
Characterization of an Emerging Recombinant Duck Circovirus in Northern Vietnam, 2023–2024

 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Samples
2.3. DNA Extraction and Polymerase Chain Reaction (PCR)
2.4. Nucleotide Sequencing and Analysis
2.5. Recombination Events and Selection Profiles of Vietnamese DuCV Strains
3. Results
3.1. Detection of Duck Circovirus in Field Samples
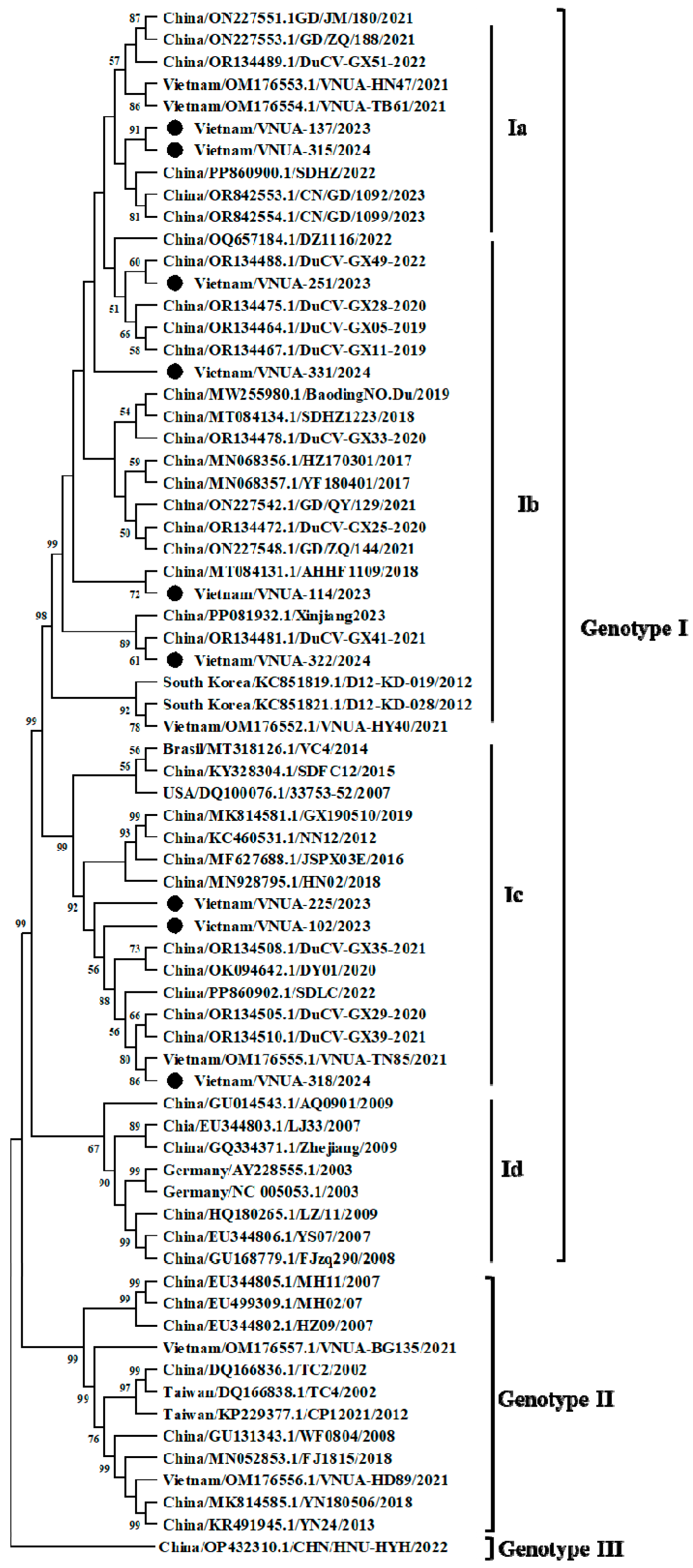
3.2. Characterization of Vietnamese DuCV Genomes
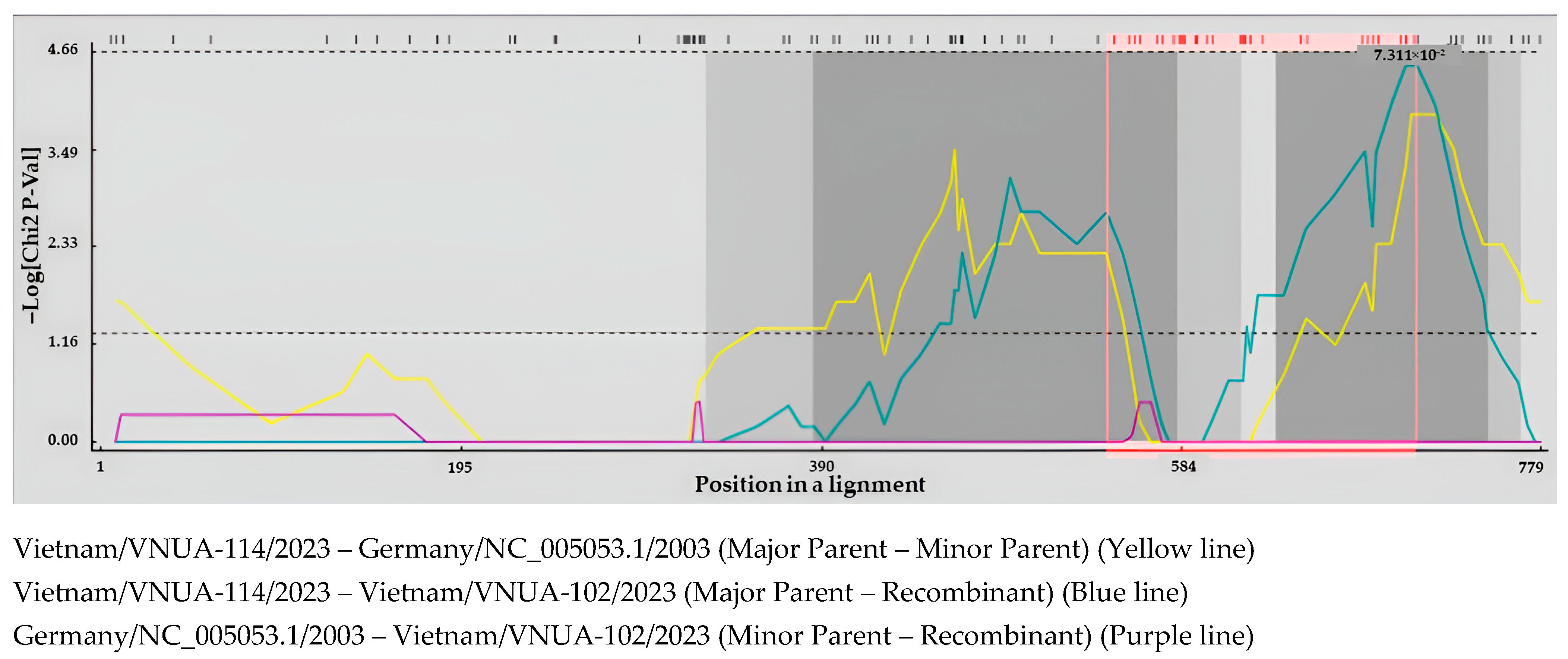
3.3. Recombination Analysis of Vietnamese DuCV Strains
3.4. Analysis of Natural Selection Profile of DuCV Gene Sequences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lei, X.; Wang, A.; Zhu, S.; Wu, S. From obscurity to urgency: A comprehensive analysis of the rising threat of duck circovirus. Vet. Res. 2024, 55, 12. [Google Scholar] [CrossRef] [PubMed]
- Hattermann, K.; Schmitt, C.; Soike, D.; Mankertz, A. Cloning and sequencing of Duck circovirus (DuCV). Arch. Virol. 2003, 148, 2471–2480. [Google Scholar] [CrossRef]
- Wang, X.; Li, L.; Shang, H.; Zhou, F.; Wang, C.; Zhang, S.; Gao, P.; Guo, P.; Zhu, R.; Sun, Z.; et al. Effects of duck circovirus on immune function and secondary infection of Avian Pathogenic Escherichia coli. Poult. Sci. 2022, 101, 101799. [Google Scholar] [CrossRef]
- Hong, Y.T.; Kang, M.; Jang, H.K. Pathogenesis of duck circovirus genotype 1 in experimentally infected Pekin ducks. Poult. Sci. 2018, 97, 3050–3057. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.Y.; Xiong, W.J.; Tang, H.; Xiao, C.T. Identification and characterization of a novel circovirus species in domestic laying ducks designated as duck circovirus 3 (DuCV3) from Hunan province, China. Vet. Microbiol. 2022, 275, 109598. [Google Scholar] [CrossRef]
- Ji, J.; Chen, Q.; Sui, C.; Yu, Z.; Xu, X.; Yao, L.; Kan, Y.; Bi, Y.; Xie, Q. Novel genotype definition and genome characteristics of duck circovirus in central and Eastern China. Transbound. Emerg. Dis. 2020, 67, 2993–3004. [Google Scholar] [CrossRef]
