
Modulation of Plant Interactions with Whitefly and Whitefly-Borne Viruses by Salicylic Acid Signaling Pathway: A Review




Abstract
1. Perspectives and Overview
2. A Brief Introduction to the SA Signaling Pathway
2.1. SA Biosynthesis
2.2. Signal Transduction in the SA Signaling Pathway
3. The Role of the SA Signaling Pathway in Plant–Whitefly Interactions
3.1. The Response of the Plant SA Signaling Pathway to Whitefly Infestation
3.2. Effects of SA Signaling Pathway on Whitefly Performance and Preference
4. Interactions of Plants with Whitefly-Borne Viruses
4.1. The Response of the SA Signaling Pathway to Whitefly-Borne Viruses
4.2. Manipulation of the SA Signaling Pathways by Viral Proteins
5. Plant–Whitefly–Virus Tripartite Interactions
5.1. Effects of Virus-Induced Plant SA Signaling Pathway on Whitefly Performance
5.2. Effects of Whitefly-Induced Plant SA Signaling Pathway on Virus Infection
6. Prospects for Future Research
6.1. The Evolution of Whitefly Interaction with Plant SA Signaling Pathway
6.2. Whitefly and Plant Factors Mediating the Induction of the SA Signaling Pathway
6.3. The Mechanisms Underlying the Induction and Mitigation of Plant SA Signaling Pathways by Whitefly-Borne Viruses
6.4. The Role of SA Signaling Pathways in Plant–Whitefly–Virus Tripartite Interactions and Beyond
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Lei, T.; Wang, X.W.; Cameron, S.; Navas-Castillo, J.; Liu, Y.Q.; Maruthi, M.N.; Omongo, C.A.; Delatte, H.; Lee, Y.K.; et al. A comprehensive framework for the delimitation of species within the Bemisia tabaci cryptic complex, a global pest-species group. Insect Sci. 2025, 32, 321–342. [Google Scholar] [CrossRef]
- Hu, J.; DeBarro, P.J.; Zhao, H.; Wang, J.; Nardi, F.; Liu, S.S. An extensive field survey combined with a phylogenetic analysis reveals rapid and widespread invasion of two alien whiteflies in China. PLoS ONE 2011, 6, e16061. [Google Scholar] [CrossRef]
- Malka, O.; Santos-Garcia, D.; Feldmesser, E.; Sharon, E.; Krause-Sakate, R.; Delatte, H.; van Brunschot, S.; Patel, M.; Visendi, P.; Mugerwa, H.; et al. Species-complex diversification and host-plant associations in Bemisia tabaci: A plant defence, detoxification perspective revealed by RNA-Seq analyses. Mol. Ecol. 2018, 27, 4241–4256. [Google Scholar] [CrossRef]
- Oliveira, M.R.V.; Henneberry, T.J.; Anderson, P. History, current status, and collaborative research projects for Bemisia tabaci. Crop Prot. 2001, 20, 709–723. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Navas-Castillo, J. Tomato chlorosis virus, a promiscuous virus with multiple host plants and whitefly vectors. Ann. Appl. Biol. 2023, 182, 29–36. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Navas-Castillo, J. Begomoviruses: What is the secret(s) of their success? Trends Plant Sci. 2023, 28, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Fiallo-Olivé, E.; Pan, L.L.; Liu, S.S.; Navas-Castillo, J. Transmission of begomoviruses and other whitefly-borne viruses: Dependence on the vector species. Phytopathology 2020, 110, 10–17. [Google Scholar] [CrossRef]
- Wang, X.W.; Blanc, S. Insect transmission of plant single-stranded DNA viruses. Annu. Rev. Entomol. 2021, 66, 389–405. [Google Scholar] [CrossRef]
- Mithöfer, A.; Boland, W. Plant defense against herbivores: Chemical aspects. Annu. Rev. Plant Biol. 2012, 63, 431–450. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhang, L.L.; Zhang, X.; Wu, X.J.; Fang, R.X. Plant defense networks against insect-borne pathogens. Trends Plant Sci. 2021, 26, 272–287. [Google Scholar] [CrossRef]
