1. Introduction
Avian infectious bronchitis (IB) is a viral, endemic, and highly contagious disease primarily affecting chickens. It causes respiratory, renal, and reproductive diseases and engenders significant economic losses to the poultry industry worldwide [
1]. In broilers, financial losses are mainly attributed to a weight gain decrease and a high mortality rate, especially in infection with nephropathogenic strains. In layers, the main signs include an egg production decrease and a deterioration in internal and external egg quality [
2]. The infectious bronchitis virus (IBV) is a single-stranded RNA virus of the
Coronaviridae family that encodes four structural proteins: spike (S), membrane (M), envelope (E), and nucleoprotein (N) [
3,
4,
5,
6,
7]. IBV is prone to changes through spontaneous mutations or genetic recombination, leading to the emergence of new variants [
8]. In Morocco, the first isolation and characterization of IBV were reported in 1983 by El Houadfi and Jones. The “Moroccan G strain” was identified in 1986, and phylogenetic studies suggested a shared origin with the 4/91 serotype [
9]. Between 1996 and 2005, three IBV genotypes were identified: Massachusetts (Mass.), a strain related to 4/91, and a nephropathogenic variant [
10]. In 2005, five nephropathogenic IBV strains were identified in Morocco [
11]. From 2009 to 2014, novel genotypes emerged, including Italy 02, which made up 32% of isolates in this period [
12]. Since 2015, a new IBV lineage has been detected in different regions of Morocco, with related viruses observed in Algeria in 2012 and Tunisia in 2016 [
