
Cellular and Humoral Immune Profiles After Hepatitis E Vaccination and Infection

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
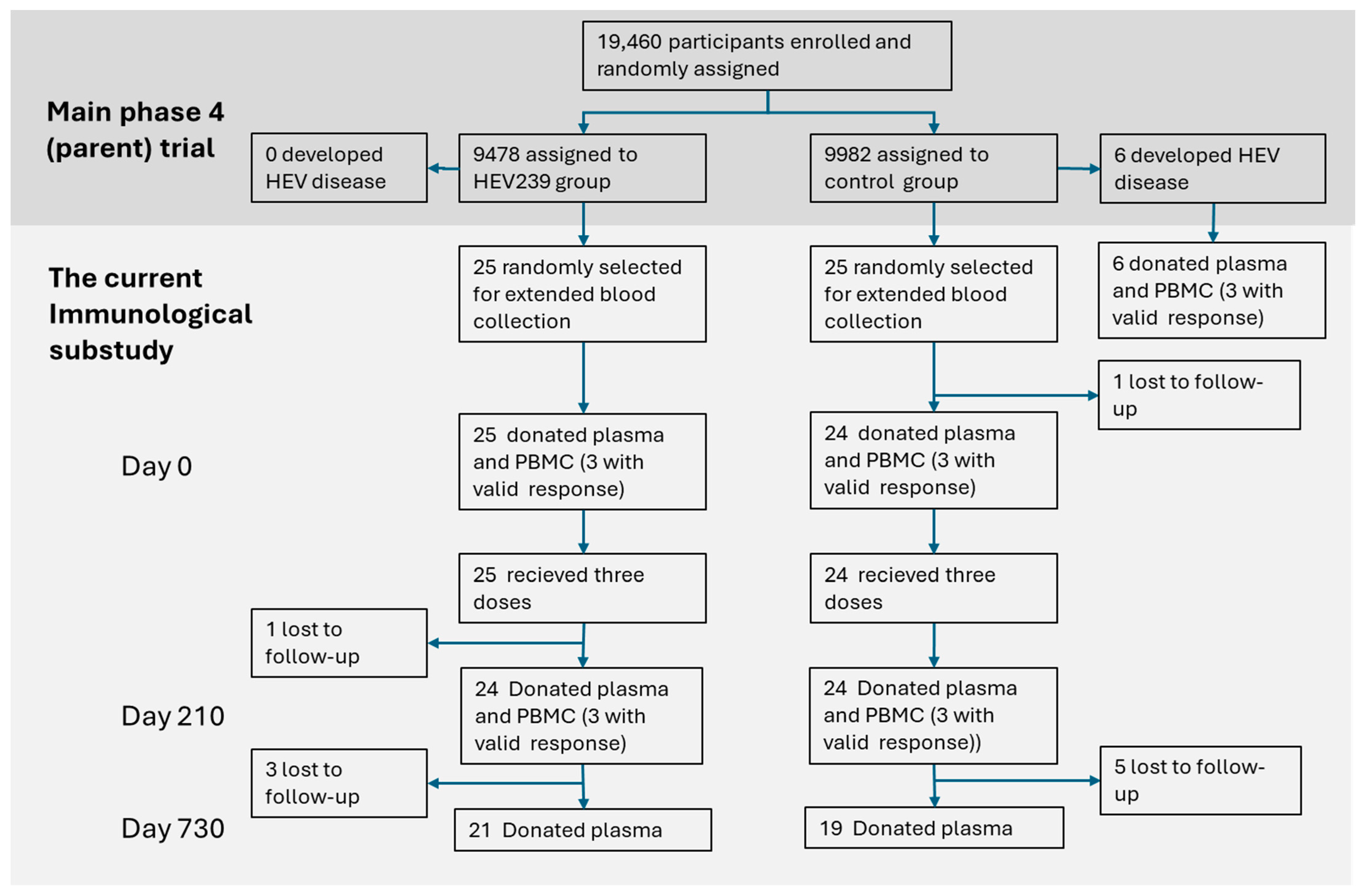
2.1. Study Population
2.2. Sample Collection and Processing
2.3. Thawing Procedure, Viability, and Recovery
2.4. ELISpot Assay (IFNγ/IL-4)
2.5. Serological Analyses
2.6. Statistical Analysis
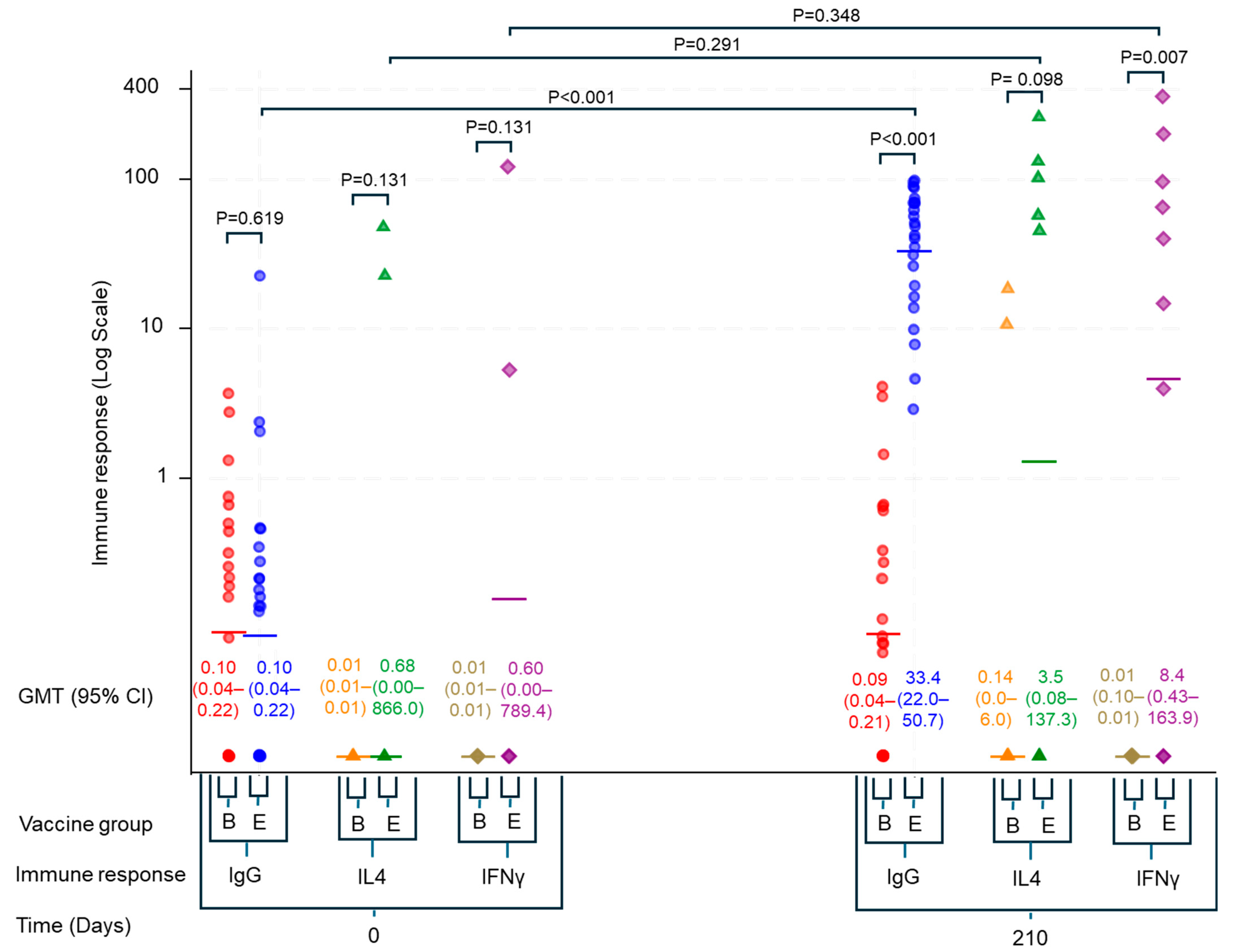
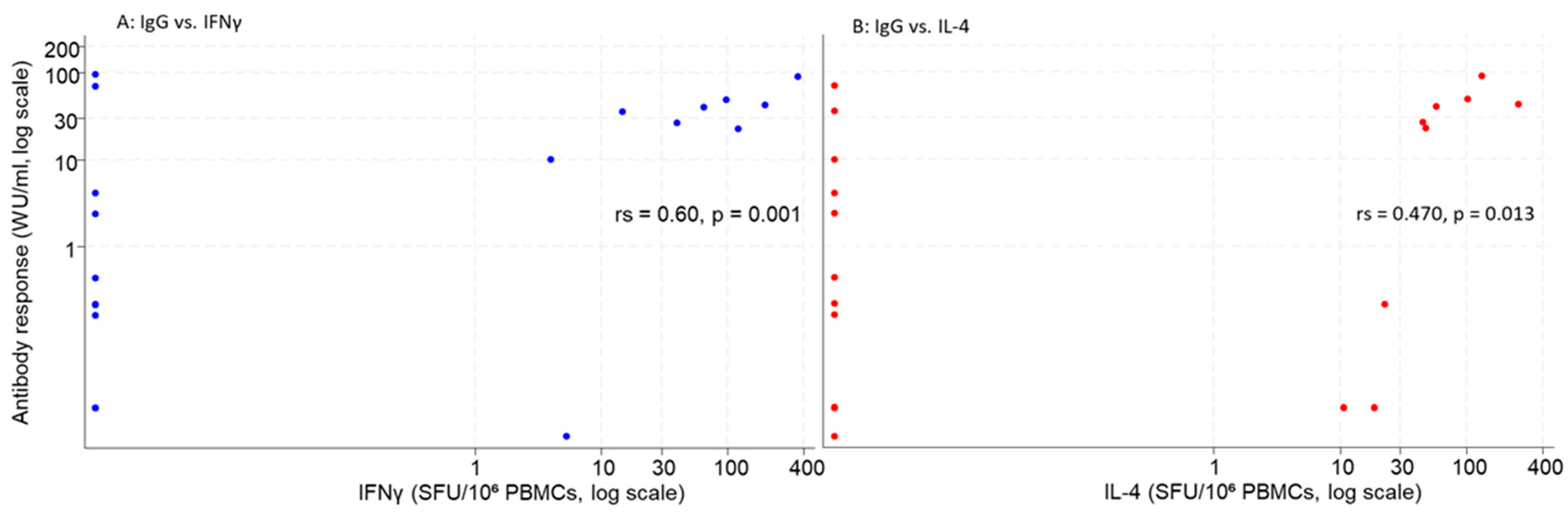
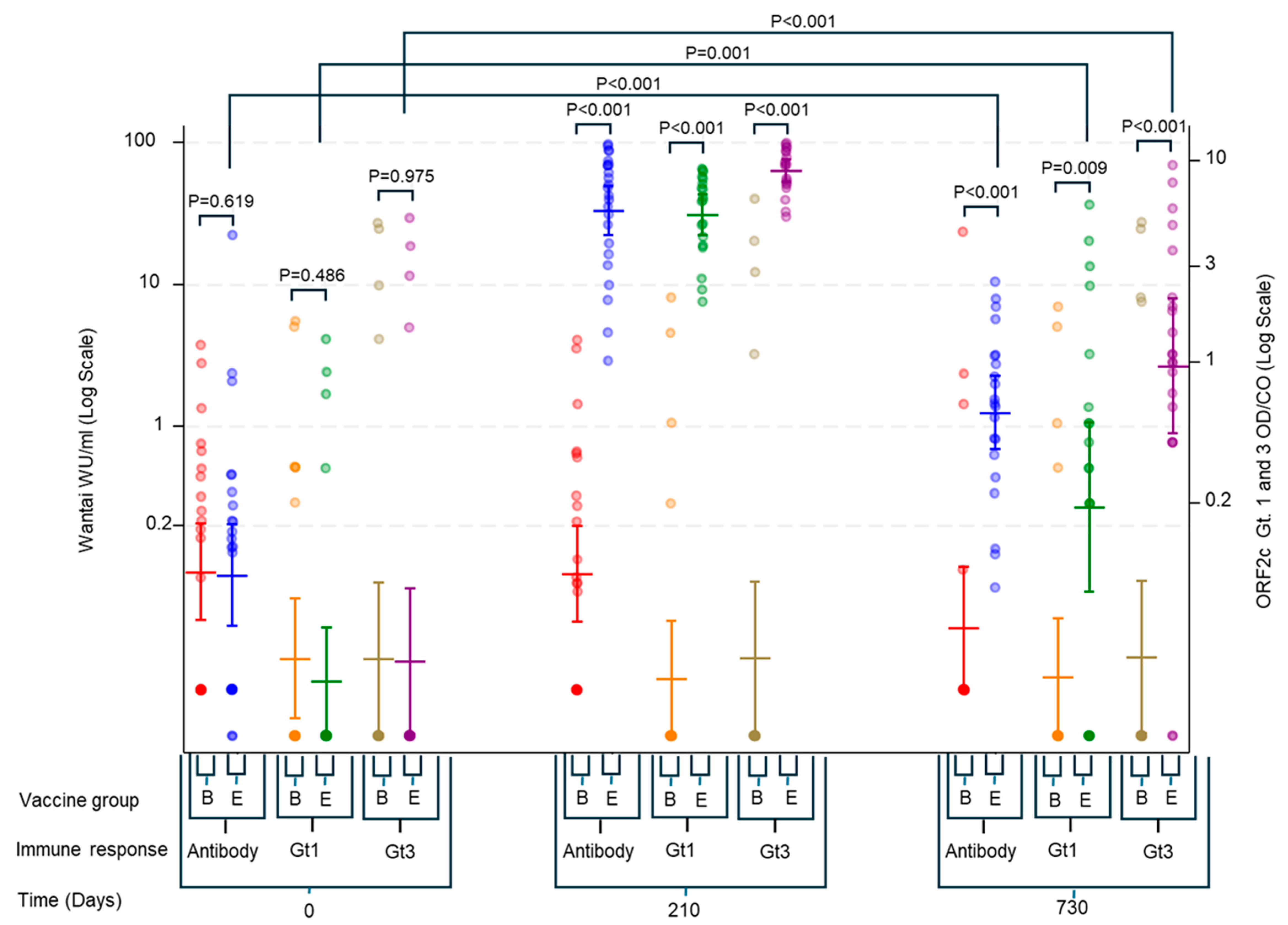
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamar, N.; Izopet, J.; Pavio, N.; Aggarwal, R.; Labrique, A.; Wedemeyer, H.; Dalton, H.R. Hepatitis e virus infection. Nat. Rev. Dis. Primers 2017, 3, 17086. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Mori, Y.; Miyazaki, N.; Cheng, R.H.; Yoshimura, M.; Unno, H.; Shima, R.; Moriishi, K.; Tsukihara, T.; Li, T.C.; et al. Biological and immunological characteristics of hepatitis e virus-like particles based on the crystal structure. Proc. Natl. Acad. Sci. USA 2009, 106, 12986–12991. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R.; Shukla, R.; Jameel, S.; Agrawal, S.; Puri, P.; Gupta, V.K.; Patil, A.P.; Naik, S. T-cell epitope mapping of orf2 and orf3 proteins of human hepatitis e virus. J. Viral Hepat. 2007, 14, 283–292. [Google Scholar] [CrossRef]
- Li, S.W.; Zhao, Q.; Wu, T.; Chen, S.; Zhang, J.; Xia, N.S. The development of a recombinant hepatitis e vaccine hev 239. Hum. Vaccines Immunother. 2015, 11, 908–914. [Google Scholar] [CrossRef]