- Ge, L.H.; Zhou, X.P.; Li, F.F. Plant-virus arms race beyond RNA interference. Trends Plant Sci. 2024, 29, 16–19. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- Chivasa, S.; Murphy, A.M.; Naylor, M.; Carr, J.P. Salicylic acid interferes with tobacco mosaic virus replication via a novel salicylhydroxamic acid-sensitive mechanism. Plant Cell 1997, 9, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Wildermuth, M.C.; Dewdney, J.; Wu, G.; Ausubel, F.M. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 2001, 414, 562–565. [Google Scholar] [CrossRef]
- Peng, Y.J.; Yang, J.F.; Li, X.; Zhang, Y.L. Salicylic acid: Biosynthesis and signaling. Annu. Rev. Plant Biol. 2021, 72, 761–791. [Google Scholar] [CrossRef]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.F.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Chen, Z.X. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Yalpani, N.; León, J.; Lawton, M.A.; Raskin, I. Pathway of salicylic acid biosynthesis in healthy and virus-inoculated tobacco. Plant Physiol. 1993, 103, 315–321. [Google Scholar] [CrossRef]
- Shine, M.B.; Yang, J.W.; El-Habbak, M.; Nagyabhyru, P.; Fu, D.Q.; Navarre, D.; Ghabrial, S.; Kachroo, P.; Kachroo, A. Cooperative functioning between phenylalanine ammonia lyase and isochorismate synthase activities contributes to salicylic acid biosynthesis in soybean. New Phytol. 2016, 212, 627–636. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Yang, G.Q.; Zhang, D.D.; Li, G.X.; Qiu, J.L.; Wu, J. Isochorismate synthase is required for phylloquinone, but not salicylic acid biosynthesis in rice. aBIOTECH 2024, 5, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Garcion, C.; Lohmann, A.; Lamodière, E.; Catinot, J.; Buchala, A.; Doermann, P.; Métraux, J.P. Characterization and biological function of the ISOCHORISMATE SYNTHASE2 gene of Arabidopsis. Plant Physiol. 2008, 147, 1279–1287. [Google Scholar] [CrossRef]
- Rekhter, D.; Lüdke, D.; Ding, Y.; Feussner, K.; Zienkiewicz, K.; Lipka, V.; Wiermer, M.; Zhang, Y.L.; Feussner, I. Isochorismate-derived biosynthesis of the plant stress hormone salicylic acid. Science 2019, 365, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Spence, M.P.; Bobokalonova, A.; Carballo, V.; Glinkerman, C.M.; Pluskal, T.; Shen, A.; Weng, J.K. PBS3 and EPS1 complete salicylic acid biosynthesis from isochorismate in Arabidopsis. Mol. Plant 2019, 12, 1577–1586. [Google Scholar] [CrossRef]
- Torrens-Spence, M.P.; Matos, J.O.; Li, T.J.; Kastner, D.W.; Kim, C.Y.; Wang, Z.Q.; Glinkerman, C.M.; Sherk, J.; Kulik, H.J.; Wang, Y.; et al. Mechanistic basis for the emergence of EPS1 as a catalyst in salicylic acid biosynthesis of Brassicaceae. Nat. Commun. 2024, 15, 10356. [Google Scholar] [CrossRef]
- Maeda, H.; Shasany, A.K.; Schnepp, J.; Orlova, I.; Taguchi, G.; Cooper, B.R.; Rhodes, D.; Pichersky, E.; Dudareva, N. RNAi suppression of Arogenate Dehydratase1 reveals that phenylalanine is synthesized predominantly via the arogenate pathway in petunia petals. Plant Cell 2010, 22, 832–849. [Google Scholar] [CrossRef]
- Qian, Y.C.; Lynch, J.H.; Guo, L.Y.; Yang, Y.; Schmelz, E.A.; Pichersky, E.; Maeda, H. Completion of the cytosolic post-chorismate phenylalanine biosynthetic pathway in plants. Nat. Commun. 2019, 10, 15. [Google Scholar] [CrossRef]
- Bussell, J.D.; Reichelt, M.; Wiszniewski, A.A.; Gershenzon, J.; Pichersky, E.; Maeda, H. Peroxisomal ATP-binding cassette transporter COMATOSE and the multifunctional protein abnormal INFLORESCENCE MERISTEM are required for the production of benzoylated metabolites in Arabidopsis seeds. Plant Physiol. 2014, 164, 48–54. [Google Scholar] [CrossRef] [PubMed]
- León, J.; Yalpani, N.; Raskin, I.; Lawton, M.A. Induction of benzoic acid 2-hydroxylase in virus-inoculated tobacco. Plant Physiol. 1993, 103, 323–328. [Google Scholar] [CrossRef]
