
Anti-Herpes Simplex Virus (Wild-Type and Drug-Resistant) Properties of Herbal KerraTM, KSTM, and MinozaTM

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. The Extraction of KerraTM, KSTM, and MinozaTM
2.3. Viruses
2.4. Cytotoxicity
2.5. Plaque Assay
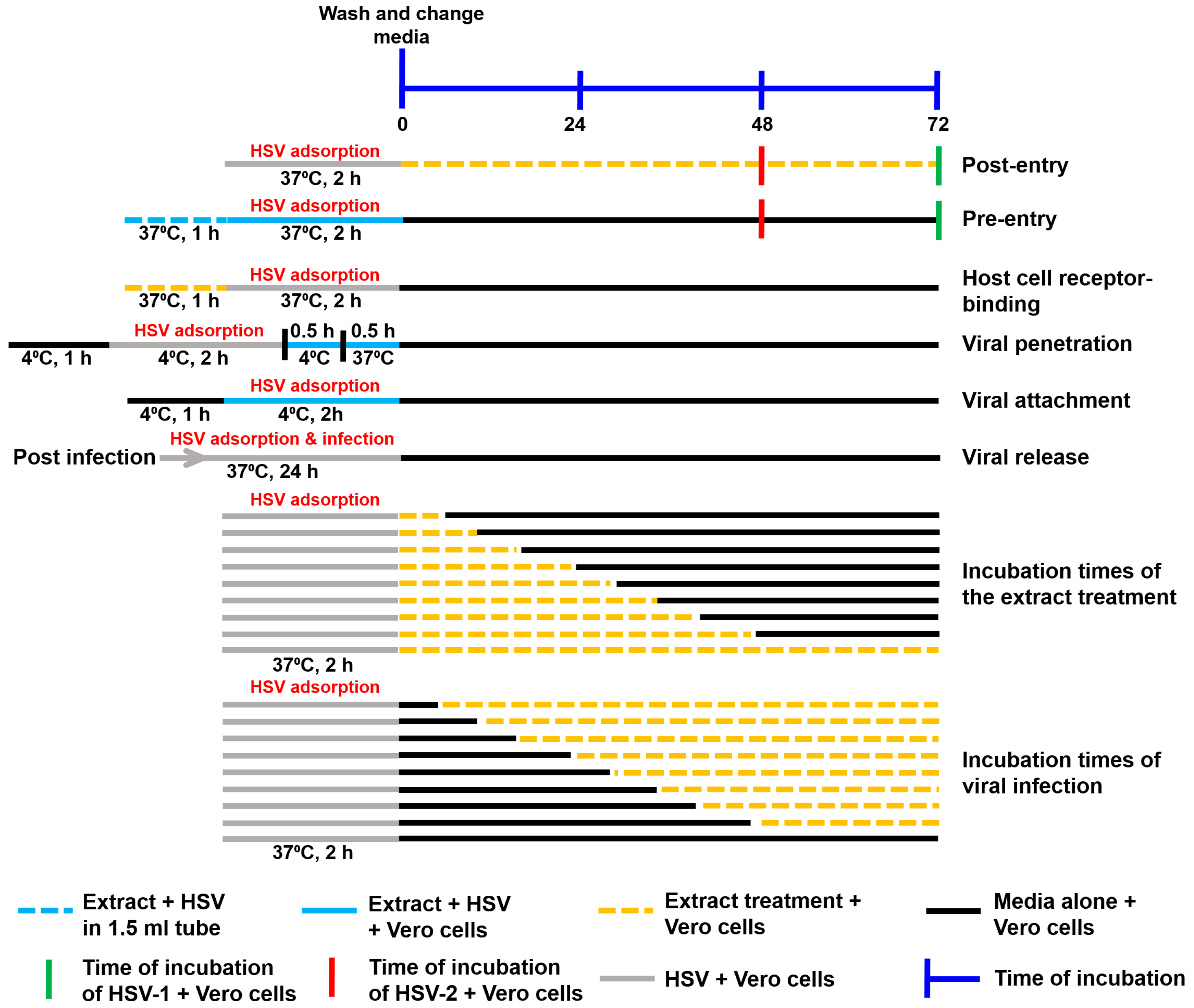
2.5.1. Post-Entry Step
2.5.2. Pre-Entry Step
2.5.3. Host Cell Receptor-Binding Assay
2.5.4. Viral Penetration
2.5.5. Viral Attachment
2.5.6. Viral Release Assay
2.5.7. Incubation Times of the Extract Treatment
2.5.8. Incubation Times of Viral Infection
2.6. Evaluation of Viral ICP4, UL30, and gD and Cellular IL1B, IL6, STAT3, and NFKB1 mRNA Expression
2.7. HSV-1 gB Protein Expression Detection by Western Blotting Analysis
2.8. Phytochemical Profile Analysis Using Liquid Chromatography–Tandem Mass Spectrometry (LC–MS/MS)
2.9. Molecular Docking Analysis and Potential Drug Target Prediction of 2-Methoxy-9H-xanthen-9-one, Isorhapontigeninfound, Piperine, Pellitorine, Sargassopenilline D, and Parmoether A
2.10. Statistical Analysis
3. Results
3.1. Cytotoxicity of KerraTM, KSTM, and MinozaTM in Vero Cells
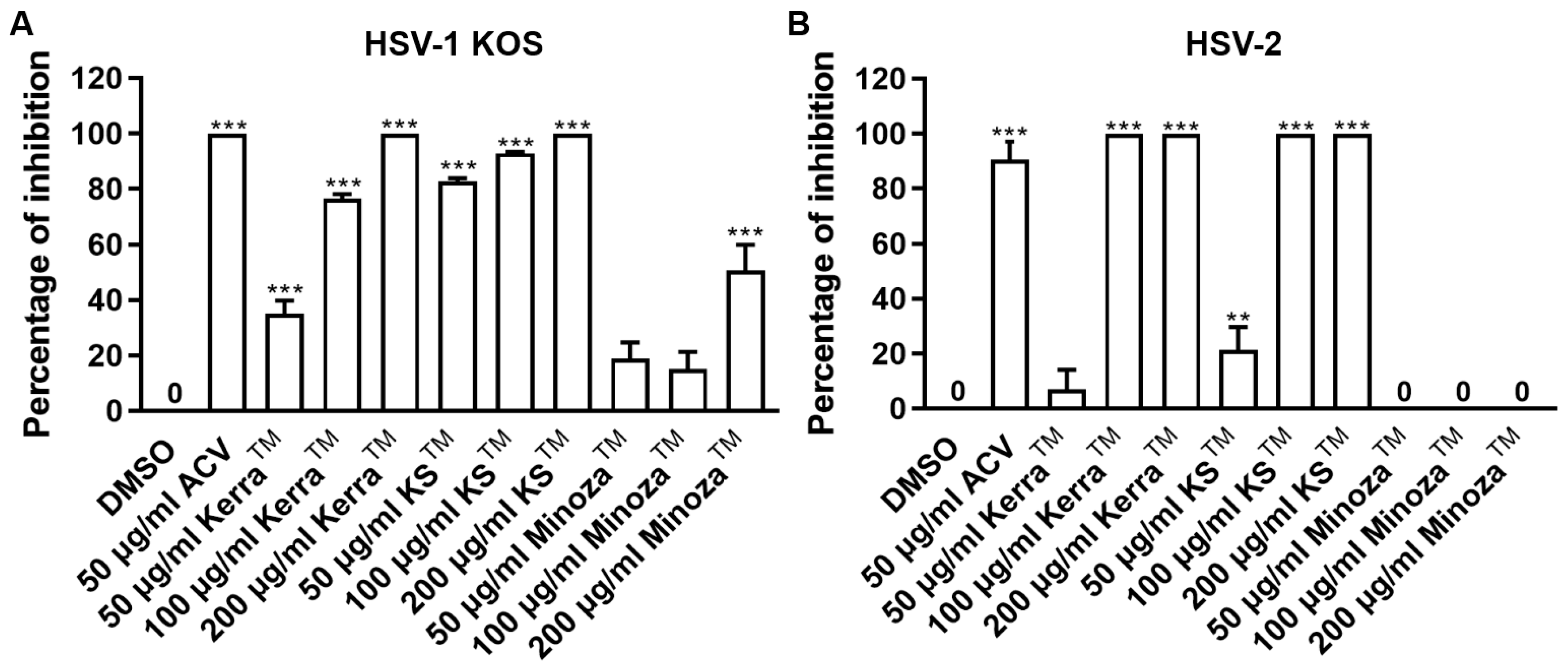
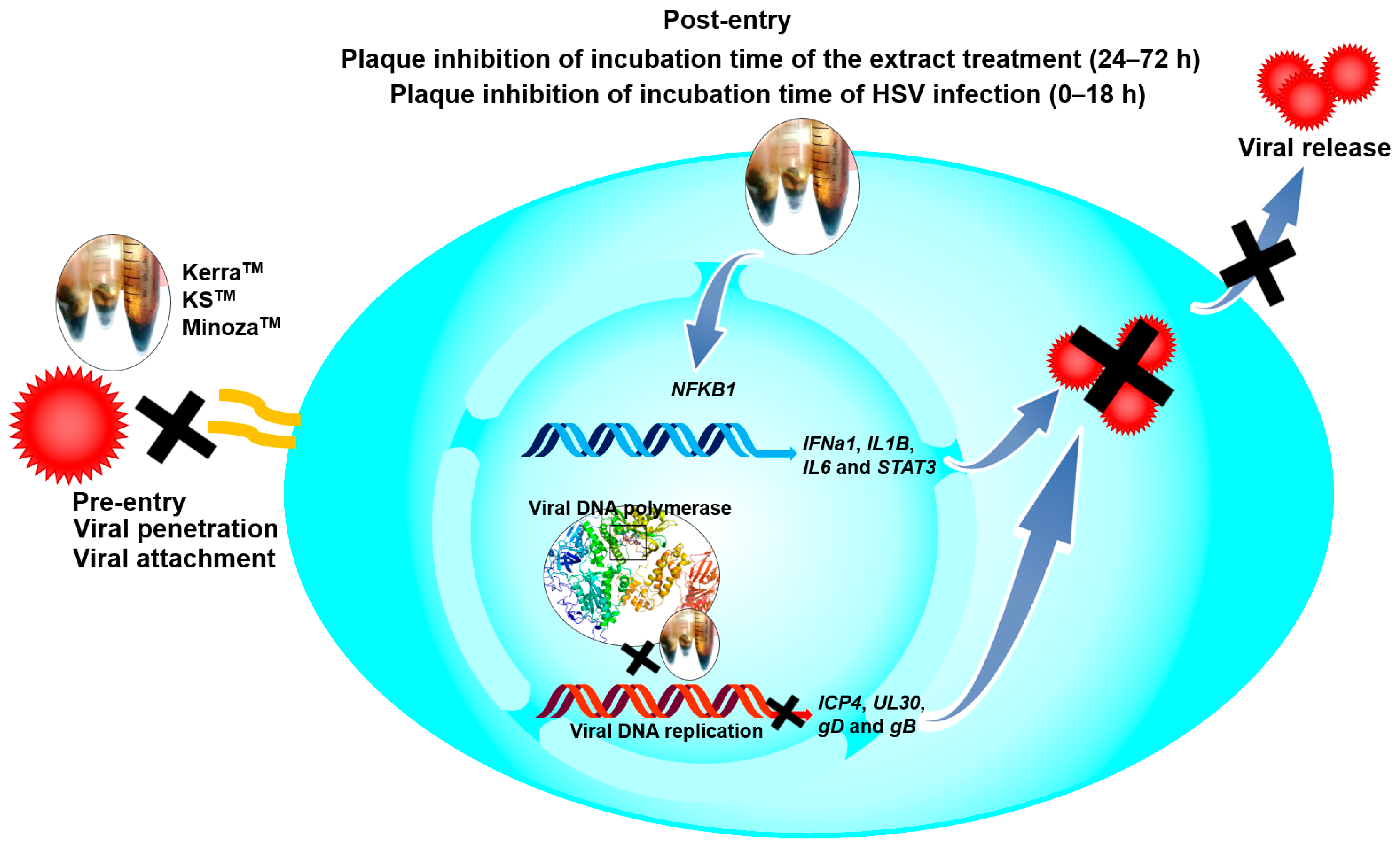
3.2. KerraTM, KSTM, and MinozaTM Significantly Inhibited the Infection with HSV-1 and HSV-2 Wild-Type Strains in the Post-Entry Step
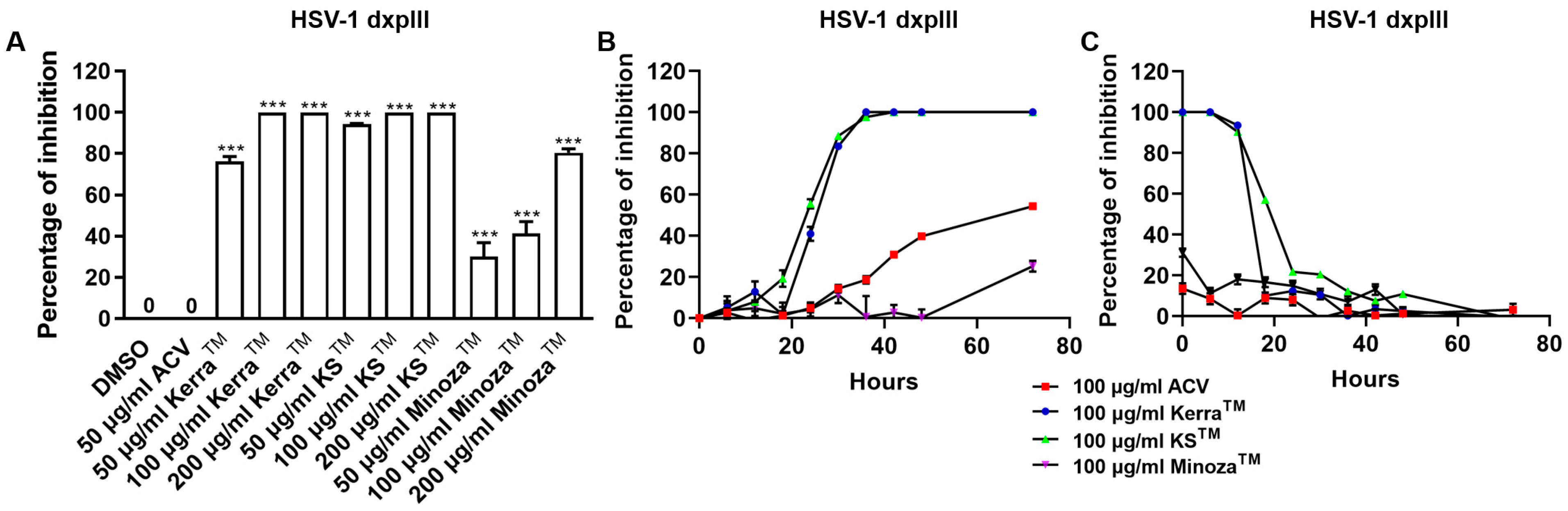
3.3. KerraTM, KSTM, and MinozaTM Significantly Inhibited HSV-1 dxpIII in the Post-Entry Step
3.4. Inhibitory Concentrations and Selective Index of KerraTM, KSTM, and MinozaTM on Anti-HSV-1 KOS, HSV-1 dxpIII, and HSV-2 in Vero Cells
3.5. KerraTM, KSTM, and MinozaTM Inhibited HSV-1 KOS, dxpIII, and HSV-2 Infection in the Pre-Entry Step
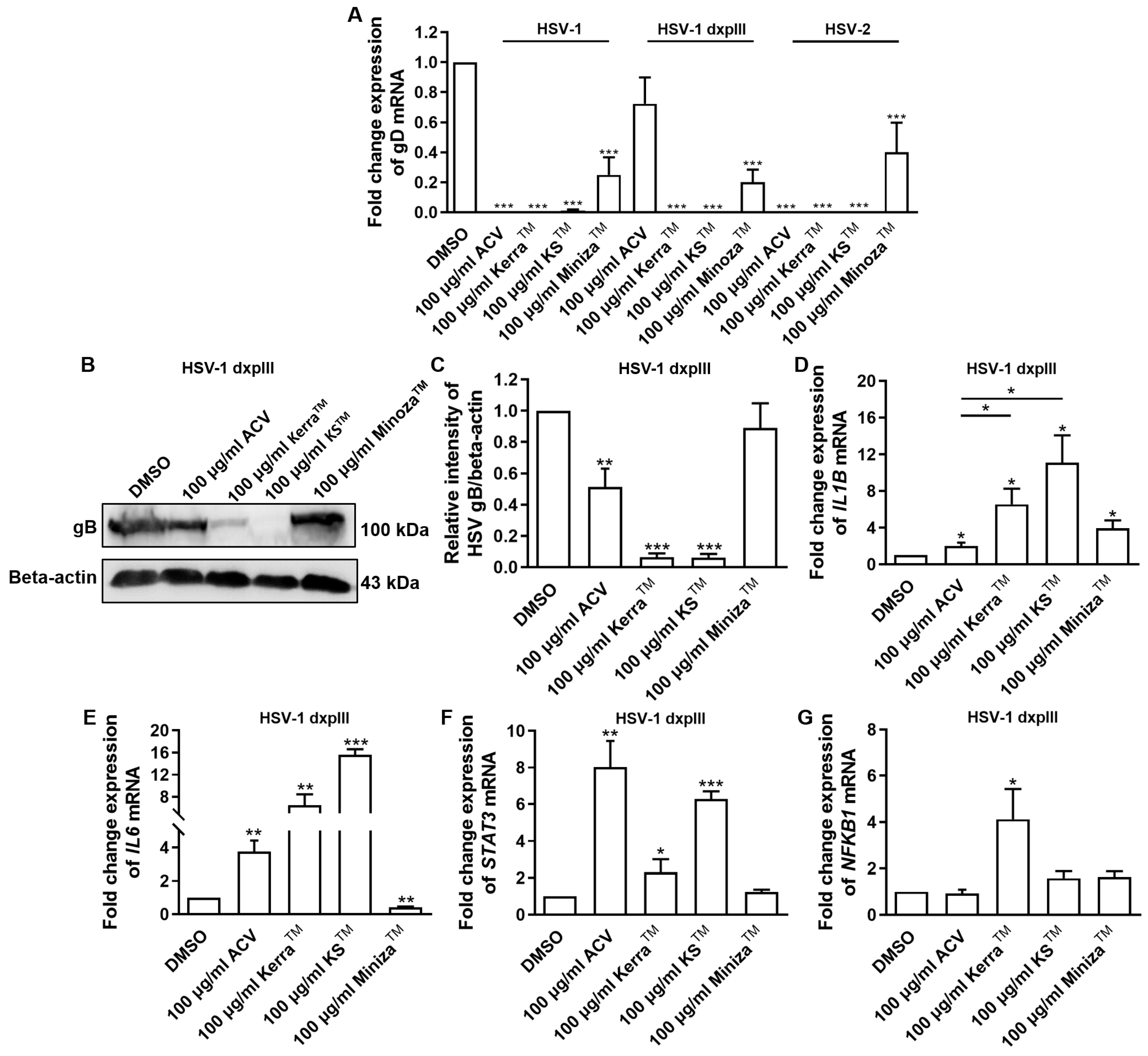
3.6. Effect of KerraTM, KSTM, and MinozaTM Treatment in HSV-Infected Vero Cells on Viral gD and Cellular IL1B, IL6, STAT3, and NFKB1 mRNA Expression
3.7. Effect of KerraTM and KSTM on Immediate Early Gene ICP4, Early Gene UL30, IL1B, and NFKB1 at Different Time Points
3.8. Phytochemical Profiling and Qualitative Metabolite Analysis
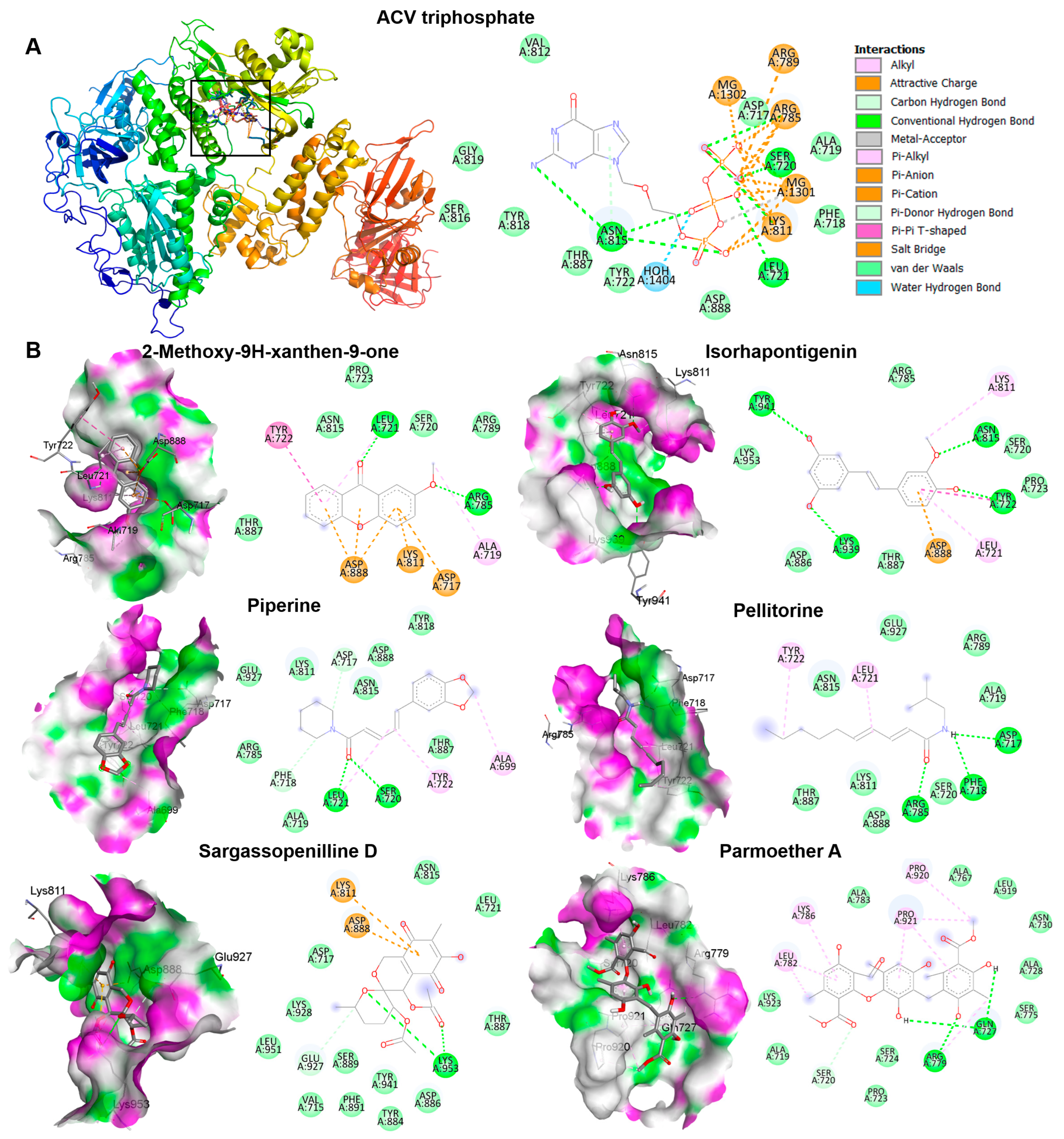