- Xiang, Q.W.; Wang, X.; Xie, Z.J.; Sun, Y.N.; Zhu, Y.L.; Wang, S.J.; Liu, H.J.; Jiang, S.J. ORF3 of duck circovirus: A novel protein with apoptotic activity. Vet. Microbiol. 2012, 159, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Banda, A.; Galloway-Haskins, R.I.; Sandhu, T.S.; Schat, K.A. Genetic analysis of a duck circovirus detected in commercial Pekin ducks in New York. Avian Dis. 2007, 51, 90–95. [Google Scholar] [CrossRef]
- Wen, H.; Wu, Y.; Yang, C.; Zhang, X.; Lian, C.; Chen, H.; Han, L. Comments on duck circovirus (DuCV) genotype definition. Gene 2014, 538, 207–208. [Google Scholar] [CrossRef]
- Zhang, Z.; Jia, R.; Lu, Y.; Wang, M.; Zhu, D.; Chen, S.; Yin, Z.; Chen, X.; Cheng, A. Identification, genotyping, and molecular evolution analysis of duck circovirus. Gene 2013, 529, 288–295. [Google Scholar] [CrossRef]
- Yu, C.D.; Kim, S.W.; Liu, C.X.; Gao, Y.H.; Li, Y.F.; Park, J.Y.; Cha, S.Y.; Jang, H.K.; Kang, M.; Wei, B. Epidemiological investigation and genetic analysis of duck circovirus in Korea from 2013 to 2022. Animals 2024, 14, 3630. [Google Scholar] [CrossRef] [PubMed]
- Kulprasertsri, S.; Songserm, T.; Phatthanakunanan, S.; Saengnual, P.; Sinwat, N.; Khamtae, R.; Lertwatcharasarakul, P. Molecular genotyping and subgenotyping of duck circovirus at duck farms in Thailand. Vet. World 2024, 17, 1990–1999. [Google Scholar] [CrossRef] [PubMed]
- Julian, L.; Piasecki, T.; Chrzastek, K.; Walters, M.; Muhire, B.; Harkins, G.W.; Martin, D.P.; Varsani, A. Extensive recombination detected among beak and feather disease virus isolates from breeding facilities in Poland. J. Gen. Virol. 2013, 94, 1086–1095. [Google Scholar] [CrossRef]
- Cha, S.Y.; Kang, M.; Cho, J.G.; Jang, H.K. Genetic analysis of duck circovirus in Pekin ducks from South Korea. Poult. Sci. 2013, 92, 2886–2891. [Google Scholar] [CrossRef]
- Fringuelli, E.; Scott, A.N.; Beckett, A.; McKillen, J.; Smyth, J.A.; Palya, V.; Glavits, R.; Ivanics, E.; Mankertz, A.; Franciosini, M.P.; et al. Diagnosis of duck circovirus infections by conventional and real-time polymerase chain reaction tests. Avian Pathol. 2005, 34, 495–500. [Google Scholar] [CrossRef]