- León, J.; Shulaev, V.; Yalpani, N.; Lawton, M.A.; Raskin, I. Benzoic acid 2-hydroxylase, a soluble oxygenase from tobacco, catalyzes salicylic acid biosynthesis. Proc. Natl. Acad. Sci. USA 1995, 92, 10413–10417. [Google Scholar] [CrossRef]
- Zavaliev, R.; Dong, X. NPR1, a key immune regulator for plant survival under biotic and abiotic stresses. Mol. Cell 2024, 84, 131–141. [Google Scholar] [CrossRef]
- Cao, H.; Glazebrook, J.; Clarke, J.D.; Volko, S.M.; Dong, X. The Arabidopsis NPR1 gene that controls systemic acquired resistance encodes a novel protein containing ankyrin repeats. Cell 1997, 88, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Mou, Z.; Fan, W.; Dong, X. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 2003, 113, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Tada, Y.; Spoel, S.H.; Pajerowska-Mukhtar, K.; Mou, Z.; Song, J.; Durrant, W.E.; Wang, D.; Dong, X.; Parker, J.E. Plant immunity requires conformational changes of NPR1 via S-nitrosylation and thioredoxins. Science 2008, 321, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.H.; Mou, Z.; Tada, Y.; Trujillo, M.; Chen, Z.; Parker, J.E.; Dong, X. Proteasome-mediated turnover of the transcription coactivator NPR1 plays dual roles in regulating plant immunity. Cell 2009, 137, 860–872. [Google Scholar] [CrossRef]
- Saleh, A.; Withers, J.; Mohan, R.; Durrant, W.E.; Dong, X.; Després, C.; Parker, J.E.; Jones, J.D. Posttranslational modifications of the master transcriptional regulator NPR1 enable dynamic but tight control of plant immune responses. Cell Host Microbe 2015, 18, 169–182. [Google Scholar] [CrossRef]
- Skelly, M.J.; Furniss, J.J.; Grey, H.; Sadanandom, A.; Spoel, S.H. Dynamic ubiquitination determines transcriptional activity of the plant immune coactivator NPR1. eLife 2019, 8, e47005. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Fan, W.; Kinkema, M.; Li, X.; Dong, X. Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene. Proc. Natl. Acad. Sci. USA 1999, 96, 6523–6528. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Tessaro, M.J.; Lassner, M.; Li, X. Knockout analysis of Arabidopsis transcription factors TGA2, TGA5, and TGA6 reveals their redundant and essential roles in systemic acquired resistance. Plant Cell 2003, 15, 2647–2653. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, C.; Zhang, Y.; Li, X.; Dong, X. The Arabidopsis Mediator complex subunit16 positively regulates salicylate-mediated systemic acquired resistance and jasmonate/ethylene-induced defense pathways. Plant Cell 2012, 24, 4294–4309. [Google Scholar] [CrossRef]
- Zhang, X.; Yao, J.; Zhang, Y.; Li, X.; Dong, X. The Arabidopsis Mediator complex subunits MED14/SWP and MED16/SFR6/IEN1 differentially regulate defense gene expression in plant immune responses. Plant J. 2013, 75, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Choi, S.M.; Kang, M.J.; Yun, S.H.; Kwon, D.J.; Noh, Y.S.; Noh, B. Salicylic acid-induced transcriptional reprogramming by the HAC–NPR1–TGA histone acetyltransferase complex in Arabidopsis. Nucleic Acids Res. 2018, 46, 11712–11725. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, M.; Qi, G.; Zhao, M.; Liu, L.; Liu, L.; Palmer, I.A.; Gassmann, W.; Fu, Z.Q. Two interacting transcriptional coactivators cooperatively control plant immune responses. Sci. Adv. 2021, 7, eabl7173. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Liao, H.; Wang, X.; Dong, X. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef]
- Ding, Y.; Sun, T.; Ao, K.; Wang, D.; Zhang, Y.; Li, Y.; Dong, X. Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of plant immunity. Cell 2018, 173, 1454–1467. [Google Scholar] [CrossRef]
