, 1789–1795. [Google Scholar] [CrossRef]
- Goodman, R.P.; Freret, T.S.; Kula, T.; Geller, A.M.; Talkington, M.W.; Tang-Fernandez, V.; Suciu, O.; Demidenko, A.A.; Ghabrial, S.A.; Beach, D.H.; et al. Clinical isolates of Trichomonas vaginalis concurrently infected by strains of up to four Trichomonasvirus species (Family Totiviridae). J. Virol. 2011, 85, 4258–4270. [Google Scholar] [CrossRef] [PubMed]
- Manny, A.R.; Hetzel, C.A.; Mizani, A.; Nibert, M.L. Discovery of a novel species of Trichomonasvirus in the human parasite Trichomonas vaginalis using transcriptome mining. Viruses 2022, 14, 548. [Google Scholar] [CrossRef]
- Bahadory, S.; Aminizadeh, S.; Taghipour, A.; Bokharaei-Salim, F.; Khanaliha, K.; Razizadeh, M.H.; Soleimani, A.; Beikzadeh, L.; Khatami, A. A systematic review and meta-analysis on the global status of Trichomonas vaginalis virus in Trichomonas vaginalis. Microb. Pathog. 2021, 158, 105058. [Google Scholar] [CrossRef]
- Khoshnan, A.; Alderete, J.F. Multiple double-stranded RNA segments are associated with virus particles infecting Trichomonas vaginalis. J. Virol. 1993, 67, 6950–6955. [Google Scholar] [CrossRef]
- Benchimol, M.; Chang, T.H.; Alderete, J.F. Visualization of new virus-like-particles in Trichomonas vaginalis. Tissue Cell 2002, 34, 406–415. [Google Scholar] [CrossRef]
- Benchimol, M.; Monteiro, S.; Chang, T.H.; Alderete, J.F. Virus in Trichomonas—An ultrastructural study. Parasitol. Int. 2002, 51, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Khoshnan, A.; Provenzano, D.; Alderete, J.F. Unique double-stranded RNAs associated with the Trichomonas vaginalis virus are synthesized by viral RNA-dependent RNA polymerase. J. Virol. 1994, 68, 7108–7114. [Google Scholar] [CrossRef] [PubMed]
- Benchimol, M.; Chang, T.H.; Alderete, J.F. Trichomonas vaginalis: Observation of coexistence of multiple viruses in the same isolate. FEMS Microbiol. Lett. 2002, 215, 197–201. [Google Scholar] [CrossRef]
- Liu, H.W.; Chu, Y.D.; Tai, J.H. Characterization of Trichomonas vaginalis virus proteins in the pathogenic protozoan T. vaginalis. Arch. Virol. 1998, 143, 963–970. [Google Scholar] [CrossRef]
- Vanacova, S.; Tachezy, J.; Kulda, J.; Flegr, J. Characterization of trichomonad species and strains by PCR fingerprinting. J. Eukaryot. Microbiol. 1997, 44, 545–552. [Google Scholar] [CrossRef]
- Ong, S.C.; Cheng, W.H.; Ku, F.M.; Tsai, C.Y.; Huang, P.J.; Lee, C.C.; Yeh, Y.M.; Rada, P.; Hrdy, I.; Narayanasamy, R.K.; et al. Identification of endosymbiotic virus in small extracellular vesicles derived from Trichomonas vaginalis. Genes 2022, 13, 531. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, H.M.; Marcet, R.; Sarracent, J. Biological roles of cysteine proteinases in the pathogenesis of Trichomonas vaginalis. Parasite 2014, 21, 54. [Google Scholar] [CrossRef]
- Carlton, J.M.; Hirt, R.P.; Silva, J.C.; Delcher, A.L.; Schatz, M.; Zhao, Q.; Wortman, J.R.; Bidwell, S.L.; Alsmark, U.C.; Besteiro, S.; et al. Draft genome sequence of the sexually transmitted pathogen Trichomonas vaginalis. Science 2007, 315, 207–212. [Google Scholar] [CrossRef]