13]. The poultry disease status in Morocco has worsened since 2016 following the introduction of AI H9N2 and its ability to cause disease alone or in combination with other pathogens [
14].
LPAIV H9N2 is a highly infectious pathogen that affects birds and humans [
15]. Due to its potential to reassort with other influenza viruses and to evolve into a more threatening strain, monitoring farms for H9N2 has become crucial to assess the virus’s spread and its evolution. Respiratory complex diseases often have a multifactorial nature, as more than one pathogen can be detected in the same flock, such as infectious bronchitis virus (IBV), low pathogenic avian influenza virus (LPAIV) H9N2, infectious laryngotracheitis (ILT), Newcastle disease virus (NDV),
Mycoplasma gallisepticum (MG),
Mycoplasma synoviae (MS), and
Escherichia coli (E. coli). Coinfections between viral agents or viral and bacterial pathogens are common in poultry farms, often resulting in more severe clinical symptoms than single infections [
16].
This study aims to (i) determine the prevalence of IBV and LPAI H9N2 in Moroccan poultry farms, (ii) identify the current IBV genotypes and lineages circulating in the country, and (iii) investigate the correlations between different avian pathogens in respiratory coinfection diseases.
4. Discussion
Infectious bronchitis virus (IBV) and low-pathogenic avian influenza virus (LPAIV) H9N2 subtype are high-risk pathogens posing serious threats to animal health, public health, and the economy. The World Organization for Animal Health (WOAH) identifies both viruses as significant dangers to the poultry industry and public health due to the outbreak potential of IBV and the zoonotic risk of H9N2 [