- Morin, S.; Atkinson, P.W.; Walling, L.L. Whitefly-plant interactions: An integrated molecular perspective. Annu. Rev. Entomol. 2024, 69, 503–525. [Google Scholar] [CrossRef]
- Zarate, S.I.; Kempema, L.A.; Walling, L.L. Silverleaf whitefly induces salicylic acid defenses and suppresses effectual jasmonic acid defenses. Plant Physiol. 2007, 143, 866–875. [Google Scholar] [CrossRef]
- Zhang, P.J.; Zheng, S.J.; van Loon, J.J.; Boland, W.; David, A.; Mumm, R.; Dicke, M. Whiteflies interfere with indirect plant defense against spider mites in Lima bean. Proc. Natl. Acad. Sci. USA 2009, 106, 21202–21207. [Google Scholar] [CrossRef]
- Puthoff, D.P.; Holzer, F.M.; Perring, T.M.; Walling, L.L. Tomato pathogenesis-related protein genes are expressed in response to Trialeurodes vaporariorum and Bemisia tabaci biotype B feeding. J. Chem. Ecol. 2010, 36, 1271–1285. [Google Scholar] [CrossRef]
- Cui, H.Y.; Sun, Y.C.; Zhao, Z.H.; Zhang, Y.J. The combined effect of elevated O3 levels and TYLCV infection increases the fitness of Bemisia tabaci Mediterranean on tomato plants. Environ. Entomol. 2019, 48, 1425–1433. [Google Scholar] [CrossRef]
- Zhang, P.J.; Xu, C.X.; Zhang, J.M.; Lu, Y.B.; Wei, J.N.; Liu, Y.Q.; David, A.; Boland, W.; Turlings, T.C.J. Phloem-feeding whiteflies can fool their host plants, but not their parasitoids. Funct. Ecol. 2013, 27, 1304–1312. [Google Scholar] [CrossRef]
- Zhang, P.J.; Li, W.D.; Huang, F.; Zhang, J.M.; Xu, F.C.; Lu, Y.B. Feeding by whiteflies suppresses downstream jasmonic acid signaling by eliciting salicylic acid signaling. J. Chem. Ecol. 2013, 39, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xue, M.; Zhao, H.P. Species-specific effects on salicylic acid content and subsequent Myzus persicae (Sulzer) performance by three phloem-sucking insects infesting Nicotiana tabacum L. Arthropod-Plant Interact. 2015, 9, 383–391. [Google Scholar] [CrossRef]
- Zhao, H.P.; Zhang, X.; Xue, M.; Zhang, X. Feeding of whitefly on tobacco decreases aphid performance via increased salicylate signaling. PLoS ONE 2015, 10, e0138584. [Google Scholar] [CrossRef]
- Vieira, S.S.; Lourenço, A.L.; da Graça, J.P.; Janegitz, T.; Salvador, M.C.; de Oliveira, M.C.N.; Hoffmann-Campo, C.B. Biological aspects of Bemisia tabaci biotype B and the chemical causes of resistance in soybean genotypes. Arthropod-Plant Interact. 2016, 10, 525–534. [Google Scholar] [CrossRef]
- Li, P.; Shu, Y.N.; Fu, S.; Liu, Y.Q.; Zhou, X.P.; Liu, S.S.; Wang, X.W. Vector and nonvector insect feeding reduces subsequent plant susceptibility to virus transmission. New Phytol. 2017, 215, 699–710. [Google Scholar] [CrossRef]
- Xu, H.X.; Qian, L.X.; Wang, X.W.; Shao, R.X.; Hong, Y.; Liu, S.S.; Wang, X.W. A salivary effector enables whitefly to feed on host plants by eliciting salicylic acid-signaling pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 490–495. [Google Scholar] [CrossRef]
- Zhang, P.J.; Wei, J.N.; Zhao, C.; Zhang, Y.F.; Li, C.Y.; Liu, S.S.; Dicke, M.; Yu, X.P.; Turlings, T.C.J. Airborne host-plant manipulation by whiteflies via an inducible blend of plant volatiles. Proc. Natl. Acad. Sci. USA 2019, 116, 7387–7396. [Google Scholar] [CrossRef]
- Silva, D.B.; Jiménez, A.; Urbaneja, A.; Pérez-Hedo, M.; Bento, J.M. Changes in plant responses induced by an arthropod influence the colonization behavior of a subsequent herbivore. Pest. Manag. Sci. 2021, 77, 4168–4180. [Google Scholar] [CrossRef]
- Li, Y.; Qu, C.; Yan, X.Y.; Sun, X.; Yin, Z.Y.; Zhao, H.P. Effect of feeding stage and density of whiteflies on subsequent aphid performance on tobacco plants. Agronomy 2022, 12, 1025. [Google Scholar] [CrossRef]