- Arroyo, R.; Cardenas-Guerra, R.E.; Figueroa-Angulo, E.E.; Puente-Rivera, J.; Zamudio-Prieto, O.; Ortega-Lopez, J. Trichomonas vaginalis cysteine proteinases: Iron response in gene expression and proteolytic activity. Biomed. Res. Int. 2015, 2015, 946787. [Google Scholar] [CrossRef]
- Miranda-Ozuna, J.F.T.; Rivera-Rivas, L.A.; Cardenas-Guerra, R.E.; Hernandez-Garcia, M.S.; Rodriguez-Cruz, S.; Gonzalez-Robles, A.; Chavez-Munguia, B.; Arroyo, R. Glucose-restriction increases Trichomonas vaginalis cellular damage towards HeLa cells and proteolytic activity of cysteine proteinases (CPs), such as TvCP2. Parasitology 2019, 146, 1156–1166. [Google Scholar] [CrossRef]
- Rivera-Rivas, L.A.; Arroyo, R. Iron restriction increases the expression of a cytotoxic cysteine proteinase TvCP2 by a novel mechanism of tvcp2 mRNA alternative polyadenylation in Trichomonas vaginalis. Biochim. Biophys. Acta Gene Regul. Mech. 2023, 1866, 194935. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Rivas, L.A.; Lorenzo-Benito, S.; Sanchez-Rodriguez, D.B.; Miranda-Ozuna, J.F.; Euceda-Padilla, E.A.; Ortega-Lopez, J.; Chavez-Munguia, B.; Lagunes-Guillen, A.; Velazquez-Valassi, B.; Jasso-Villazul, L.; et al. The effect of iron on Trichomonas vaginalis TvCP2: A cysteine proteinase found in vaginal secretions of trichomoniasis patients. Parasitology 2020, 147, 760–774. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Pengtao, G.; Ju, Y.; Jianhua, L.; He, L.; Guocai, Z.; Xichen, Z. Differential protein expressions in virus-infected and uninfected Trichomonas vaginalis. Korean J. Parasitol. 2017, 55, 121–128. [Google Scholar] [CrossRef]
- Provenzano, D.; Khoshnan, A.; Alderete, J.F. Involvement of dsRNA virus in the protein composition and growth kinetics of host Trichomonas vaginalis. Arch. Virol. 1997, 142, 939–952. [Google Scholar] [CrossRef] [PubMed]
- Alderete, J.F. Iron modulates phenotypic variation and phosphorylation of P270 in double-stranded RNA virus-infected Trichomonas vaginalis. Infect. Immun. 1999, 67, 4298–4302. [Google Scholar] [CrossRef]
- Zimmann, N.; Rada, P.; Zarsky, V.; Smutna, T.; Zahonova, K.; Dacks, J.; Harant, K.; Hrdy, I.; Tachezy, J. Proteomic analysis of Trichomonas vaginalis phagolysosome, lysosomal targeting, and unconventional secretion of cysteine peptidases. Mol. Cell Proteom. 2022, 21, 100174. [Google Scholar] [CrossRef]
- Musatovova, O.; Alderete, J.F. The Trichomonas vaginalis phenotypically varying P270 immunogen is highly conserved except for numbers of repeated elements. Microb. Pathog. 1999, 27, 93–104. [Google Scholar]
- Musatovova, O.; Alderete, J.F. Molecular analysis of the gene encoding the immunodominant phenotypically varying P270 protein of Trichomonas vaginalis. Microb. Pathog. 1998, 24, 223–239. [Google Scholar] [CrossRef]
- Dailey, D.C.; Alderete, J.F. The phenotypically variable surface protein of Trichomonas vaginalis has a single, tandemly repeated immunodominant epitope. Infect. Immun. 1991, 59, 2083–2088. [Google Scholar] [CrossRef]
- Alderete, J.F. Localization of the phenotypically varying P270 protein on dsRNA virus-positive and negative Trichomonas vaginalis isolates. Am. J. Biomed. Sci. Res. 2021, 14, 199–209. [Google Scholar] [CrossRef]
- Wang, A.; Wang, C.C.; Alderete, J.F. Trichomonas vaginalis phenotypic variation occurs only among trichomonads infected with the double-stranded RNA virus. J. Exp. Med. 1987, 166, 142–150. [Google Scholar] [CrossRef]