3.9. Docking Results for Candidate Most Abundant Phytochemicals in KerraTM, KSTM, and MinozaTM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tedyanto, E.H.; Susilawathi, N.M.; Indradewi, A. Herpes simplex virus encephalitis: A literature review. Rom. J. Neurol. 2021, 20, 438. [Google Scholar] [CrossRef]
- Anzivino, E.; Fioriti, D.; Mischitelli, M.; Bellizzi, A.; Barucca, V.; Chiarini, F.; Pietropaolo, V. Herpes simplex virus infection in pregnancy and in neonate: Status of art of epidemiology, diagnosis, therapy and prevention. Virol. J. 2009, 6, 40. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Viejo-Borbolla, A. Pathogenesis and virulence of herpes simplex virus. Virulence 2021, 12, 2670–2702. [Google Scholar] [CrossRef] [PubMed]
- Heath, J.R.; Dembowski, J.A. Fashionably late: Temporal regulation of HSV-1 late gene transcription. PLoS Pathog. 2022, 18, e1010536. [Google Scholar] [CrossRef]
- Bai, L.; Xu, J.; Zeng, L.; Zhang, L.; Zhou, F. A review of HSV pathogenesis, vaccine development, and advanced applications. Mol. Biomed. 2024, 5, 35. [Google Scholar] [CrossRef]
- Hassan, S.T.; Šudomová, M.; Berchová-Bímová, K.; Šmejkal, K.; Echeverría, J. Psoromic acid, a lichen-derived molecule, inhibits the replication of HSV-1 and HSV-2, and inactivates HSV-1 DNA polymerase: Shedding light on antiherpetic properties. Molecules 2019, 24, 2912. [Google Scholar] [CrossRef]
- Wald, A.; Ashley-Morrow, R. Serological testing for herpes simplex virus (HSV)–1 and HSV-2 infection. Clin. Infect. Dis. 2002, 35, S173–S182. [Google Scholar] [CrossRef]
- James, C.; Harfouche, M.; Welton, N.J.; Turner, K.M.; Abu-Raddad, L.J.; Gottlieb, S.L.; Looker, K.J. Herpes simplex virus: Global infection prevalence and incidence estimates, 2016. Bull. World Health Organ. 2020, 98, 315. [Google Scholar] [CrossRef]
- Lau, J.; Balasubramaniam, R. Oral Herpes Simplex Virus Infections. In Sexually Transmissible Oral Diseases; John Wiley & Sons: Hoboken, NJ, USA, 2023; pp. 159–167. [Google Scholar] [CrossRef]
- Piperi, E.; Papadopoulou, E.; Georgaki, M.; Dovrat, S.; Bar Illan, M.; Nikitakis, N.G.; Yarom, N. Management of oral herpes simplex virus infections: The problem of resistance. A narrative review. Oral Dis. 2024, 30, 877–894. [Google Scholar] [CrossRef]