- Hu, J.; Sun, G.; Yang, Y.; Jiao, X.; Chen, Z.; Zhang, Y. Pepper previously infested by MED facilitates settling and oviposition by MEAM1 of the Bemisia tabaci species complex. J. Pest. Sci. 2023, 96, 1019–1034. [Google Scholar] [CrossRef]
- Liu, Y.X.; Han, W.H.; Wang, J.X.; Zhang, Y.L.; Li, Y.; Li, M.; Liu, S.S.; Wang, X.W. Differential induction of JA/SA determines plant defense against successive leaf-chewing and phloem-feeding insects. J. Pest. Sci. 2025, 98, 1085–1100. [Google Scholar] [CrossRef]
- Wu, H.; Han, W.H.; Liang, K.L.; Wang, J.X.; Zhang, F.B.; Ji, S.X.; Wang, X.W. Using salicylic acid-responsive promoters to drive the expression of jasmonic acid-regulated genes enhances plant resistance to whiteflies. Pest Manag Sci. 2024. [Google Scholar] [CrossRef]
- Song, H.D.; Zhang, F.B.; Ji, S.X.; Zhang, Y.L.; Li, Y.; Liu, L.; Liu, S.S.; Wang, X.W. The SA-WRKY70-PR-Callose axis mediates plant defense against whitefly eggs. Int. J. Mol. Sci. 2024, 25, 12076. [Google Scholar] [CrossRef]
- Zhang, J.R.; Liu, Y.M.; Li, D.; Wu, Y.J.; Zhao, S.X.; Wang, X.W.; Liu, S.S.; Walling, L.L.; Pan, L.L. Viral proteins resolve the virus-vector conundrum during hemipteran-mediated transmission by subverting salicylic acid signaling pathway. Nat. Commun. 2024, 15, 9448. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.B.; Pan, H.P.; Xie, W.; Jiao, X.G.; Fang, Y.; Chen, G.; Yang, X.; Wu, Q.J.; Wang, S.L.; Zhang, Y.J. Three-way interactions between the tomato plant, tomato yellow leaf curl virus, and Bemisia tabaci (Hemiptera: Aleyrodidae) facilitate virus spread. J. Econ. Entomol. 2014, 107, 920–926. [Google Scholar] [CrossRef]
- Rodríguez-Álvarez, C.I.; López-Climent, M.F.; Gómez-Cadenas, A.; Kaloshian, I.; Nombela, G. Salicylic acid is required for Mi-1-mediated resistance of tomato to whitefly Bemisia tabaci, but not for basal defense to this insect pest. Bull. Entomol. Res. 2015, 105, 574–582. [Google Scholar] [CrossRef]
- Yang, J.W.; Yi, H.S.; Kim, H.; Lee, B.; Lee, S.; Ghim, S.Y.; Ryu, C.M. Whitefly infestation of pepper plants elicits defence responses against bacterial pathogens in leaves and roots and changes the below-ground microflora. J. Ecol. 2011, 99, 46–56. [Google Scholar] [CrossRef]
- Park, Y.S.; Ryu, C.M. Understanding cross-communication between aboveground and belowground tissues via transcriptome analysis of a sucking insect whitefly-infested pepper plants. Biochem. Biophys. Res. Commun. 2014, 443, 272–277. [Google Scholar] [CrossRef]
- Shi, X.B.; Chen, G.; Tian, L.X.; Zhang, Y.L.; Li, Y.; Liu, L.; Liu, S.S.; Wang, X.W. The salicylic acid-mediated release of plant volatiles affects the host choice of Bemisia tabaci. Int. J. Mol. Sci. 2016, 17, 1048. [Google Scholar] [CrossRef]
- Jafarbeigi, F.; Samih, M.A.; Alaei, H.; Shirani, H. Induced tomato resistance against Bemisia tabaci triggered by salicylic acid, β-aminobutyric acid, and Trichoderma. Neotrop. Entomol. 2020, 49, 456–467. [Google Scholar] [CrossRef]
- Ueda, H.; Kugimiya, S.; Tabata, J.; Noh, Y.S.; Noh, B. Accumulation of salicylic acid in tomato plant under biological stress affects oviposition preference of Bemisia tabaci. J. Plant Interact. 2018, 14, 73–78. [Google Scholar] [CrossRef]
- Ascencio-Ibáñez, J.T.; Sozzani, R.; Lee, T.J.; Chu, T.M.; Wolfinger, R.D.; Cella, R.; Hanley-Bowdoin, L. Global analysis of Arabidopsis gene expression uncovers a complex array of changes impacting pathogen response and cell cycle during geminivirus infection. Plant Physiol. 2018, 148, 436–454. [Google Scholar] [CrossRef]