- Soike, D.; Albrecht, K.; Hattermann, K.; Schmitt, C.; Mankertz, A. Novel circovirus in mulard ducks with developmental and feathering disorders. Vet. Rec. 2004, 154, 792–793. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, N.; Zhang, L.; Jiang, W.; Fan, X.; Wang, X.; Miao, R.; Zhai, X.; Wei, L.; Jiang, S.; et al. Research Note: Complete genome cloning and genetic evolution analysis of four Cherry Valley duck circovirus strains in China in 2022. Poult. Sci. 2023, 102, 102920. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Yao, X.Y.; Yang, H.H.; Zhang, Y.Q.; Liu, H.; Sun, J.; Lv, Z.H.; Huang, S.J.; Zhang, X.L. Research note: Genetic diversity of duck circoviruses circulating in partial areas of Guangdong province, southern China. Poult. Sci. 2022, 101, 102032. [Google Scholar] [CrossRef]
- Tran, G.T.H.; Mai, N.T.; Bui, V.N.; Dao, T.D.; Trinh, D.Q.; Vu, T.T.T.; Le, V.P.; Dong, V.H. Duck circovirus in northern Vietnam: Genetic characterization and epidemiological analysis. Arch. Virol. 2022, 167, 1871–1877. [Google Scholar] [CrossRef]
- Jiang, S.J.; Zhang, X.X.; Liu, S.N.; Wang, Y.; Kong, T.B.; Wei, X.L.; Sun, Y.N.; Zhao, Q. PCR detection and sequence analysis of duck circovirus in sick muscovy ducks. Virol. Sin. 2008, 23, 265–271. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Murrell, B.; Moola, S.; Mabona, A.; Weighill, T.; Sheward, D.; Kosakovsky Pond, S.L.; Scheffler, K. FUBAR: A fast, unconstrained bayesian approximation for inferring selection. Mol. Biol. Evol. 2013, 30, 1196–1205. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Bai, C.X.; Guo, X.; Gao, W.H.; Li, M.L.; Wang, J.; Li, Y.D. Molecular characteristics of a novel duck circovirus subtype 1d emerging in Anhui, China. Virus Res. 2021, 295, 198216. [Google Scholar] [CrossRef]
- Desvaux, S.; Nguyen, C.O.; Vu, D.T.; Henriquez, C.; Ky, V.D.; Roger, F.; Fenwick, S.; Goutard, F. Risk of Introduction in Northern Vietnam of HPAI Viruses from China: Description, Patterns and Drivers of Illegal Poultry Trade. Transbound. Emerg. Dis. 2016, 63, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Vijaykrishna, D.; Duan, L.; Bahl, J.; Zhang, J.X.; Webster, R.G.; Peiris, J.S.; Chen, H.; Smith, G.J.; Guan, Y. Identification of the progenitors of Indonesian and Vietnamese avian influenza A (H5N1) viruses from southern China. J. Virol. 2008, 82, 3405–3414. [Google Scholar] [CrossRef]