- Avery, R.K.; Arav-Boger, R.; Marr, K.A.; Kraus, E.; Shoham, S.; Lees, L.; Trollinger, B.; Shah, P.; Ambinder, R.; Neofytos, D. Outcomes in transplant recipients treated with foscarnet for ganciclovir-resistant or refractory cytomegalovirus infection. Transplantation 2016, 100, e74–e80. [Google Scholar] [CrossRef]
- Priengprom, T.; Ekalaksananan, T.; Kongyingyoes, B.; Suebsasana, S.; Aromdee, C.; Pientong, C. Synergistic effects of acyclovir and 3, 19-isopropylideneandrographolide on herpes simplex virus wild types and drug-resistant strains. BMC Complement. Altern. Med. 2015, 15, 56. [Google Scholar] [CrossRef] [PubMed]
- Kongkaew, C.; Chaiyakunapruk, N. Efficacy of Clinacanthus nutans extracts in patients with herpes infection: Systematic review and meta-analysis of randomised clinical trials. Complement. Ther. Med. 2011, 19, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Monirifard, R. Clinical trial of efficacy of a Combined herbal remedy on Herpes Labialis. Stud. Med. Sci. 2011, 22, 315–321. [Google Scholar]
- Pipelzadeh, M.H.; Amin, M.; Khozani, A.S.; Radmanesh, M. Shallominthe active antimicrobial constituent of persian shallot in treatment of oral herpes: A double-blind randomized clinical trial. Jundishapur J. Nat. Pharm. Prod. 2014, 9, e17372. [Google Scholar] [CrossRef] [PubMed]
- Toulabi, T.; Delfan, B.; Rashidipour, M.; Yarahmadi, S.; Ravanshad, F.; Javanbakht, A.; Almasian, M. The efficacy of olive leaf extract on healing herpes simplex virus labialis: A randomized double-blind study. Explore 2022, 18, 287–292. [Google Scholar] [CrossRef]
- Hafidh, R.R.; Abdulamir, A.S.; Abu Bakar, F.; Sekawi, Z.; Jahansheri, F.; Jalilian, F.A. Novel antiviral activity of mung bean sprouts against respiratory syncytial virus and herpes simplex virus−1: An in vitro study on virally infected Vero and MRC-5 cell lines. BMC Complement. Altern. Med. 2015, 15, 179. [Google Scholar] [CrossRef]
- Boff, L.; Silva, I.; Argenta, D.; Farias, L.; Alvarenga, L.; Pádua, R.; Braga, F.; Leite, J.; Kratz, J.; Simões, C. Strychnos pseudoquina A. St. Hil.: A Brazilian medicinal plant with promising in vitro antiherpes activity. J. Appl. Microbiol. 2016, 121, 1519–1529. [Google Scholar] [CrossRef]