- Muñoz-Chimeno, M.; Díaz-Sánchez, N.; Morago, L.; Rodríguez-Paredes, V.; Barturen, S.; Rodríguez-Recio, Á.; García-Lugo, M.A.; Zamora, M.I.; Mateo, M.; Sánchez-Martínez, M.; et al. Performance comparison of four hepatitis e antibodies detection methods. Microorganisms 2024, 12, 1875. [Google Scholar] [CrossRef]
- Simmonds, P.; Adriaenssens, E.M.; Lefkowitz, E.J.; Oksanen, H.M.; Siddell, S.G.; Zerbini, F.M.; Alfenas-Zerbini, P.; Aylward, F.O.; Dempsey, D.M.; Dutilh, B.E.; et al. Changes to virus taxonomy and the ictv statutes ratified by the international committee on taxonomy of viruses (2024). Arch. Virol. 2024, 169, 236. [Google Scholar] [CrossRef]
- Kamar, N.; Dalton, H.R.; Abravanel, F.; Izopet, J. Hepatitis e virus infection. Clin. Microbiol. Rev. 2014, 27, 116–138. [Google Scholar] [CrossRef]
- Ahmed, R.; Nasheri, N. Animal reservoirs for hepatitis e virus within the paslahepevirus genus. Vet. Microbiol. 2023, 278, 109618. [Google Scholar] [CrossRef]
- Chiereghin, A.; Verucchi, G.; Lazzarotto, T. Cmv-specific cell-mediated immunity in immunocompetent adults with primary cmv infection: A case series and review of the literature. Viruses 2021, 13, 816. [Google Scholar] [CrossRef]
- Dembinski, J.L.; Mihret, A.; Yimer, S.A.; Tessema, B.; Trieu, M.C.; Tarekegn, A.; Getachew, N.; Cox, R.J.; Oftung, F.; Haneberg, B.; et al. High prevalence of humoral and cellular immunity to influenza viruses in preschool children living in addis ababa, ethiopia. Open Forum Infect. Dis. 2017, 4, ofx026. [Google Scholar] [CrossRef]