- Sun, W.; Zheng, M.; Cao, H.; Lu, H.; Wei, X.; Pan, Y.; Zhang, H.; Su, J.; Li, J.; Jin, N. Genome sequences of a vovel recombinant duck circovirus in China. Genome Announc. 2016, 4, e01181-16. [Google Scholar] [CrossRef]
- Wang, D.; Xie, X.; Zhang, D.; Ma, G.; Wang, X.; Zhang, D. Detection of duck circovirus in China: A proposal on genotype classification. Vet. Microbiol. 2011, 147, 410–415. [Google Scholar] [CrossRef]
- Grenfell, B.T.; Pybus, O.G.; Gog, J.R.; Wood, J.L.; Daly, J.M.; Mumford, J.A.; Holmes, E.C. Unifying the epidemiological and evolutionary dynamics of pathogens. Science 2004, 303, 327–332. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | GenBank Accession Number | Strain | Country | Host | Year | Genotype |
|---|---|---|---|---|---|---|
| 1 | OR842553.1 | CN/GD/1092 | China | Duck | 2023 | I |
| 2 | OR842554.1 | CN/GD/1099 | China | Duck | 2023 | I |
| 3 | PP860902.1 | SDLC | China | Duck | 2023 | I |
| 4 | PP081932.1 | Xinjiang2023 | China | Duck | 2023 | I |
| 5 | OR134489.1 | DuCV-GX51 | China | Duck | 2022 | I |
| 6 | PP860900.1 | SDHZ | China | Duck | 2022 | I |
| 7 | OQ657184.1 | DZ1116 | China | Duck | 2022 | I |
| 8 | OR134488.1 | GX49 | China | Duck | 2022 | I |
| 9 | OM176552.1 | VNUA-HY40 | Vietnam | Duck | 2021 | I |
| 10 | OM176553.1 | VNUA-HN47 | Vietnam | Duck | 2021 | I |
| 11 | OM176554.1 | VNUA-TB61 | Vietnam | Duck | 2021 | I |
| 12 | OM176555.1 | VNUA-TN85 | Vietnam | Duck | 2021 | I |
| 13 | ON227551.1 | GD/JM/180 | China | Duck | 2021 | I |
| 14 | ON227553.1 | GD/ZQ/188 | China | Duck | 2021 | I |
| 15 | ON227542.1 | GD/QY/129 | China | Duck | 2021 | I |
| 16 | ON227548.1 | GD/ZQ/144 | China | Duck | 2021 | I |
| 17 | OK094642.1 | DY01 | China | Duck | 2020 | I |
| 18 | OR134472.1 | GX25 | China | Duck | 2020 | I |
| 19 | OR134475.1 | GX28 | China | Duck | 2020 | I |
| 20 | OR134505.1 | GX29 | China | Duck | 2020 | I |
| 21 | OR134478.1 | GX33 | China | Duck | 2020 | I |
| 22 | OR134508.1 | GX35 | China | Duck | 2021 | I |
| 23 | OR134510.1 | GX39 | China | Duck | 2021 | I |