- Ojha, D.; Das, R.; Sobia, P.; Dwivedi, V.; Ghosh, S.; Samanta, A.; Chattopadhyay, D. Pedilanthus tithymaloides inhibits HSV infection by modulating NF-κB signaling. PLoS ONE 2015, 10, e0139338. [Google Scholar] [CrossRef]
- Kuo, Y.-C.; Kuo, Y.-H.; Lin, Y.-L.; Tsai, W.-J. Yatein from Chamaecyparis obtusa suppresses herpes simplex virus type 1 replication in HeLa cells by interruption the immediate-early gene expression. Antiviral. Res. 2006, 70, 112–120. [Google Scholar] [CrossRef]
- Choowongkomon, K.; Choengpanya, K.; Pientong, C.; Ekalaksananan, T.; Talawat, S.; Srathong, P.; Chuerduangphui, J. The Inhibitory Effect of KerraTM, KSTM, and MinozaTM on Human Papillomavirus Infection and Cervical Cancer. Medicina 2023, 59, 2169. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A. Drug-like properties and the causes of poor solubility and poor permeability. J. Pharmacol. Toxicol. Methods 2000, 44, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Veber, D.F.; Johnson, S.R.; Cheng, H.-Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef]
- Devasthanam, A.S. Mechanisms underlying the inhibition of interferon signaling by viruses. Virulence 2014, 5, 270–277. [Google Scholar] [CrossRef]
- Orzalli, M.H.; Smith, A.; Jurado, K.A.; Iwasaki, A.; Garlick, J.A.; Kagan, J.C. An antiviral branch of the IL-1 signaling pathway restricts immune-evasive virus replication. Mol. Cell 2018, 71, 825–840.e6. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, R.A.; Pesnicak, L.; Cabral, E.S.; Godleski, M.; Straus, S.E. Lack of interleukin-6 (IL-6) enhances susceptibility to infection but does not alter latency or reactivation of herpes simplex virus type 1 in IL-6 knockout mice. J. Virol. 1999, 73, 8145–8151. [Google Scholar] [CrossRef]
- Kuchipudi, S.V. The complex role of STAT3 in viral infections. J. Immunol. Res. 2015, 2015, 272359. [Google Scholar] [CrossRef]
- Seetaha, S.; Khamplong, P.; Wanaragthai, P.; Aiebchun, T.; Ratanabunyong, S.; Krobthong, S.; Yingchutrakul, Y.; Rattanasrisomporn, J.; Choowongkomon, K. KERRA, mixed medicinal plant extracts, inhibits SARS-CoV-2 targets enzymes and feline coronavirus. Covid 2022, 2, 621–632. [Google Scholar] [CrossRef]