- Srivastava, R.; Aggarwal, R.; Jameel, S.; Puri, P.; Gupta, V.K.; Ramesh, V.S.; Bhatia, S.; Naik, S. Cellular immune responses in acute hepatitis e virus infection to the viral open reading frame 2 protein. Viral Immunol 2007, 20, 56–65. [Google Scholar] [CrossRef]
- Taherkhani, R.; Farshadpour, F.; Makvandi, M.; Memari, H.R.; Samarbafzadeh, A.R.; Sharifi, N.; Naeimi, B.; Tajbakhsh, S.; Akbarzadeh, S. Cytokine profiles and cell proliferation responses to truncated orf2 protein in iranian patients recovered from hepatitis e infection. J. Trop. Med. 2015, 2015, 523560. [Google Scholar] [CrossRef]
- Clerici, M.; Shearer, G.M. A th1-->th2 switch is a critical step in the etiology of hiv infection. Immunol. Today 1993, 14, 107–111. [Google Scholar] [CrossRef]
- Walker, C.M. Adaptive immune responses in hepatitis a virus and hepatitis e virus infections. Cold Spring Harb. Perspect. Med. 2019, 9, a033472. [Google Scholar] [CrossRef]
- Kemming, J.; Gundlach, S.; Panning, M.; Huzly, D.; Huang, J.; Lütgehetmann, M.; Pischke, S.; Wiesch, J.S.Z.; Emmerich, F.; Llewellyn-Lacey, S.; et al. Mechanisms of cd8+ t-cell failure in chronic hepatitis e virus infection. J. Hepatol. 2022, 77, 978–990. [Google Scholar] [CrossRef]