| 24 | OR134481.1 | GX41 | China | Duck | 2021 | I |
| 25 | OR134464.1 | GX05 | China | Duck | 2019 | I |
| 26 | OR134467.1 | GX11 | China | Duck | 2019 | I |
| 27 | MW255980.1 | BaodingNO.Du | China | Duck | 2019 | I |
| 28 | MK814581.1 | GX190510 | China | Duck | 2019 | I |
| 29 | MT084134.1 | SDHX1223 | China | Duck | 2018 | I |
| 30 | MT084131.1 | AHHF1109 | China | Duck | 2018 | I |
| 31 | MN928795.1 | HN02 | China | Duck | 2018 | I |
| 32 | MN068356.1 | HZ170301 | China | Duck | 2017 | I |
| 33 | MN068357.1 | YF180401 | China | Duck | 2017 | I |
| 34 | MF627688.1 | JSPX03E | China | Duck | 2016 | I |
| 35 | KY328304.1 | SDFC12 | China | Duck | 2015 | I |
| 36 | GU014543.1 | AQ0901 | China | Duck | 2009 | I |
| 37 | KC460531.1 | NN12/2012 | China | Duck | 2012 | I |
| 38 | HQ180265.1 | LZ/11/09 | China | Muscovy duck | 2009 | I |
| 39 | GU168779.1 | FJzq290 | China | Muscovy duck | 2008 | I |
| 40 | GQ334371.1 | Zhejiang | China | Duck | 2008 | I |
| 41 | EU344803.1 | LJ33 | China | Duck | 2007 | I |
| 42 | EU344806.1 | YS07 | China | Muscovy duck | 2007 | I |
| 43 | MT318126.1 | VC4 | Brasil | Mallard | 2014 | I |
| 44 | KC851819.1 | D12-KD-019 | South Korea | Duck | 2012 | I |
| 45 | KC851821.1 | D12-KD-028 | South Korea | Duck | 2012 | I |
| 46 | DQ100076.1 | 33753-52 | USA | Duck | 2007 | I |
| 47 | AY228555.1 | Germany | Mallard duck | 2003 | I | |
| 48 | NC_005053.1 | Germany | Mallard duck | 2003 | I | |
| 49 | OM176556.1 | VNUA-HD89 | Vietnam | Duck | 2021 | II |
| 50 | OM176557.1 | VNUA-BG135 | Vietnam | Duck | 2021 | II |
| 51 | MK814585.1 | YN180506 | China | Duck | 2018 | II |
| 52 | MN052853.1 | FJ1815 | China | Muscovy duck | 2018 | II |
| 53 | KR491945.1 | YN24-2013 | China | Muscovy duck | 2013 | II |
| 54 | KP229377.1 | CP12021 | Taiwan (China) | Muscovy duck | 2012 | II |
| 55 | GU131343.1 | WF0804 | China | Duck | 2008 | II |
| 56 | EU344805.1 | MH11 | China | Muscovy duck | 2007 | II |
| 57 | EU499309.1 | MH02/07 | China | Mule duck | 2007 | II |
| 58 | EU344802.1 | HZ09 | China | Muscovy duck | 2007 | II |
| 59 | DQ166836.1 | TC2 | Taiwan (China) | Muscovy duck | 2002 | II |
| 60 | DQ166838.1 | TC4 | Taiwan (China) | Muscovy duck | 2002 | II |