24,
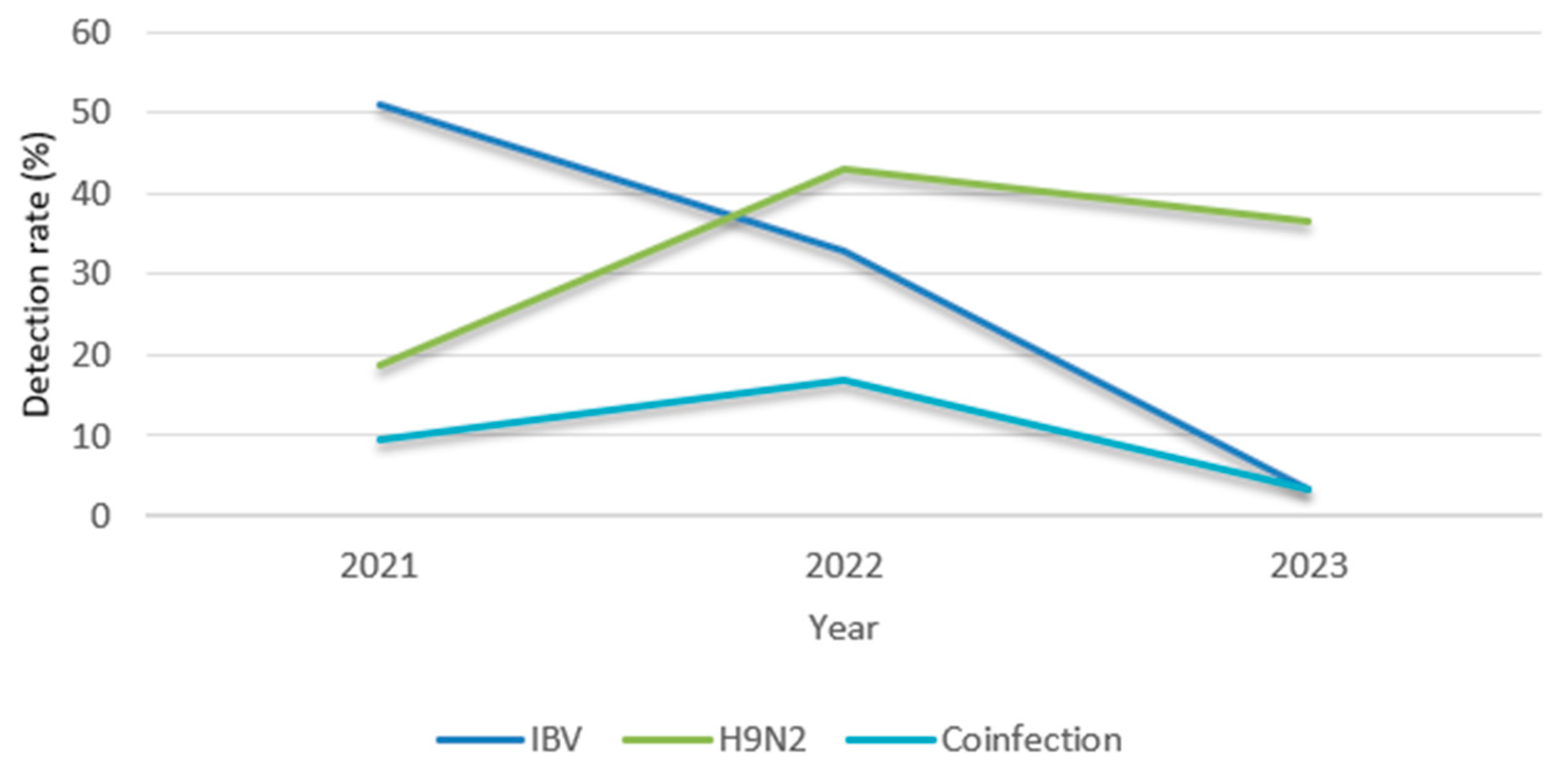
25]. This study investigated the prevalence of IBV and H9N2 in 183 broiler farms across seven regions of Morocco from 2021 to 2023. The detection rates were 33.33% for IBV and 34.97% for H9N2, which were nearly equivalent, suggesting that common risk factors facilitate their spread. Previous studies reported higher prevalence rates, with 51.7% of the flocks testing positive for IBV and 58% for H9N2 [
12,
26]. The coinfection rate was 12.57%, with a peak of 17% observed in 2022. The same year, H9N2 cases peaked at 44%, indicating increased viral circulation. Yearly variations in infection rates have been observed in other studies. They could be attributed to various reasons, including differences in biosecurity and control measures, poultry density, and the hygienic conditions of the studied farms [
27,
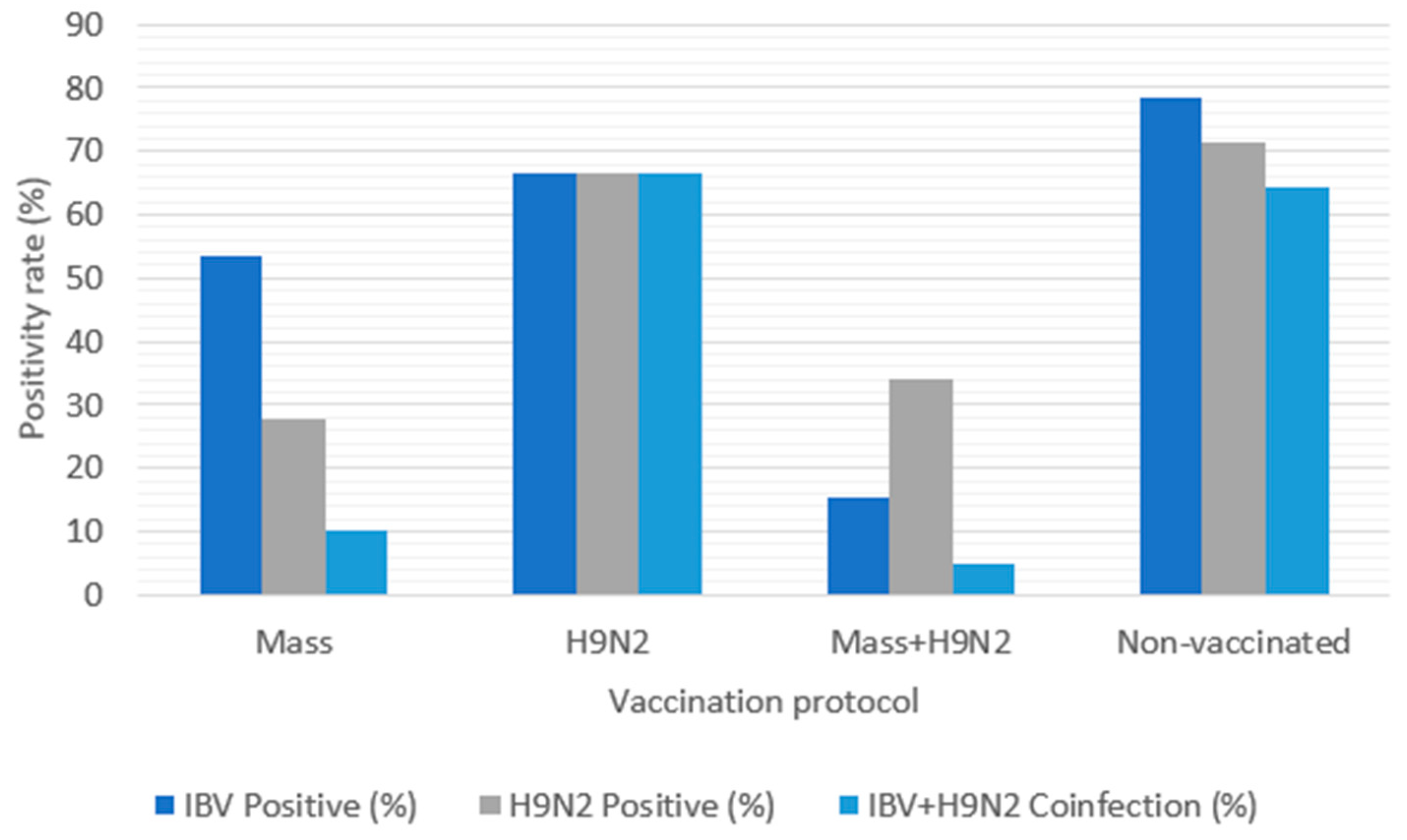
28]. Despite extensive vaccination coverage, 77.05% of vaccinated farms tested positive for IBV and 57.81% for H9N2. Positive cases showed high mortality rates and respiratory symptoms, with predominant respiratory lesions including congestive tracheitis, airsacculitis, and sinusitis. Tracheitis is a common clinical sign of IBV infections due to the viral damage of ciliated epithelial cells [
29]. H9N2 also affects the respiratory tract and is characterized by airsacculitis, tracheitis, and sinus enlargement [
30]. Broilers aged 21–40 days were particularly susceptible to these coinfections due to their immature immune systems, which may be further compromised by stress and suboptimal environmental conditions [
31].