- Luna-Rivero, M.S.; Hernández-Zepeda, C.; Villanueva-Alonzo, H.; Minero-García, Y.; Castell-González, S.E.; Moreno-Valenzuela, O.A. Expression of genes involved in the salicylic acid pathway in type h1 thioredoxin transiently silenced pepper plants during a begomovirus compatible interaction. Mol. Genet. Genom. 2016, 291, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Kundu, A.; Patel, A.; Paul, S.; Pal, A. Transcript dynamics at early stages of molecular interactions of MYMIV with resistant and susceptible genotypes of the leguminous host, Vigna mungo. PLoS ONE 2015, 10, e0124687. [Google Scholar] [CrossRef] [PubMed]
- Jeevalatha, A.; Siddappa, S.; Kumar, R.; Tiwari, R.K.; Lal, M.K.; Sharma, S.; Chakrabarti, S.K.; Singh, B.P. RNA-seq analysis reveals an early defense response to tomato leaf curl New Delhi virus in potato cultivar Kufri Bahar. Funct. Integr. Genom. 2023, 23, 215. [Google Scholar] [CrossRef] [PubMed]
- Roshan, P.; Kulshreshtha, A.; Kumar, S.; Sharma, R.; Kumar, V.; Kumar, M.; Kumar, S.; Kumar, A.; Ranjan, A.; Sharma, Y.K.; et al. AV2 protein of Tomato Leaf Curl Palampur Virus promotes systemic necrosis in Nicotiana benthamiana and interacts with host catalase2. Sci. Rep. 2018, 8, 1273. [Google Scholar] [CrossRef]
- Miozzi, L.; Napoli, C.; Sardo, L.; Accotto, G.P. Transcriptomics of the interaction between the monopartite phloem-limited geminivirus Tomato Yellow Leaf Curl Sardinia Virus and Solanum lycopersicum highlights a role for plant hormones, autophagy, and plant immune system fine tuning during infection. PLoS ONE 2014, 9, e89951. [Google Scholar] [CrossRef]
- Sade, D.; Sade, N.; Shriki, O.; Lerner, S.; Gebremedhin, A.; Karavani, A.; Brotman, Y.; Osorio, S.; Fernie, A.R.; Willmitzer, L.; et al. Water balance, hormone homeostasis, and sugar signaling are all involved in tomato resistance to Tomato yellow leaf curl virus. Plant Physiol. 2014, 165, 1684–1697. [Google Scholar] [CrossRef]
- Su, Q.; Preisser, E.L.; Zhou, X.M.; Li, Y.; Wang, X.W. Manipulation of host quality and defense by a plant virus improves performance of whitefly vectors. J. Econ. Entomol. 2015, 108, 11–19. [Google Scholar] [CrossRef]
- Zingariello, E.; Larocca, M.; Rossano, R.; Crescenzi, A.; Fanigliulo, A.; Viggiano, A. Comparative analysis of induction of Pr-1 protein between acibenzolar-S-methyl treatment and virus infection by TSWV and TYLCV in Solanum lycopersicum L. Acta Hortic. 2015, 1069, 265–270. [Google Scholar] [CrossRef]
- Cui, H.; Sun, Y.; Chen, F.; Zhang, Y.; Ge, F. Elevated O3 and TYLCV infection reduce the suitability of tomato as a host for the whitefly Bemisia tabaci. Int. J. Mol. Sci. 2016, 17, 1964. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, Z.; Liu, Y.; Sun, S.; Wang, H.; Geng, X. Phenotype and signaling pathway analysis to explore the interaction between tomato plants and TYLCV in different organs. Plant Sci. 2024, 339, 111955. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, X.; Yao, X.; Zhang, P.; Fang, R.; Ye, J. A 7-amino-acid motif of Rep protein essential for virulence is critical for triggering host defense against Sri Lankan cassava mosaic virus. Mol. Plant Microbe Interact. 2020, 33, 78–86. [Google Scholar] [CrossRef]
- Zhao, S.S.; Li, Y. Current understanding of the interplays between host hormones and plant viral infections. PLoS Pathog. 2021, 17, e1009242. [Google Scholar] [CrossRef]
- Li, T.; Huang, Y.; Xu, Z.S.; Li, Y.; Liu, L.; Wang, X.W. Salicylic acid-induced differential resistance to the tomato yellow leaf curl virus among resistant and susceptible tomato cultivars. BMC Plant Biol. 2019, 19, 173. [Google Scholar] [CrossRef]
- Medina-Puche, L.; Tan, H.; Dogra, V.; Van den Ackerveken, G.; Bouwmeester, H.J.; Kant, M.R. A defense pathway linking plasma membrane and chloroplasts and co-opted by pathogens. Cell 2020, 182, 1109–1124. [Google Scholar] [CrossRef]