| 61 | OP432310.1 | HNU-HYH | China | Duck | 2022 | III |
| Farm Number | Location | No. of Tested Samples | No. of Positive Samples | Positive Rate |
|---|---|---|---|---|
| 1 | Thainguyen | 5 | 1 | 20 |
| 2 | Bacgiang | 5 | 2 | 40 |
| 3 | Haiduong | 5 | 2 | 40 |
| 4 | Bacgiang | 5 | 2 | 40 |
| 5 | Nghean | 5 | 1 | 20 |
| 6 | Thainguyen | 5 | 1 | 20 |
| 7 | Phutho | 5 | 3 | 60 |
| 8 | Hanoi | 5 | 3 | 60 |
| 9 | Hanam | 5 | 1 | 20 |
| Total | 45 | 16 | 35.56 | |
| Virus Strain | Nucleotide Identity (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Vietnam/ VNUA-102/2023 | Vietnam/ VNUA-114/2023 | Vietnam/ VNUA-137/2023 | Vietnam/ VNUA-225/2023 | Vietnam/ VNUA-251/2023 | Vietnam/ VNUA-318/2024 | Vietnam/ VNUA-322/2024 | Vietnam/ VNUA-315/2024 | Vietnam/ VNUA-331/2024 | |
| Vietnam/VNUA-102/2023 | 100 | ||||||||
| Vietnam/VNUA-114/2023 | 97.03 | 100 | |||||||
| Vietnam/VNUA-137/2023 | 97.03 | 99.19 | 100 | ||||||
| Vietnam/VNUA-225/2023 | 99.44 | 97.18 | 97.08 | 100 | |||||
| Vietnam/VNUA-251/2023 | 96.93 | 99.19 | 99.49 | 97.08 | 100 | ||||
| Vietnam/VNUA-318/2024 | 99.39 | 96.93 | 97.03 | 99.24 | 96.88 | 100 | |||
| Vietnam/VNUA-322/2024 | 97.13 | 99.29 | 99.19 | 97.29 | 99.29 | 97.08 | 100 | ||
| Vietnam/VNUA-315/2024 | 97.08 | 99.24 | 99.84 | 97.13 | 99.54 | 96.98 | 99.24 | 100 | |
| Vietnam/VNUA-331/2024 | 97.29 | 99.34 | 99.44 | 97.34 | 99.44 | 97.19 | 99.34 | 99.49 | 100 |
| Virus Strain | Nucleotide Identity (%) | |||||
|---|---|---|---|---|---|---|
| Vietnam/OM176553.1/ VNUA-HN47/2021 | Vietnam/OM176554.1/ VNUA-TB61/2021 | Vietnam/OM176552.1/ VNUA-HY40/2021 | Vietnam/OM176555.1/ VNUA-TN85/2021 | Vietnam/OM176556.1/ VNUA-HD89/2021 | Vietnam/OM176557.1/ VNUA-BG135/2021 | |
| Vietnam/VNUA-102/2023 | 96.98 | |||||
| Vietnam/VNUA-114/2023 | 99.14 | 99.04 | ||||
| Vietnam/VNUA-137/2023 | 99.54 | 99.44 | ||||
| Vietnam/VNUA-225/2023 | 97.03 | 97.03 | 97.04 | |||
| Vietnam/VNUA-251/2023 | 99.34 | 99.24 | 97.54 | 96.98 | ||
| Vietnam/VNUA-318/2024 | 96.88 | 96.88 | 97.19 | 99.79 | 83.50 | |
| Vietnam/VNUA-322/2024 | 99.14 | 99.04 | 97.64 | 97.19 | 83.70 | 84.44 |
| Vietnam/VNUA-315/2024 | 99.59 | 99.49 | 97.49 | 97.08 | 83.40 | 84.04 |