- Wu, T.; Wu, X.L.; Ou, S.H.; Lin, C.X.; Cheng, T.; Li, S.W.; Ng, M.H.; Zhang, J.; Xia, N.S. Difference of t cell and b cell activation in two homologous proteins with similar antigenicity but great distinct immunogenicity. Mol. Immunol. 2007, 44, 3261–3266. [Google Scholar] [CrossRef]
- Øverbø, J.; Aziz, A.; Zaman, K.; Clemens, J.; Julin, C.H.; Qadri, F.; Stene-Johansen, K.; Biswas, R.; Islam, S.; Bhuiyan, T.R.; et al. Immunogenicity and safety of a two-dose regimen with hepatitis e virus vaccine in healthy adults in rural bangladesh: A randomized, double-blind, controlled, phase 2/pilot trial. Vaccine 2023, 41, 1059–1066. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, X.; Su, Y.; Zhuang, C.; Tang, Z.; Huang, X.; Chen, Q.; Zhu, K.; Hu, X.; Ying, D.; et al. Long-term efficacy of a recombinant hepatitis e vaccine in adults: 10-year results from a randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2024, 403, 813–823. [Google Scholar] [CrossRef]
- Wen, G.-P.; He, L.; Tang, Z.-M.; Wang, S.-L.; Zhang, X.; Chen, Y.-Z.; Lin, X.; Liu, C.; Chen, J.-X.; Ying, D.; et al. Quantitative evaluation of protective antibody response induced by hepatitis e vaccine in humans. Nat. Commun. 2020, 11, 3971. [Google Scholar] [CrossRef]
- Zaman, K.; Julin, C.H.; Aziz, A.B.; Stene-Johansen, K.; Yunus, M.; Qadri, F.; Gurley, E.S.; Sandbu, S.; Øverbø, J.; Dembinski, J.L.; et al. Safety and effectiveness of a recombinant hepatitis e vaccine in women of childbearing age in rural bangladesh: A phase 4, double-blind, cluster-randomised, controlled trial. Lancet Glob. Health 2024, 12, e1288–e1299. [Google Scholar] [CrossRef]