Field data showed an exacerbation of clinical signs and gross lesions in cases co-infected with IBV and H9N2, demonstrating that mixed infections enhance pathogenicity and increase mortality rates, reaching 44.26%, causing substantial economic losses [
32,
33,
34]. Furthermore, malpractices, inadequate security measures, and/or stress factors could affect the weekly mortality rate, ranging from 0.1% to 16.4% [
35].
However, the infection sequence may contribute to pathogenic differences [
36,
37]. Several studies have shown that coinfection with IBV and H9N2 enhances clinical signs, irrespective of the IBV strain involved, and extends the shedding period of H9N2 [
37,
38,
39,
40]. This synergistic effect is thought to result from the IBV-induced severe inflammatory response, which increases the pathogenicity of H9N2 [
36]. It could also result from the presence of a trypsin-like serine protease encoded by IBV, which facilitates the cleavage activation of H9N2 hemagglutinin, thereby improving its replication [
41,
42]. In contrast, in vitro and ovo studies reported different results, indicating viral interference that was not observed in vivo. The quantitative results of simultaneous coinfections with H9N2 and IBV revealed that interference between the two viruses led to a reduction in viral growth. However, in cases of superinfection, the second virus—whether H9N2 or IBV—reduced the growth of the initially inoculated virus [
38]. These findings may inform future strategies for virus control and the development of appropriate vaccination protocols.
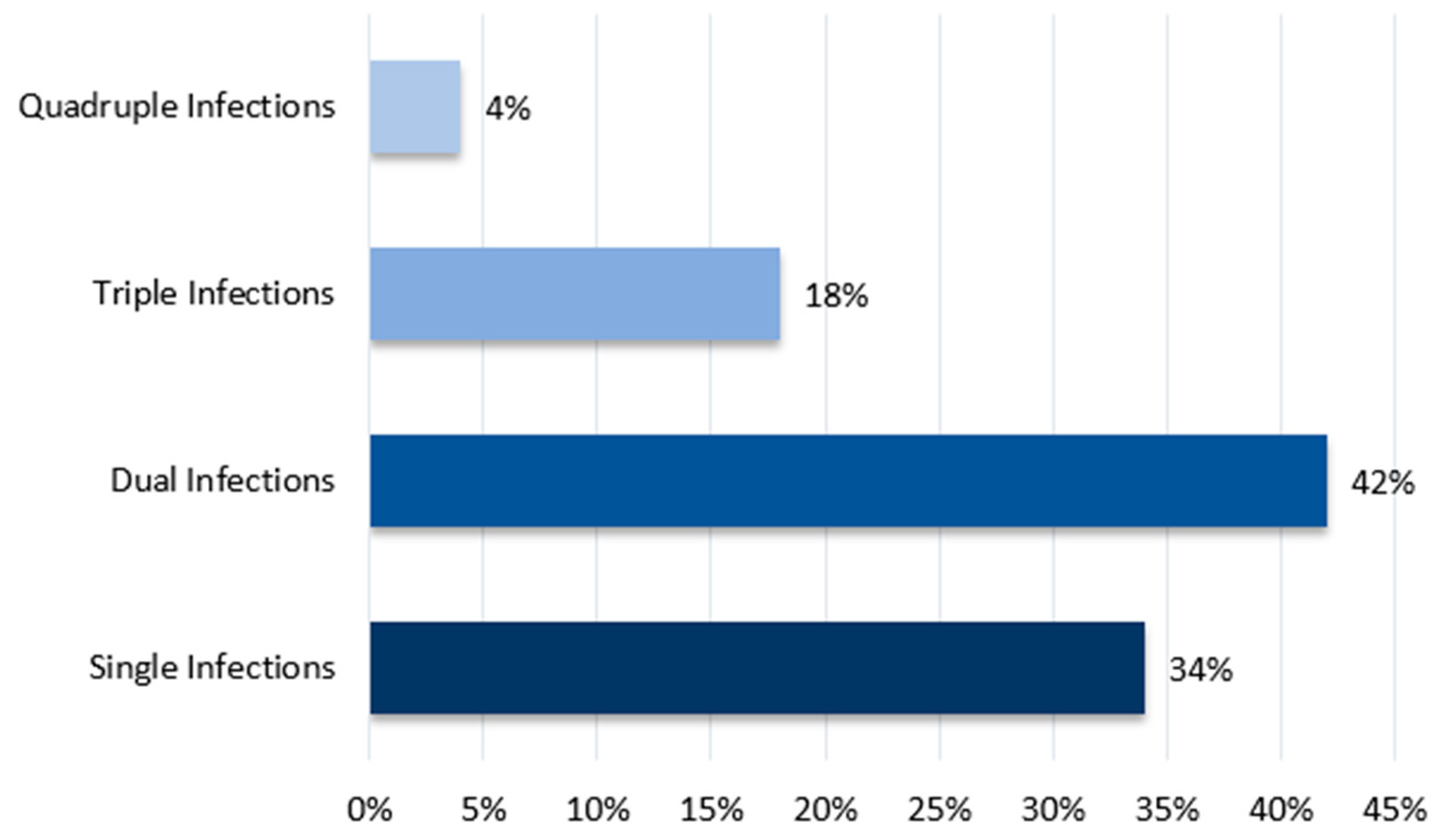