| Vietnam/VNUA-331/2024 | 99.39 | 99.29 | 97.79 | 97.29 | 83.65 | 84.19 |
| Strain Name | Virus with the Highest Nucleotide Identity | ||||
|---|---|---|---|---|---|
| GenBank Accession Number | Strain Name | Country | Year | % | |
| Vietnam/VNUA-102/2023 | OK094642.1 | DY01 | China | 2020 | 99.54 |
| Vietnam/VNUA-114/2023 | OR134472.1 | GX25 | China | 2020 | 99.49 |
| Vietnam/VNUA-137/2023 | OR134489.1 | GX51 | China | 2022 | 99.64 |
| OR842553.1 | CN/GD/1092 | China | 2023 | ||
| Vietnam/VNUA-225/2023 | OK094642.1 | DY01 | China | 2020 | 99.24 |
| Vietnam/VNUA-251/2023 | OR134464.1 | GX05 | China | 2019 | 99.59 |
| OR134467.1 | GX11 | China | 2019 | ||
| Vietnam/VNUA-318/2024 | OR134510.1 | GX39 | China | 2021 | 99.64 |
| OK094642.1 | DY01 | China | 2020 | ||
| Vietnam/VNUA-322/2024 | OR134472.1 | GX25 | China | 2020 | 99.49 |
| Vietnam/VNUA-315/2024 | OR134489.1 | GX51 | China | 2022 | 99.69 |
| Vietnam/VNUA-331/2024 | OR134472.1 | GX25 | China | 2020 | 99.74 |
| Virus Strain | Genotype | Amino Acid Residues on Cap Protein | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 12 | 47 | 55 | 82 | 106 | 107 | 155 | 156 | 177 | 183 | 194 | 197 | 205 | 236 | ||
| Majority | G | N | S | Q | N | K | T | T | V | V | T | Y | R | D | |
| VNUA-102/2023 | I | . | H | . | R | S | T | . | A | . | I | G | H | K | N |
| VNUA-114/2023 | I | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| VNUA-137/2023 | I | . | . | N | . | . | . | S | . | . | . | . | . | . | . |
| VNUA-225/2023 | I | . | H | S | R | S | T | . | A | . | I | G | H | K | N |
| VNUA-251/2023 | I | . | . | N | . | S | . | . | . | . | . | . | . | ||
| VNUA-318/2024 | I | . | H | . | R | S | T | . | . | . | I | G | H | N | |
| VNUA-322/2024 | I | . | . | . | . | . | . | . | . | I | . | . | . | . | . |
| VNUA-315/2024 | I | . | . | N | . | . | . | S | . | . | . | . | . | . | . |
| VNUA-331/2024 | I | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| VNUA-HN47/2021 | I | . | . | N | . | . | . | . | . | . | . | . | . | . | . |
| VNUA-TB61/2021 | I | S | . | N | . | . | . | . | . | . | . | . | . | . | . |
| VNUA-HY40/2021 | I | . | . | . | . | S | T | . | . | . | . | . | . | . | . |