- Trakoolsilp, B.; Vannabhum, M.; Akarasereenont, P.; Booranasubkajorn, S. Development and validation of UPLC-TQD/MS method for quantification of six bioactive constituents in Thai herbal Benjakul formula. Thai J. Pharm. Sci. 2024, 47, 3. [Google Scholar] [CrossRef]
- Saehlee, S.; Seetaha, S.; Klankaew, W.; Srathong, P.; Choowongkomon, K.; Choengpanya, K. Anti-human immunodeficiency virus-1 property of Thai herbal Extract Kerra™. Pharmaceuticals 2024, 17, 917. [Google Scholar] [CrossRef]
- Karasneh, G.A.; Shukla, D. Herpes simplex virus infects most cell types in vitro: Clues to its success. Virol. J. 2011, 8, 481. [Google Scholar] [CrossRef] [PubMed]
- Crumpacker, C.S. Mechanism of action of foscarnet against viral polymerases. Am. J. Med. 1992, 92, S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Leinbach, S.S.; Reno, J.M.; Lee, L.F.; Isbell, A.; Boezi, J.A. Mechanism of phosphonoacetate inhibition of herpesvirus-induced DNA polymerase. Biochemistry 1976, 15, 426–430. [Google Scholar] [CrossRef]
- Elion, G.B. Mechanism of action and selectivity of acyclovir. Am. J. Med. 1982, 73, 7–13. [Google Scholar] [CrossRef]
- Rabelo, V.W.-H.; Romeiro, N.C.; de Palmer Paixão, I.C.N.; Abreu, P.A. Mechanism of resistance to acyclovir in thymidine kinase mutants from Herpes simplex virus type 1: A computational approach. J. Biomol. Struct. Dyn. 2020, 38, 2116–2127. [Google Scholar] [CrossRef] [PubMed]
- Kimberlin, D.W.; Whitley, R.J. Antiviral resistance: Mechanisms, clinical significance, and future implications. J. Antimicrob. Chemother. 1996, 37, 403–421. [Google Scholar] [CrossRef]
- Godowski, P.J.; Knipe, D.M. Transcriptional control of herpesvirus gene expression: Gene functions required for positive and negative regulation. Proc. Natl. Acad. Sci. USA 1986, 83, 256–260. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal. Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Hunter, C.A.; Jones, S.A. IL-6 as a keystone cytokine in health and disease. Nat. Immunol. 2015, 16, 448–457. [Google Scholar] [CrossRef]
- Murphy, E.A.; Davis, J.M.; Brown, A.S.; Carmichael, M.D.; Ghaffar, A.; Mayer, E.P. Effect of IL-6 deficiency on susceptibility to HSV-1 respiratory infection and intrinsic macrophage antiviral resistance. J. Interferon Cytokine Res. 2008, 28, 589–596. [Google Scholar] [CrossRef]