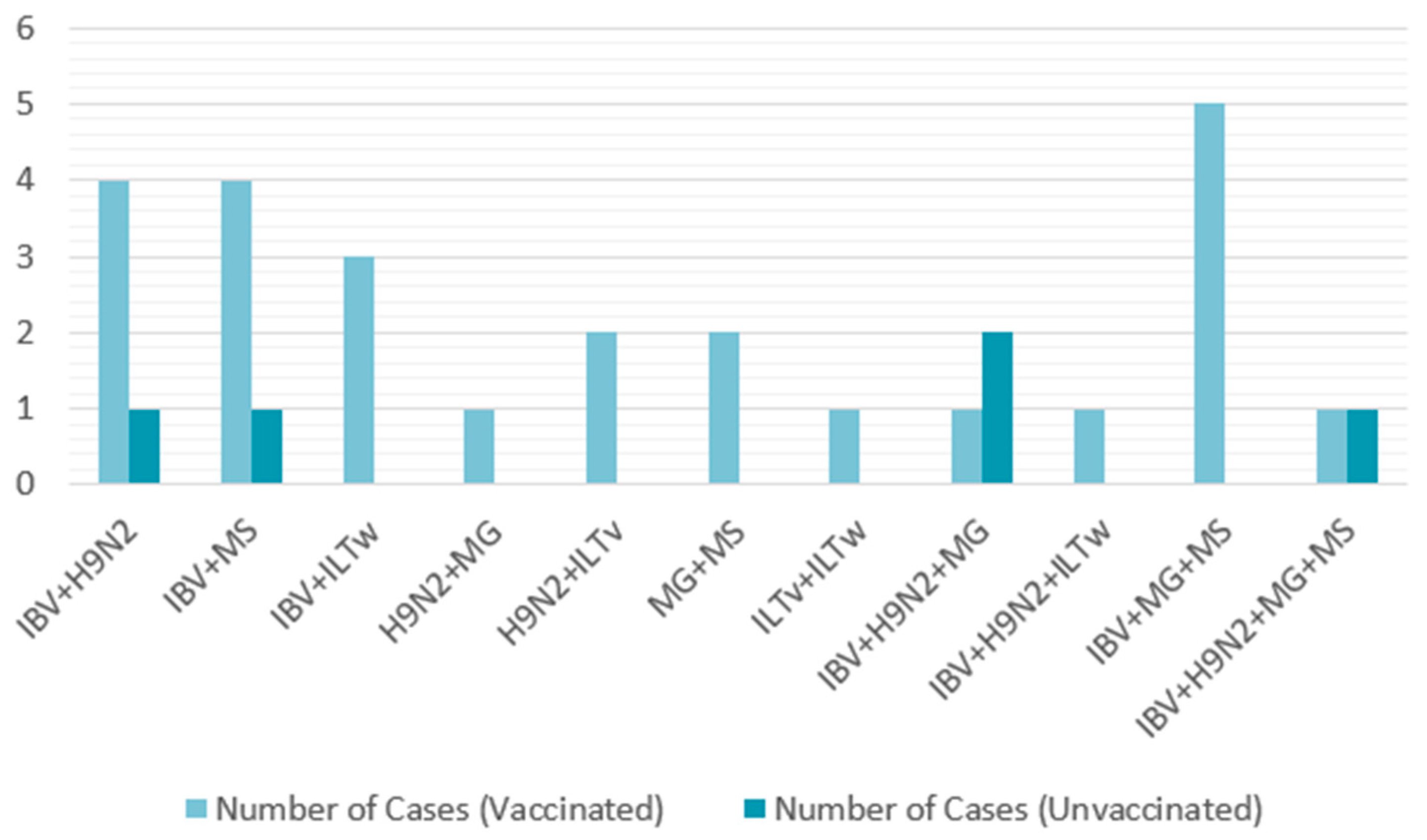
To further explore potential coinfections with other pathogens, 50 farms positive for IBV were randomly selected from the 183 previously studied. These farms were tested for other respiratory pathogens. Single infections were more common, detected in 37.16% of cases, whereas coinfections had a detection rate of 22.95%. The most frequent combinations were IBV + H9N2 and IBV + MS for dual infections, IBV + H9N2 + MG and IBV + MG + MS for triple infections, and IBV + H9N2 + MG + MS for a notable quadruple infection. Therefore, IBV emerged as the predominant virus that facilitated the introduction of other pathogens. A similar study conducted by Jbenyeni et al. from 2018 to 2020 in Tunisia showed a high prevalence of IBV and LPAIV H9N2 in poultry farms, with the same major viral combination of IBV + H9N2, stressing H9N2’s contribution to multifactorial respiratory diseases [
43]. In Ghana, the detection of H9N2 was frequently associated with IBV detection, causing an exacerbation of clinical signs [
44].
Regarding the geographic distribution of pathogens in Morocco, Rabat-Sale-Kenitra registered the highest diversity of infections, with IBV + H9N2 being the most common dual infection. Casablanca-Settat also recorded several cases involving IBV, MG, and MS. Additionally, quadruple infections were detected in these regions, indicating their status as hotspots for viral activity and numerous pathogens. The high prevalence observed in these two regions is likely attributed to the high poultry activity due to the growing industry in these areas. Arbani et al. have reported a high prevalence of LPAI H9N2 in the same regions [