- Khoshnan, A.; Alderete, J.F. Trichomonas vaginalis with a double-stranded RNA virus has upregulated levels of phenotypically variable immunogen mRNA. J. Virol. 1994, 68, 4035–4038. [Google Scholar] [CrossRef]
- Alderete, J.F.; Kasmala, L.; Metcalfe, E.; Garza, G.E. Phenotypic variation and diversity among Trichomonas vaginalis isolates and correlation of phenotype with trichomonal virulence determinants. Infect. Immun. 1986, 53, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Stafkova, J.; Rada, P.; Meloni, D.; Zarsky, V.; Smutna, T.; Zimmann, N.; Harant, K.; Pompach, P.; Hrdy, I.; Tachezy, J. Dynamic secretome of Trichomonas vaginalis: Case study of beta-amylases. Mol. Cell Proteom. 2018, 17, 304–320. [Google Scholar] [CrossRef]
- Rada, P.; Kellerova, P.; Verner, Z.; Tachezy, J. Investigation of the secretory pathway in Trichomonas vaginalis argues against a moonlighting function of hydrogenosomal enzymes. J. Eukaryot. Microbiol. 2019, 66, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Twu, O.; de Miguel, N.; Lustig, G.; Stevens, G.C.; Vashisht, A.A.; Wohlschlegel, J.A.; Johnson, P.J. Trichomonas vaginalis exosomes deliver cargo to host cells and mediate hostratioparasite interactions. PLoS Pathog. 2013, 9, e1003482. [Google Scholar] [CrossRef]
- Kochanowsky, J.A.; Mira, P.M.; Elikaee, S.; Muratore, K.; Rai, A.K.; Riestra, A.M.; Johnson, P.J. Trichomonas vaginalis extracellular vesicles up-regulate and directly transfer adherence factors promoting host cell colonization. Proc. Natl. Acad. Sci. USA 2024, 121, e2401159121. [Google Scholar] [CrossRef]
- Rada, P.; Hrdy, I.; Zdrha, A.; Narayanasamy, R.K.; Smutna, T.; Horackova, J.; Harant, K.; Benes, V.; Ong, S.C.; Tsai, C.Y.; et al. Double-Stranded RNA viruses are released from Trichomonas vaginalis inside small extracellular vesicles and modulate the exosomal cargo. Front. Microbiol. 2022, 13, 893692. [Google Scholar] [CrossRef] [PubMed]
- Nievas, Y.R.; Coceres, V.M.; Midlej, V.; de Souza, W.; Benchimol, M.; Pereira-Neves, A.; Vashisht, A.A.; Wohlschlegel, J.A.; Johnson, P.J.; de Miguel, N. Membrane-shed vesicles from the parasite Trichomonas vaginalis: Characterization and their association with cell interaction. Cell. Mol. Life Sci. 2018, 75, 2211–2226. [Google Scholar] [CrossRef]
- Santana de Andrade, J.C.; Benchimol, M.; de Souza, W. Stimulation of microvesicle secretion in Trichomonas vaginalis. Exp. Parasitol. 2024, 259, 108722. [Google Scholar] [CrossRef]
- Artuyants, A.; Campos, T.L.; Rai, A.K.; Johnson, P.J.; Dauros-Singorenko, P.; Phillips, A.; Simoes-Barbosa, A. Extracellular vesicles produced by the protozoan parasite Trichomonas vaginalis contain a preferential cargo of tRNA-derived small RNAs. Int. J. Parasitol. 2020, 50, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Flegr, J.; Cerkasov, J.; Kulda, J.; Tachezy, J.; Stokrov, J. The dsRNA of Trichomonas vaginalis is associated with virus like particles and does not correlate with metronidazole resistance. Folia Microbiol. 1987, 32, 345–348. [Google Scholar] [CrossRef]
