
Impact of Vaccine-Elicited Anti-Spike IgG4 Antibodies on Fc-Effector Functions Against SARS-CoV-2

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Human Subjects
2.3. Plasma Samples and Antibodies
2.4. Plasmids
2.5. Protein Expression and Purification
2.6. Cell Lines
2.7. Anti-Nucleocapsid (N) Assay
2.8. Anti-S ELISA
2.9. Cell-Surface Staining and Flow Cytometry Analysis
2.10. Antibody-Dependent Cellular Cytotoxicity Assay
2.11. Antibody-Dependent Cellular Phagocytosis Assay
2.12. Pseudovirus Neutralization Assay
2.13. Statistical Analysis
3. Results
3.1. Longitudinal Humoral Responses Elicited by mRNA Vaccination in a Cohort of Health Care Workers
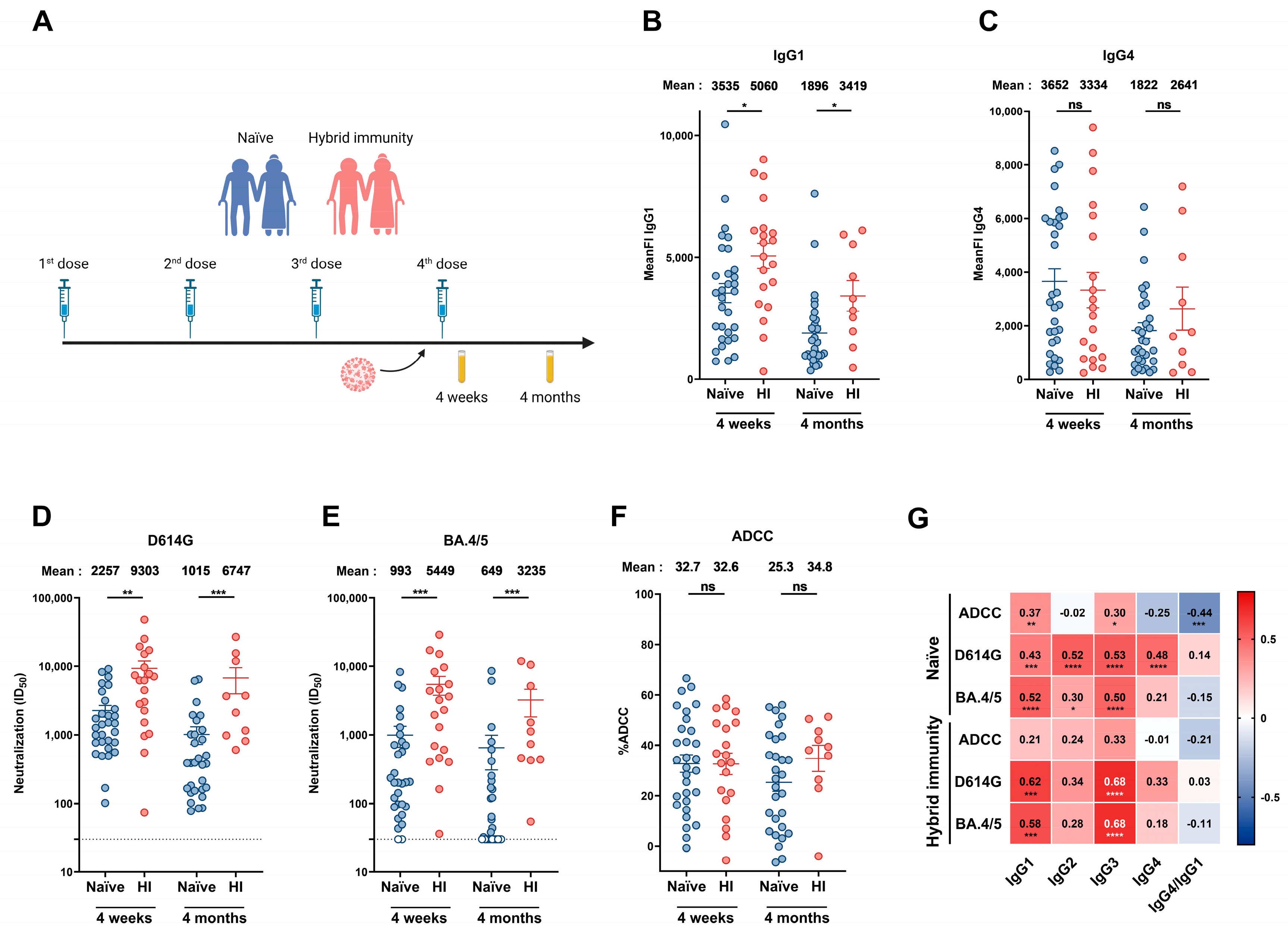
3.2. Humoral Responses Elicited in Donors Aged over 70 Years
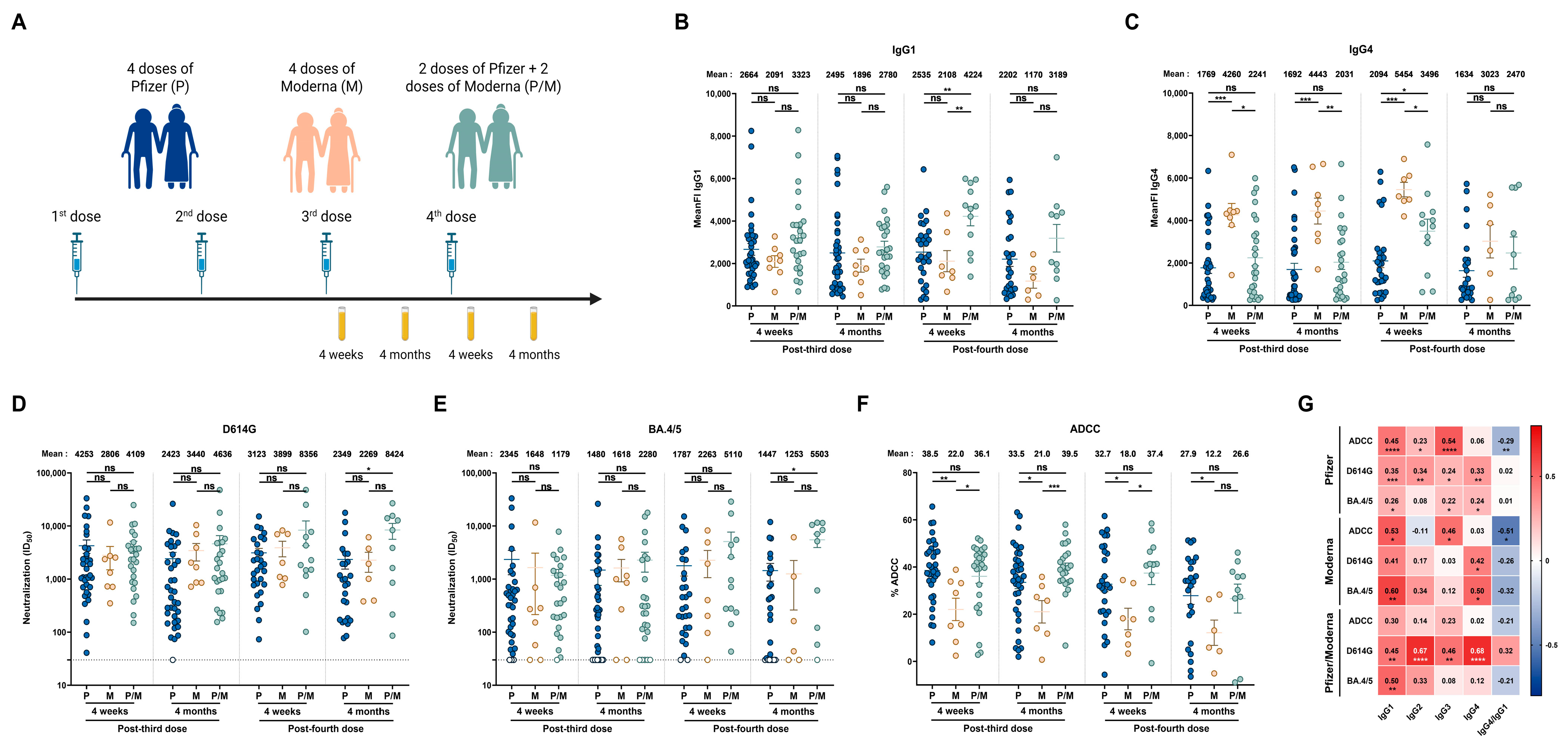
3.3. Impact of Vaccine Platform on Humoral Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Yue, H.; Bai, X.; Wang, J.; Yu, Q.; Liu, W.; Pu, J.; Wang, X.; Hu, J.; Xu, D.; Li, X.; et al. Clinical characteristics of coronavirus disease 2019 in Gansu province, China. Ann. Palliat. Med. 2020, 9, 1404–1412. [Google Scholar] [CrossRef]
- WHO. WHO Coronavirus (COVID-19) Dashboard; WHO: Geneva, Switzerland, 2024. [Google Scholar]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Thomas, S.J.; Moreira, E.D., Jr.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Polack, F.P.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine through 6 Months. N. Engl. J. Med. 2021, 385, 1761–1773. [Google Scholar] [CrossRef] [PubMed]
- Markov, P.V.; Ghafari, M.; Beer, M.; Lythgoe, K.; Simmonds, P.; Stilianakis, N.I.; Katzourakis, A. The evolution of SARS-CoV-2. Nat. Rev. Microbiol. 2023, 21, 361–379. [Google Scholar] [CrossRef]
- Yang, S.; Yu, Y.; Xu, Y.; Jian, F.; Song, W.; Yisimayi, A.; Wang, P.; Wang, J.; Liu, J.; Yu, L.; et al. Fast evolution of SARS-CoV-2 BA.2.86 to JN.1 under heavy immune pressure. Lancet Infect. Dis. 2024, 24, e70–e72. [Google Scholar] [CrossRef]