- Zaman, K.; Dudman, S.; Stene-Johansen, K.; Qadri, F.; Yunus, M.; Sandbu, S.; Gurley, E.S.; Overbo, J.; Julin, C.H.; Dembinski, J.L.; et al. Hev study protocol: Design of a cluster-randomised, blinded trial to assess the safety, immunogenicity and effectiveness of the hepatitis e vaccine hev 239 (hecolin) in women of childbearing age in rural bangladesh. BMJ Open 2020, 10, e033702. [Google Scholar] [CrossRef]
- Mauthe, A.; Cedrone, E.; Villar-Hernández, R.; Rusch, E.; Springer, M.; Schuster, M.; Preyer, R.; Dobrovolskaia, M.A.; Gutekunst, M. Ifn-γ/il-2 double-color fluorospot assay for monitoring human primary t cell activation: Validation, inter-laboratory comparison, and recommendations for clinical studies. AAPS J. 2025, 27, 81. [Google Scholar] [CrossRef]
- Gottschalk, P.G.; Dunn, J.R. The five-parameter logistic: A characterization and comparison with the four-parameter logistic. Anal. Biochem. 2005, 343, 54–65. [Google Scholar] [CrossRef]
- Csernalabics, B.; Marinescu, M.S.; Maurer, L.; Kelsch, L.; Werner, J.; Baumann, K.; Zoldan, K.; Panning, M.; Reuken, P.; Bruns, T.; et al. Efficient formation and maintenance of humoral and cd4 t-cell immunity targeting the viral capsid in acute-resolving hepatitis e infection. J. Hepatol. 2024, 80, 564–575. [Google Scholar] [CrossRef]