45]. In contrast, fewer cases were reported in Marrakech-Safi, while only one was detected in the Eastern region, suggesting regional variation in infection dynamics.
Another notable pathogen detected in our study was ILT. While infections involving this virus were less frequent, its presence in dual and triple infections across three regions suggested ILT may synergistically exacerbate respiratory symptoms. Several studies have demonstrated that coinfections contribute to enhanced respiratory signs and increased mortality rates [
46,
47]. For example, chickens co-infected with IBV and H9N2 showed more severe clinical signs. Similarly, a study of the effect of LPAIV H3N8 and MG coinfection found an increased MG colonization rate in lungs and internal organs, which resulted in higher mortality rates [
46]. These findings were supported by a challenge study using H9N2 and MG, where co-inoculation caused increased tracheal cast formation and higher mortality [
48]. Another study highlighted the synergistic role of MS and IBV in enhancing inflammation in the respiratory tract and exacerbating lesions, particularly in airsacculitis syndrome [
49].
The phylogenetic analysis showed that all IBV viruses isolated belonged to two distinct genotypes: 793B (GI-13) and D181 (GII-1). The homology analysis showed that the 793B strain was similar to the Spain/92/51, Morocco/4-91/IBV02/2014, and Morocco/4-91/IBV16/2014 strains, and the sequence identity ranged from 99% to 100%.