- Li, P.; Zhang, L.; Yan, X.T.; Liu, Y.Q.; Liu, L.; Liu, S.S.; Wang, X.W. Suppression of TGA2-mediated salicylic acid defence by tomato yellow leaf curl virus C2 via disruption of TCP7-like transcription factor activity in tobacco. Plant Cell Environ. 2025, 48, 4039–4050. [Google Scholar] [CrossRef]
- VanDoorn, A.; de Vries, M.; Kant, M.R.; Schuurink, R.C. Whiteflies glycosylate salicylic acid and secrete the conjugate via their honeydew. J. Chem. Ecol. 2015, 41, 52–58. [Google Scholar] [CrossRef]
- Walling, L.L. Avoiding effective defenses: Strategies employed by phloem-feeding insects. Plant Physiol. 2008, 146, 859–866. [Google Scholar] [CrossRef]
- Peng, W.; Sheng, S.; Liu, K.R.; Rong, P.; Na, L.; Hu, B.; Wang, L.M.; Wang, H.H.; Afzal, A.J.; Geng, X.Q. Physiological and transcriptomic analyses revealed gene networks involved in heightened resistance against tomato yellow leaf curl virus infection in salicylic acid and jasmonic acid treated tomato plants. Front. Microbiol. 2022, 13, 970139. [Google Scholar] [CrossRef]
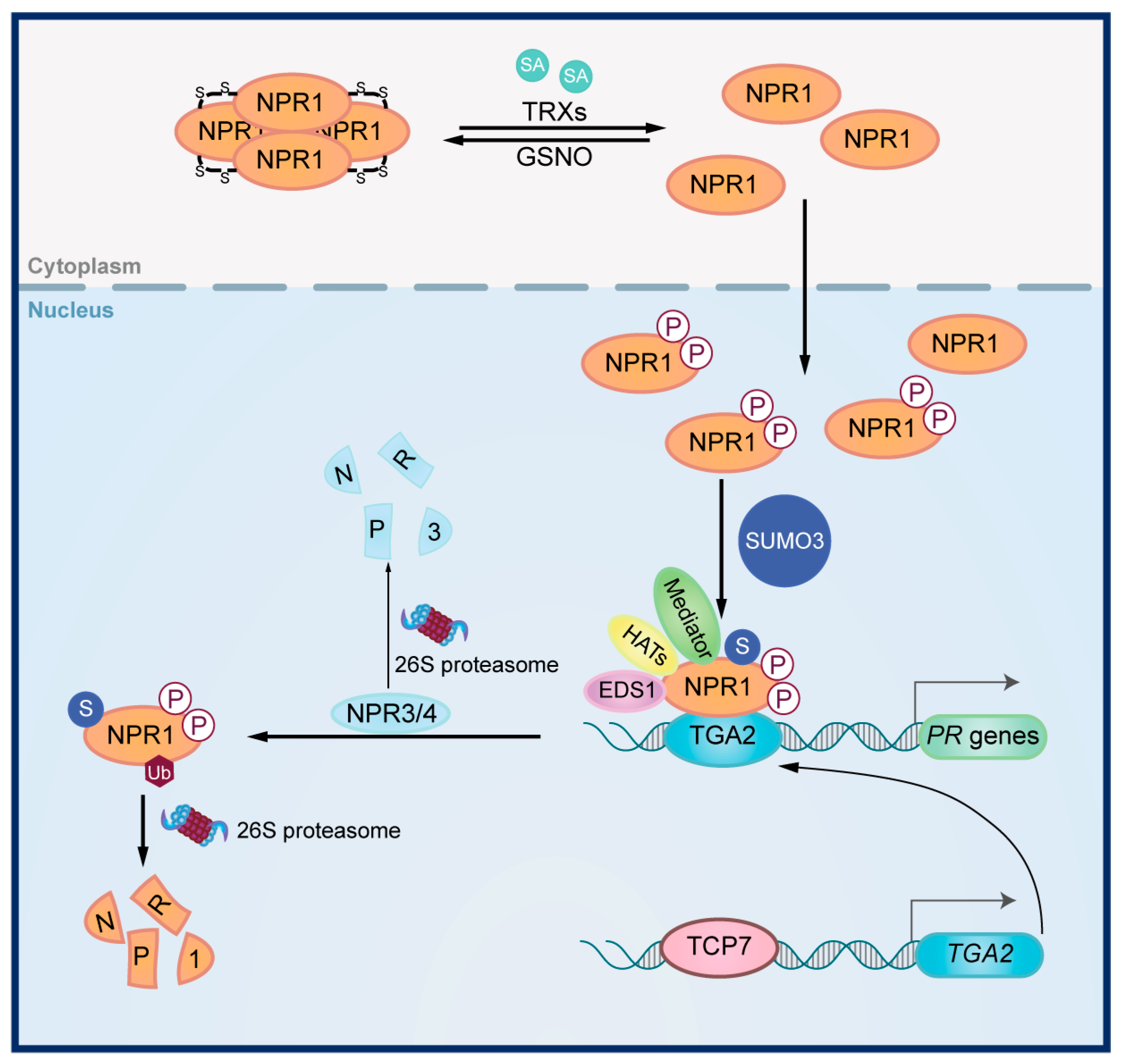

{kind=link}
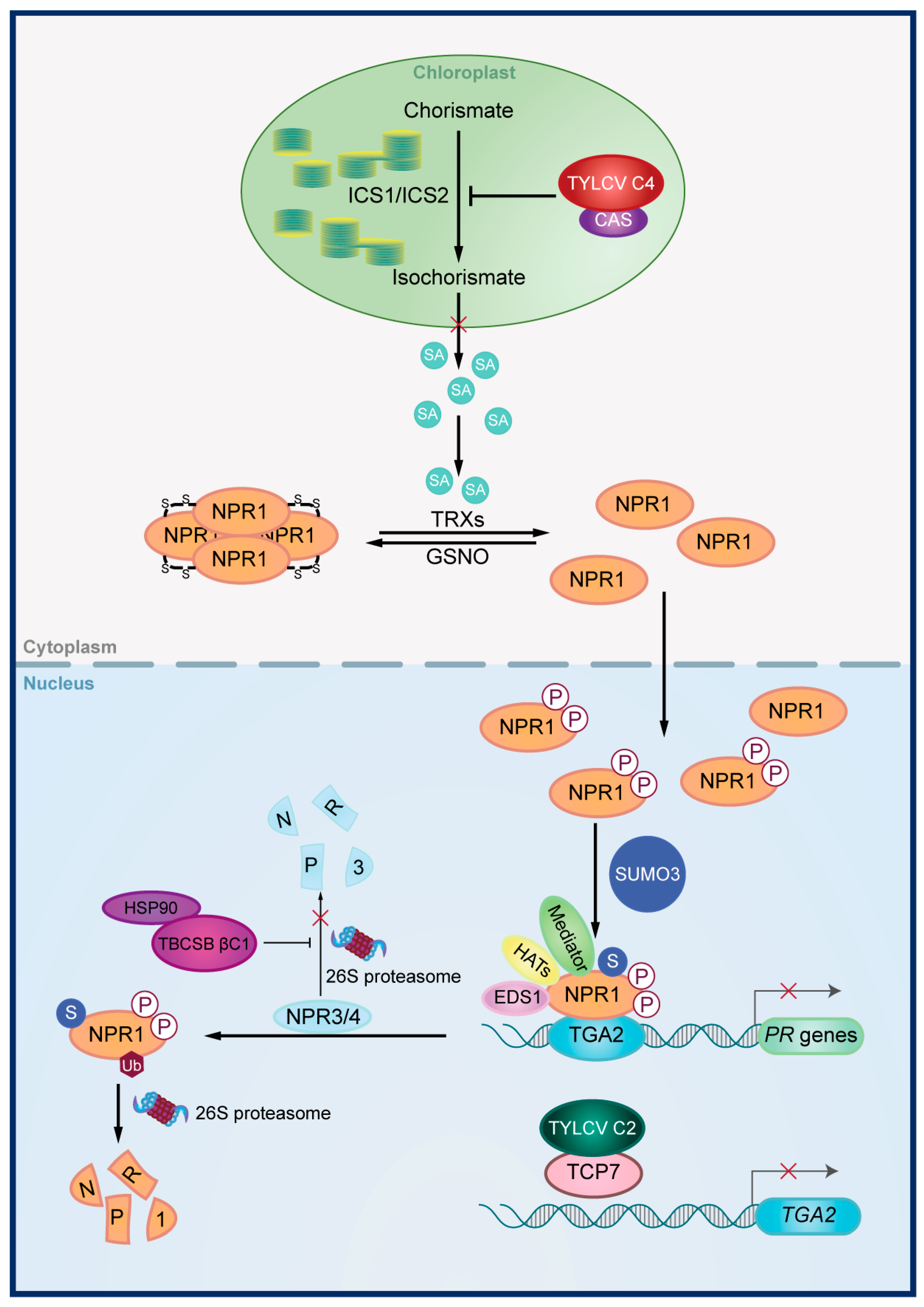
{kind=link}
| Whitefly Species | Developmental Stage | Plant Species | Effect on SA Signaling Pathway | Reference |
|---|---|---|---|---|
| MEAM1 | Nymph | Arabidopsis thaliana | Increased expression of SA-related genes (PR1, BGL2, PR5, SID2, EDS5, and PAD4) | [47] |
| MEAM1 | Adult | Phaseolus lunatus | Increased SA content | [48] |
| MEAM1 | Adult | Solanum lycopersicum | Increased expression of SA-related genes (PAL, PR1, and PR4) | [49] |
| MEAM1 | Adult | S. lycopersicum | Increased SA content | [50] |
| MEAM1 | Nymph | A. thaliana | Increased expression of SA-related genes (EDS1 and PR1) | [51] |
| MEAM1 | Adult | A. thaliana | Increased SA contents and increased expression of SA-related genes (PAD4 and PR1) | [52] |
| MEAM1 | Nymph | Nicotiana tabacum | Increased SA content | [53] |
| MEAM1 | Adult | N. tabacum | Increased SA content | [53] |
| MEAM1 | Nymph | N. tabacum | Increased SA content | [54] |
| MEAM1 | Nymph | Glycine max | Increased SA content | [55] |
| MEAM1 | Adult | N. tabacum | Increased expression of SA-related genes (PAL, ICS, NPR1, and BGL2) | [56] |
| MEAM1 | Adult | N. tabacum | Increased SA content | [57] |
| MEAM1 | Adult | S. lycopersicum | Increased expression of SA-related genes (PR1a and PR1b) | [58] |
| MEAM1 | Mixed (adult, nymph, and egg) | S. lycopersicum | Increased expression of SA-related genes (PR1) | [59] |
| MEAM1 | Nymph | N. tabacum | Increased SA content and increased expression of SA-related genes (PR1a and PR2a) | [60] |
| MEAM1 | Adult | N. tabacum | Increased SA content and increased expression of SA-related genes (PR1a and PR2a) | [60] |
| MEAM1 | Adult | C. annuum | Unchanged SA content and expression of SA-related genes (PR1) | [61] |
| MEAM1 | Adult | N. tabacum | Increased SA content | [62] |
| MEAM1 | Adult | N. tabacum | Increased expression of SA-related genes (PR1 and BGL) | [63] |
| MEAM1 | Egg | N. tabacum | Increased SA content and increased expression of SA-related genes (PR1b, PR1c, PR1, and PR5) | [64] |
| MEAM1 | Adult | N. tabacum | Increased SA content | [65] |
| MEAM1 | Adult | S. lycopersicum | Increased SA content | [65] |
| MEAM1 | Adult | N. benthamiana | Increased SA content | [65] |
| MED | Adult | S. lycopersicum | Increased SA content | [66] |
| MED | Adult | S. lycopersicum | Increased SA content | [67] |
| MED | Adult | C. annuum | Unchanged SA content and expression of SA-related genes (PR1) | [61] |
| Unspecified | Adult | Capsicum annuum | Increased expression of SA-related genes (PR1, PR4, and PR10) | [68] |