- Maruzuru, Y.; Ichinohe, T.; Sato, R.; Miyake, K.; Okano, T.; Suzuki, T.; Koshiba, T.; Koyanagi, N.; Tsuda, S.; Watanabe, M. Herpes simplex virus 1 VP22 inhibits AIM2-dependent inflammasome activation to enable efficient viral replication. Cell Host Microbe 2018, 23, 254–265.e7. [Google Scholar] [CrossRef] [PubMed]
- Mayer-Barber, K.D.; Yan, B. Clash of the Cytokine Titans: Counter-regulation of interleukin-1 and type I interferon-mediated inflammatory responses. Cell Mol. Immunol. 2017, 14, 22–35. [Google Scholar] [CrossRef]
- Du, T.; Zhou, G.; Roizman, B. Modulation of reactivation of latent herpes simplex virus 1 in ganglionic organ cultures by p300/CBP and STAT3. Proc. Natl. Acad. Sci. USA 2013, 110, E2621–E2628. [Google Scholar] [CrossRef] [PubMed]
- Danastas, K.; Miranda-Saksena, M.; Cunningham, A.L. Herpes simplex virus type 1 interactions with the interferon system. Int. J. Mol. Sci. 2020, 21, 5150. [Google Scholar] [CrossRef] [PubMed]
- Provance, O.K.; Lewis-Wambi, J. Deciphering the role of interferon alpha signaling and microenvironment crosstalk in inflammatory breast cancer. Breast Cancer Res. 2019, 21, 59. [Google Scholar] [CrossRef] [PubMed]
- Rong, Q.; Alexander, T.; Koski, G.; Rosenthal, K.S. Multiple mechanisms for HSV-1 induction of interferon α production by peripheral blood mononuclear cells. Arch. Virol. 2003, 148, 329–344. [Google Scholar] [CrossRef]
- Sutter, J.; Bruggeman, P.J.; Wigdahl, B.; Krebs, F.C.; Miller, V. Manipulation of oxidative stress responses by non-thermal plasma to treat herpes simplex virus type 1 infection and disease. Int. J. Mol. Sci. 2023, 24, 4673. [Google Scholar] [CrossRef]
- Beristain-Bauza, S.D.C.; Hernández-Carranza, P.; Cid-Pérez, T.S.; Ávila-Sosa, R.; Ruiz-López, I.I.; Ochoa-Velasco, C.E. Antimicrobial activity of ginger (Zingiber officinale) and its application in food products. Food Rev. Int. 2019, 35, 407–426. [Google Scholar] [CrossRef]
- Vannabhum, M.; Mahajaroensiri, S.; Pattanapholkornsakul, S.; Tantiwongsekunakorn, A.; Thippayacharoentam, T.; Tripatara, P.; Akarasereenont, P. Metabolomics of personalized body elements in Thai traditional medicine response to herbal medicine for body elements balancing in healthy volunteers. Evid. Based Complement. Alternat. Med. 2023, 2023, 6684263. [Google Scholar] [CrossRef]