- Snipes, L.J.; Gamard, P.M.; Narcisi, E.M.; Beard, C.B.; Lehmann, T.; Secor, W.E. Molecular epidemiology of metronidazole resistance in a population of Trichomonas vaginalis clinical isolates. J. Clin. Microbiol. 2000, 38, 3004–3009. [Google Scholar] [CrossRef]
- Hampl, V.; Vanacova, S.; Kulda, J.; Flegr, J. Concordance between genetic relatedness and phenotypic similarities of Trichomonas vaginalis strains. BMC Evol. Biol. 2001, 1, 11. [Google Scholar] [CrossRef]
- Malla, N.; Kaul, P.; Sehgal, R.; Gupta, I. The presence of dsRNA virus in Trichomonas vaginalis isolates from symptomatic and asymptomatic Indian women and its correlation with in vitro metronidazole sensitivity. Indian. J. Med. Microbiol. 2011, 29, 152–157. [Google Scholar] [CrossRef] [PubMed]
- da Luz Becker, D.; dos Santos, O.; Frasson, A.P.; de Vargas Rigo, G.; Macedo, A.J.; Tasca, T. High rates of double-stranded RNA viruses and Mycoplasma hominis in Trichomonas vaginalis clinical isolates in South Brazil. Infect. Genet. Evol. 2015, 34, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Wendel, K.A.; Rompalo, A.M.; Erbelding, E.J.; Chang, T.H.; Alderete, J.F. Double-stranded RNA viral infection of Trichomonas vaginalis infecting patients attending a sexually transmitted diseases clinic. J. Infect. Dis. 2002, 186, 558–561. [Google Scholar] [CrossRef]
- Fraga, J.; Rojas, L.; Sariego, I.; Fernandez-Calienes, A.; Nunez, F.A. Double-Stranded RNA viral infection of Trichomonas vaginalis and association with clinical presentation. Acta Protozool. 2007, 46, 93–98. [Google Scholar]
- Fraga, J.; Rojas, L.; Sariego, I.; Fernandez-Calienes, A. Double-stranded RNA viral infection of Trichomonas vaginalis and correlation with genetic polymorphism of isolates. Exp. Parasitol. 2011, 127, 593–599. [Google Scholar] [CrossRef]
- El-Gayar, E.K.; Mokhtar, A.B.; Hassan, W.A. Molecular characterization of double-stranded RNA virus in Trichomonas vaginalis Egyptian isolates and its association with pathogenicity. Parasitol. Res. 2016, 115, 4027–4036. [Google Scholar] [CrossRef]
- Jehee, I.; van der Veer, C.; Himschoot, M.; Hermans, M.; Bruisten, S. Direct detection of Trichomonas vaginalis virus in Trichomonas vaginalis positive clinical samples from the Netherlands. J. Virol. Methods 2017, 250, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Fichorova, R.N.; Lee, Y.; Yamamoto, H.S.; Takagi, Y.; Hayes, G.R.; Goodman, R.P.; Chepa-Lotrea, X.; Buck, O.R.; Murray, R.; Kula, T.; et al. Endobiont viruses sensed by the human host—Beyond conventional antiparasitic therapy. PLoS ONE 2012, 7, e48418. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graves, K.J.; Novak, J.; Muzny, C.A. Trichomonas vaginalis Virus: Current Insights and Emerging Perspectives. Viruses 2025, 17, 898. https://doi.org/10.3390/v17070898
Graves KJ, Novak J, Muzny CA. Trichomonas vaginalis Virus: Current Insights and Emerging Perspectives. Viruses. 2025; 17(7):898. https://doi.org/10.3390/v17070898
Chicago/Turabian StyleGraves, Keonte J., Jan Novak, and Christina A. Muzny. 2025. "Trichomonas vaginalis Virus: Current Insights and Emerging Perspectives" Viruses 17, no. 7: 898. https://doi.org/10.3390/v17070898
APA StyleGraves, K. J., Novak, J., & Muzny, C. A. (2025). Trichomonas vaginalis Virus: Current Insights and Emerging Perspectives. Viruses, 17(7), 898. https://doi.org/10.3390/v17070898