- Planas, D.; Staropoli, I.; Michel, V.; Lemoine, F.; Donati, F.; Prot, M.; Porrot, F.; Guivel-Benhassine, F.; Jeyarajah, B.; Brisebarre, A.; et al. Distinct evolution of SARS-CoV-2 Omicron XBB and BA.2.86/JN.1 lineages combining increased fitness and antibody evasion. Nat. Commun. 2024, 15, 2254. [Google Scholar] [CrossRef]
- Tauzin, A.; Benlarbi, M.; Medjahed, H.; Gregoire, Y.; Perreault, J.; Gendron-Lepage, G.; Gokool, L.; Morrisseau, C.; Arlotto, P.; Tremblay, C.; et al. Humoral Responses against BQ.1.1 Elicited after Breakthrough Infection and SARS-CoV-2 mRNA Vaccination. Vaccines 2023, 11, 242. [Google Scholar] [CrossRef]
- INSPQ. Les Variants du SRAS-CoV-2; INSPQ: Québec, QC, Canada, 2025. Available online: https://www.inspq.qc.ca/covid-19/donnees/variants (accessed on 3 December 2024).
- Tauzin, A.; Nicolas, A.; Ding, S.; Benlarbi, M.; Medjahed, H.; Chatterjee, D.; Dionne, K.; Gong, S.Y.; Gendron-Lepage, G.; Bo, Y.; et al. Spike recognition and neutralization of SARS-CoV-2 Omicron subvariants elicited after the third dose of mRNA vaccine. Cell Rep. 2023, 42, 111998. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Kurhade, C.; Patel, S.; Kitchin, N.; Tompkins, K.; Cutler, M.; Cooper, D.; Yang, Q.; Cai, H.; Muik, A.; et al. Neutralization of BA.4-BA.5, BA.4.6, BA.2.75.2, BQ.1.1, and XBB.1 with Bivalent Vaccine. N. Engl. J. Med. 2023, 388, 854–857. [Google Scholar] [CrossRef]
- Wang, Q.; Bowen, A.; Tam, A.R.; Valdez, R.; Stoneman, E.; Mellis, I.A.; Gordon, A.; Liu, L.; Ho, D.D. SARS-CoV-2 neutralising antibodies after bivalent versus monovalent booster. Lancet Infect. Dis. 2023, 23, 527–528. [Google Scholar] [CrossRef] [PubMed]
- Benlarbi, M.; Ding, S.; Belanger, E.; Tauzin, A.; Poujol, R.; Medjahed, H.; El Ferri, O.; Bo, Y.; Bourassa, C.; Hussin, J.; et al. Temperature-dependent Spike-ACE2 interaction of Omicron subvariants is associated with viral transmission. mBio 2024, 15, e0090724. [Google Scholar] [CrossRef]