- Wu, J.; Guo, Y.; Lu, X.; Huang, F.; Lv, F.; Wei, D.; Shang, A.; Yang, J.; Pan, Q.; Jiang, B.; et al. Th1/th2 cells and associated cytokines in acute hepatitis e and related acute liver failure. J. Immunol. Res. 2020, 2020, 6027361. [Google Scholar] [CrossRef]
- Gu, Y.; Tang, X.; Zhang, X.; Song, C.; Zheng, M.; Wang, K.; Zhang, J.; Ng, M.H.; Hew, C.L.; Li, S.; et al. Structural basis for the neutralization of hepatitis e virus by a cross-genotype antibody. Cell Res. 2015, 25, 604–620. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, H.; Harrison, T.J.; Lang, S.; Huang, G.; Wang, Y. Cross-protection of hepatitis e virus genotypes 1 and 4 in rhesus macaques. J. Med. Virol. 2008, 80, 824–832. [Google Scholar] [CrossRef]
- Dähnert, L.; Aliabadi, E.; Fast, C.; Hrabal, I.; Schröder, C.; Behrendt, P.; Protzer, U.; Groschup, M.H.; Eiden, M. Immunisation of pigs with recombinant hev vaccines does not protect from infection with hev genotype 3. One Health 2024, 18, 100674. [Google Scholar] [CrossRef]
- Germann, A.; Oh, Y.-J.; Schmidt, T.; Schön, U.; Zimmermann, H.; von Briesen, H. Temperature fluctuations during deep temperature cryopreservation reduce pbmc recovery, viability and t-cell function. Cryobiology 2013, 67, 193–200. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant | Vaccine | Immune Response | Day 0 | Day 210 |
|---|---|---|---|---|
| 1 | HEV | WU/mL | 2.40 | 48.66 |
| ORF2c gt.1 | 0.7 | 9.2 | ||
| ORF2c gt.3 | 2.7 | 12.2 | ||
| IL4 | 0.014 | 102.67 | ||
| IFNγ | 0.014 | 97.34 | ||
| 2 | HEV | WU/mL | 0.19 | 9.97 |
| ORF2c gt.1 | 0.014 | 4.2 | ||