- Che Sulaiman, I.S.; Mohamad, A.; Ahmed, O.H. Murdannia loriformis: A review of ethnomedicinal uses, phytochemistry, pharmacology, contemporary application, and toxicology. Evid. Based Complement. Alternat. Med. 2021, 2021, 9976202. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | (mg/mL) |
|---|---|
| ACV | 7.545 ± 0.133 |
| KerraTM | 2.846 ± 0.025 |
| KSTM | 2.349 ± 0.026 |
| MinozaTM | 11.670 ± 1.804 |
| Extract | HSV-1 KOS | HSV-1 dxpIII | HSV-2 | |||
|---|---|---|---|---|---|---|
| (μg/mL) | SI (Unitless) | (μg/mL) | SI (Unitless) | (μg/mL) | SI (Unitless) | |
| ACV | 0.283 ± 0.000 | 26,554 | >2000 | <3.757 | 0.360 ± 0.000 | 20,875 |
| KerraTM | 66.543 ± 4.759 | 42.769 | 41.394 ± 0.471 | 68.753 | 72.519 ± 2.480 | 39.244 |
| KSTM | 25.476 ± 3.430 | 92.204 | 38.257 ± 0.04 | 61.400 | 67.065 ± 3.525 | 35.025 |
| MinozaTM | 187.807 ± 39.869 | 62.138 | 73.086 ± 6.682 | 159.669 | >200 | <58.35 |
| Extract | Phytochemicals | |||||
|---|---|---|---|---|---|---|
| Name/Formula | Method | Ref. | ||||
| KerraTM | 2-Methoxy-9H-xanthen-9-one, Isorhapontigenin, Betaine, C20H28O4, trans-Anethole, Eicosatetraynoic acid, NP-020078, NP-003294, C20H30O5, and N1-(3-chlorophenyl)-2-[2-(trifluoromethyl)-4 | LC-MS/MS | [29] | |||
| KSTM | Piperine, pellitorine, 6-gingerol, piperlonguminine, plumbagin, and 6-shogaol | UPLC-TQD/MS | [30] | |||
| MinozaTM | Name | Formula | m/z | RT (min) | Molecular Weight (Da) | Area |
| Sargassopenilline D | 395.133 | 3.56 | 394.12576 | 1.12 × 1010 | ||
| Parmoether A | 541.16974 | 5.053 | 540.16249 | 7.94 × 109 | ||
| 4R-aminopentanoic acid | 118.08586 | 0.379 | 117.07859 | 4.57 × 109 | ||
| (R)-2,4,5-Trimethoxydalbergiquinol | 285.14847 | 6.366 | 284.14119 | 1.11 × 109 | ||
| Pseudosindorin | 273.07559 | 4.477 | 272.06832 | 8.57 × 108 | ||
| 5,3′-Dihydroxy-6,7,4′,5′-tetramethoxyflavanone | 377.12302 | 4.436 | 376.11574 | 8.30 × 108 | ||
| iso-Debromo-laurinterol | 217.15854 | 6.349 | 216.15128 | 8.22 × 108 | ||
| 2-(5-((2Z,5Z,8Z,11Z)-tetradeca-2,5,8,11-tetraen-1-yl)furan-2-yl)-ethanoic acid | 315.19531 | 6.091 | 314.18804 | 6.55 × 108 | ||