- Anand, S.P.; Prevost, J.; Nayrac, M.; Beaudoin-Bussieres, G.; Benlarbi, M.; Gasser, R.; Brassard, N.; Laumaea, A.; Gong, S.Y.; Bourassa, C.; et al. Longitudinal analysis of humoral immunity against SARS-CoV-2 Spike in convalescent individuals up to 8 months post-symptom onset. Cell Rep. Med. 2021, 2, 100290. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.; Gendron-Lepage, G.; Nayrac, M.; Anand, S.P.; Bourassa, C.; Medjahed, H.; Goyette, G.; Dube, M.; Bazin, R.; Kaufmann, D.E.; et al. Evolution of Anti-RBD IgG Avidity following SARS-CoV-2 Infection. Viruses 2022, 14, 532. [Google Scholar] [CrossRef]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG subclasses and allotypes: From structure to effector functions. Front. Immunol. 2014, 5, 520. [Google Scholar] [CrossRef]
- Damelang, T.; Rogerson, S.J.; Kent, S.J.; Chung, A.W. Role of IgG3 in Infectious Diseases. Trends Immunol. 2019, 40, 197–211. [Google Scholar] [CrossRef]
- Rispens, T.; Huijbers, M.G. The unique properties of IgG4 and its roles in health and disease. Nat. Rev. Immunol. 2023, 23, 763–778. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N.; Redwan, E.M.; Makis, W.; Rubio-Casillas, A. IgG4 Antibodies Induced by Repeated Vaccination May Generate Immune Tolerance to the SARS-CoV-2 Spike Protein. Vaccines 2023, 11, 991. [Google Scholar] [CrossRef] [PubMed]
- Irrgang, P.; Gerling, J.; Kocher, K.; Lapuente, D.; Steininger, P.; Habenicht, K.; Wytopil, M.; Beileke, S.; Schafer, S.; Zhong, J.; et al. Class switch toward noninflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination. Sci. Immunol. 2023, 8, eade2798. [Google Scholar] [CrossRef]
- Gelderloos, A.T.; Verheul, M.K.; Middelhof, I.; de Zeeuw-Brouwer, M.L.; van Binnendijk, R.S.; Buisman, A.M.; van Kasteren, P.B. Repeated COVID-19 mRNA vaccination results in IgG4 class switching and decreased NK cell activation by S1-specific antibodies in older adults. Immun. Ageing 2024, 21, 63. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.; Beaudoin-Bussieres, G.; Benlarbi, M.; Nayrac, M.; Bo, Y.; Gendron-Lepage, G.; Medjahed, H.; Perreault, J.; Gokool, L.; Arlotto, P.; et al. Humoral Responses Elicited after a Fifth Dose of SARS-CoV-2 mRNA Bivalent Vaccine. Viruses 2023, 15, 1926. [Google Scholar] [CrossRef]