| ORF2c gt.3 | 0.014 | 8.0 | ||
| IL4 | 0.014 | 22.66 | ||
| IFNγ | 0.014 | 4.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Øverbø, J.; Dembinski, J.L.; Nilsen, T.R.; Sriranganathan, V.; Dimova-Svetoslavova, V.P.; Aziz, A.; Zaman, K.; Julin, C.H.; Qadri, F.; Stene-Johansen, K.; et al. Cellular and Humoral Immune Profiles After Hepatitis E Vaccination and Infection. Viruses 2025, 17, 901. https://doi.org/10.3390/v17070901
Øverbø J, Dembinski JL, Nilsen TR, Sriranganathan V, Dimova-Svetoslavova VP, Aziz A, Zaman K, Julin CH, Qadri F, Stene-Johansen K, et al. Cellular and Humoral Immune Profiles After Hepatitis E Vaccination and Infection. Viruses. 2025; 17(7):901. https://doi.org/10.3390/v17070901
Chicago/Turabian StyleØverbø, Joakim, Jennifer L. Dembinski, Toril Ranneberg Nilsen, Vethanayaki Sriranganathan, Veselka Petrova Dimova-Svetoslavova, Asma Aziz, K Zaman, Cathinka Halle Julin, Firdausi Qadri, Kathrine Stene-Johansen, and et al. 2025. "Cellular and Humoral Immune Profiles After Hepatitis E Vaccination and Infection" Viruses 17, no. 7: 901. https://doi.org/10.3390/v17070901
APA StyleØverbø, J., Dembinski, J. L., Nilsen, T. R., Sriranganathan, V., Dimova-Svetoslavova, V. P., Aziz, A., Zaman, K., Julin, C. H., Qadri, F., Stene-Johansen, K., Bhuiyan, T. R., Haque, W., & Dudman, S. (2025). Cellular and Humoral Immune Profiles After Hepatitis E Vaccination and Infection. Viruses, 17(7), 901. https://doi.org/10.3390/v17070901