| Hypnosin | 182.05577 | 0.357 | 181.04849 | 5.60 × 108 | ||
| 4-Hydroxy-2′,4′-dimethoxydihydrochalcone | 287.12781 | 6.858 | 286.12055 | 5.06 × 108 | ||
| Phytochemicals | Binding Energy (kcal/mol) | MW g/mol (<500) | LogP (<5) | HBA (<10) | HBD (<5) | RB (<10) | TPA Å2 <140 | Water Solubility | Drug-Likeness |
|---|---|---|---|---|---|---|---|---|---|
| 2-Methoxy-9H-xanthen-9-one | −7.0 | 226.23 | 2.85 | 3 | 0 | 1 | 39.44 | Moderately soluble | Yes |
| Isorhapontigenin | −6.7 | 258.27 | 2.63 | 4 | 3 | 3 | 69.92 | Soluble | Yes [31] |
| Piperine | −7.0 | 285.34 | 3.03 | 3 | 0 | 4 | 38.77 | Soluble | Yes |
| Pellitorine | −5.4 | 223.35 | 3.64 | 1 | 1 | 9 | 29.10 | Soluble | Yes |
| Sargassopenilline D | −6.9 | 394.37 | 0.95 | 9 | 1 | 4 | 125.43 | Soluble | Yes |
| Parmoether A | −8.0 | 540.52 | 3.83 | 11 | 5 | 9 | 140.34 | Poorly soluble | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loymunkong, C.; Choowongkomon, K.; Heawchaiyaphum, C.; Chatchawankanpanich, N.; Pientong, C.; Ekalaksananan, T.; Chuerduangphui, J. Anti-Herpes Simplex Virus (Wild-Type and Drug-Resistant) Properties of Herbal KerraTM, KSTM, and MinozaTM. Viruses 2025, 17, 889. https://doi.org/10.3390/v17070889
Loymunkong C, Choowongkomon K, Heawchaiyaphum C, Chatchawankanpanich N, Pientong C, Ekalaksananan T, Chuerduangphui J. Anti-Herpes Simplex Virus (Wild-Type and Drug-Resistant) Properties of Herbal KerraTM, KSTM, and MinozaTM. Viruses. 2025; 17(7):889. https://doi.org/10.3390/v17070889
Chicago/Turabian StyleLoymunkong, Chaleampol, Kiattawee Choowongkomon, Chukkris Heawchaiyaphum, Nutchanat Chatchawankanpanich, Chamsai Pientong, Tipaya Ekalaksananan, and Jureeporn Chuerduangphui. 2025. "Anti-Herpes Simplex Virus (Wild-Type and Drug-Resistant) Properties of Herbal KerraTM, KSTM, and MinozaTM" Viruses 17, no. 7: 889. https://doi.org/10.3390/v17070889
APA StyleLoymunkong, C., Choowongkomon, K., Heawchaiyaphum, C., Chatchawankanpanich, N., Pientong, C., Ekalaksananan, T., & Chuerduangphui, J. (2025). Anti-Herpes Simplex Virus (Wild-Type and Drug-Resistant) Properties of Herbal KerraTM, KSTM, and MinozaTM. Viruses, 17(7), 889. https://doi.org/10.3390/v17070889