| VNUA-TN85/2021 | I | . | H | . | R | S | T | . | . | . | I | G | H | K | N |
| VNUA-HD89/2021 | II | A | H | . | . | . | . | . | . | I | . | G | . | K | E |
| VNUA-BG135/2021 | II | A | H | . | . | G | . | . | . | I | . | G | . | K | E |
| Method | Recombination p-Value |
|---|---|
| GENECONV | 3.71 × 10−2 |
| MaxChi | 3.26 × 10−2 |
| Chimaera | 3.16 × 10−2 |
| SiScan | 9.58 × 10−5 |
| PhylPro | 3.42 × 10−2 |
| No. | Recombination Event | Major Parent | Minor Parent |
|---|---|---|---|
| 1 | Vietnam/VNUA-102/2023 | Vietnam/VNUA-114/2023 | Germany/NC_005053.1/2003 |
| 2 | Vietnam/VNUA-225/2023 | Vietnam/OM176552.1/VNUA-HY40/2021 | Germany/NC_005053.1/2003 |
| 3 | Vietnam/VNUA-318/2024 | Vietnam/OM176552.1/VNUA-HY40/2021 | Germany/NC_005053.1/2003 |
| Site | α | β | β − α | Prob [α > β] | Prob [α < β] | Bayes Factor [α and β] |
|---|---|---|---|---|---|---|
| 112 | 0.62 | 4.98 | 4.36 | 0.02 | 0.95 | 42.93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, H.V.; Trinh, D.Q.; Tran, G.H.T.; Vu, T.T.; Nguyen, T.H.B.; Rattanasrisomporn, A.; Bui, D.A.T.; Rattanasrisomporn, J. Characterization of an Emerging Recombinant Duck Circovirus in Northern Vietnam, 2023–2024. Viruses 2025, 17, 732. https://doi.org/10.3390/v17050732
Dong HV, Trinh DQ, Tran GHT, Vu TT, Nguyen THB, Rattanasrisomporn A, Bui DAT, Rattanasrisomporn J. Characterization of an Emerging Recombinant Duck Circovirus in Northern Vietnam, 2023–2024. Viruses. 2025; 17(5):732. https://doi.org/10.3390/v17050732
Chicago/Turabian StyleDong, Hieu Van, Dai Quang Trinh, Giang Huong Thi Tran, Thanh Thi Vu, Thinh Hung Ba Nguyen, Amonpun Rattanasrisomporn, Dao Anh Tran Bui, and Jatuporn Rattanasrisomporn. 2025. "Characterization of an Emerging Recombinant Duck Circovirus in Northern Vietnam, 2023–2024" Viruses 17, no. 5: 732. https://doi.org/10.3390/v17050732
APA StyleDong, H. V., Trinh, D. Q., Tran, G. H. T., Vu, T. T., Nguyen, T. H. B., Rattanasrisomporn, A., Bui, D. A. T., & Rattanasrisomporn, J. (2025). Characterization of an Emerging Recombinant Duck Circovirus in Northern Vietnam, 2023–2024. Viruses, 17(5), 732. https://doi.org/10.3390/v17050732