| Unspecified | Adult | C. annuum | Increased expression of SA-related genes (BGL, PR1, PR5, and Hin1) | [69] |
| Viral Species | Plant Species | Response of the SA Signaling Pathway | Reference |
|---|---|---|---|
| Cabbage leaf curl virus | A. thaliana | Increased expression of SA-related genes (EDS1, PAD4, SAG101, FMO1, ALD1, SID2, EDS5, NPR1, NPR2, NPR3, NPR4, WRKY70, TGA1, TGA3, TGA5, PR1, PR2, and PR5) | [73] |
| Euphorbia mosaic virus | C. annuum | Increased SA content and increased expression of SA-related genes (NPR1 and PR10) | [74] |
| Mungbean yellow mosaic India virus | Vigna mungo | Increased expression of SA-related genes (PR1 and PAL) | [75] |
| Tobacco curly shoot virus | N. benthamiana | Unchanged SA content and increased expression of SA-related genes (PR1a and PR2) | [65] |
| Tomato leaf curl New Delhi virus | S. tuberosum | Increased expression of SA-related genes (PAL) | [76] |
| Tomato leaf curl Palampur virus | S. lycopersicum | Increased expression of SA-related genes (NPR1, PAD4, PAL, PR1, and PR5) | [77] |
| Tomato yellow leaf curl Sardinia virus | S. lycopersicum | Decreased expression of SA-related genes (PAL) | [78] |
| Tomato yellow leaf curl virus | S. lycopersicum | Increased SA content and increased expression of SA-related genes (PR1) | [79] |
| Tomato yellow leaf curl virus | S. lycopersicum | Increased SA content and increased expression of SA-related genes (PR1a) | [80] |
| Tomato yellow leaf curl virus | S. lycopersicum | Increased expression of SA-related genes (PR1) | [81] |
| Tomato yellow leaf curl virus | S. lycopersicum | Increased SA content and increased expression of SA-related genes (PAL and PR1) | [82] |
| Tomato yellow leaf curl virus | S. lycopersicum | Increased SA content and increased expression of SA-related genes (PR1) | [82] |
| Tomato yellow leaf curl virus | S. lycopersicum | Increased expression of SA-related genes (PR1, NPR1, PAD4 and EDS1) | [83] |
| Sri Lankan cassava mosaic virus | N. benthamiana | Increased expression of SA-related genes (NPR1, PR1a and PR5) | [84] |
| Viral Species | Viral Effector | Plant Species | Modulation of the SA Signaling Pathway | Reference |
|---|---|---|---|---|
| Tobacco curly shoot virus | βC1 | N. benthamiana | Decreased expression of SA-related genes (PR1a and PR2) | [77] |
| Tomato leaf curl Palampur virus | AV2 | N. benthamiana | Increased expression of SA-related genes (NPR1, PR1, and PR5) | [77] |
| Tomato yellow leaf curl virus | C4 | A. thaliana | Decreased SA content and expression of SA-related genes (PR1) | [87] |
| Tomato yellow leaf curl virus | C2 | N. tabacum | Decreased expression of SA-related genes (BGL2) | [88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.-X.; Wang, S.-D.; Liu, Y.-Q.; Pan, L.-L. Modulation of Plant Interactions with Whitefly and Whitefly-Borne Viruses by Salicylic Acid Signaling Pathway: A Review. Viruses 2025, 17, 825. https://doi.org/10.3390/v17060825
Zhao S-X, Wang S-D, Liu Y-Q, Pan L-L. Modulation of Plant Interactions with Whitefly and Whitefly-Borne Viruses by Salicylic Acid Signaling Pathway: A Review. Viruses. 2025; 17(6):825. https://doi.org/10.3390/v17060825
Chicago/Turabian StyleZhao, Shi-Xing, Su-Dan Wang, Yin-Quan Liu, and Li-Long Pan. 2025. "Modulation of Plant Interactions with Whitefly and Whitefly-Borne Viruses by Salicylic Acid Signaling Pathway: A Review" Viruses 17, no. 6: 825. https://doi.org/10.3390/v17060825
APA StyleZhao, S.-X., Wang, S.-D., Liu, Y.-Q., & Pan, L.-L. (2025). Modulation of Plant Interactions with Whitefly and Whitefly-Borne Viruses by Salicylic Acid Signaling Pathway: A Review. Viruses, 17(6), 825. https://doi.org/10.3390/v17060825