First isolated in 1985 in France [
50], 793B entered the United Kingdom in 1991 to finally spread to most European countries [
51]. A 2018 survey in 14 European countries demonstrated that QX and 793B were the most prevalent strains [
52]. The Middle East registered 43.66% of 793B cases between 2009 and 2014 [
53], and Asia registered a high prevalence of the strain, especially in Iran (42.8%) [
54]. The strain was named “G”, but in 2004 sequencing, it was shown that the variant was genetically linked to the 4/91 genotype [
12,
55]. Since then, 793B and Massachusetts (Mass.) have been the most prevalent serotypes worldwide [
56]. In a broader context, IBV registered an important genetic diversity worldwide, with the most common serotypes being Mass-type, 4/91 (793B or CR88)-like, D274-like (D207, D212 or D1466, D3896), D3128, QX-like, and Italy02 [
57]. While the Mass. and Italy02 genotypes were prevalent in Morocco between 2010 and 2014, these genotypes were not detected during this study, highlighting a shift in the distribution of strains in Morocco [
12]. The same trend was seen in Europe, where the Italy02 genotype, prevalent in 2023, has declined since then [
58]. In China, while the QX genotype has increased, vaccine-like genotypes such as Mass. decreased from 50.4% to 4.4% [
59]. The shifts are mainly caused by a change in poultry management, viral mutations and recombination, and vaccination practices [
60,
61]. Phylogenetic clustering revealed a clear distinction among Moroccan isolate genotypes, with no evidence of other lineages circulating. The Moroccan IBV strains clustered closely with European and Middle Eastern isolates, especially with the 793B and D181 lineages. These detections suggest a recent introduction via the poultry trade with European countries [
62], or a late detection due to the late initiation of sequencing efforts [
63]. Remarkably, the D181 strain was detected for the first time in Morocco and Africa. First reported by Molenaar et al. in 2020, D181 is considered a new serotype and the second lineage within genotype II (GII) [
64,
65]. Phylogenetic analysis showed 90.9–95% similarity with D181 strains isolated in Dutch layers (strain CK/NL/D181/2018) [
65].
In summary, various respiratory pathogens are circulating in Moroccan broiler farms, with the prevalence of IBV and H9N2 playing a key role in exacerbating multiple infections and compromising the immune system of chickens. Despite extensive monitoring and vaccination efforts, the presence of several viruses has reduced vaccine efficacy and complicated effective strategy implementation for controlling viral spread and mitigating disease severity. These findings suggest that the current vaccination strategies do not provide sufficient protection. The vaccination limitations may be attributed to various factors: antigenic variation between vaccine strains, improper vaccine administrations, or co-infections with other pathogens that could enhance viral pathogenicity. These factors compromise chickens’ immune response and optimize viral spread [
66]. Therefore, vaccination alone does not provide sufficient protection. The need to regularly monitor field strains, adapt vaccination programs, and instill good health management strategies could help reduce poultry losses [
66]. To achieve better goals, it is essential to implement massive standardized vaccination strategies, control flock density, and improve biosecurity practices [
67].
Interactions between different viral respiratory pathogens play a crucial role in determining the outcome of coinfections. However, most studies on viral interference in complex respiratory infections fail to fully capture the field conditions, where chickens are usually exposed to multiple infectious agents simultaneously [
32,
38,
39,
68].
This study underscores the severity of viral coinfections that resulted in high morbidity and mortality rates. Respiratory diseases are complex and deserve more attention as they threaten the poultry industry. The high prevalence of viral avian diseases in Morocco can be attributed to poor sanitary conditions and inadequate vaccination programs. Our findings emphasize the urgent need to implement effective sanitary measures to limit the emergence of new variants and the further spread of viruses across the country. This approach should focus on updating vaccination programs to cover circulating genotypes, strengthen biosecurity measures to reduce viral introduction, improve vaccine administration, and reduce stress factors to decrease chickens’ susceptibility to infections [
66].

 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}