- Beaudoin-Bussieres, G.; Tauzin, A.; Dionne, K.; Gendron-Lepage, G.; Medjahed, H.; Perreault, J.; Levade, I.; Alfadhli, L.; Bo, Y.; Bazin, R.; et al. A Recent SARS-CoV-2 Infection Enhances Antibody-Dependent Cellular Cytotoxicity against Several Omicron Subvariants following a Fourth mRNA Vaccine Dose. Viruses 2023, 15, 1274. [Google Scholar] [CrossRef]
- Germain, M.; Lewin, A.; Bazin, R.; Dieude, M.; Perreault, J.; Boivin, A.; Gregoire, Y.; Renaud, C. Cohort profile: A Quebec-based plasma donor biobank to study COVID-19 immunity (PlasCoV). BMJ Open 2023, 13, e068803. [Google Scholar] [CrossRef]
- Li, W.; Chen, Y.; Prevost, J.; Ullah, I.; Lu, M.; Gong, S.Y.; Tauzin, A.; Gasser, R.; Vezina, D.; Anand, S.P.; et al. Structural basis and mode of action for two broadly neutralizing antibodies against SARS-CoV-2 emerging variants of concern. Cell Rep. 2022, 38, 110210. [Google Scholar] [CrossRef]
- Beaudoin-Bussieres, G.; Chen, Y.; Ullah, I.; Prevost, J.; Tolbert, W.D.; Symmes, K.; Ding, S.; Benlarbi, M.; Gong, S.Y.; Tauzin, A.; et al. A Fc-enhanced NTD-binding non-neutralizing antibody delays virus spread and synergizes with a nAb to protect mice from lethal SARS-CoV-2 infection. Cell Rep. 2022, 38, 110368. [Google Scholar] [CrossRef]
- Jennewein, M.F.; MacCamy, A.J.; Akins, N.R.; Feng, J.; Homad, L.J.; Hurlburt, N.K.; Seydoux, E.; Wan, Y.H.; Stuart, A.B.; Edara, V.V.; et al. Isolation and characterization of cross-neutralizing coronavirus antibodies from COVID-19+ subjects. Cell Rep. 2021, 36, 109353. [Google Scholar] [CrossRef]
- Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.D.; So, R.T.Y.; Lv, H.; Mok, C.K.P.; Wilson, I.A. A highly conserved cryptic epitope in the receptor binding domains of SARS-CoV-2 and SARS-CoV. Science 2020, 368, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Prevost, J.; Ladinsky, M.S.; Stone, H.; Lu, M.; Anand, S.P.; Beaudoin-Bussieres, G.; Symmes, K.; Benlarbi, M.; Ding, S.; et al. Live imaging of SARS-CoV-2 infection in mice reveals that neutralizing antibodies require Fc function for optimal efficacy. Immunity 2021, 54, 2143–2158.e15. [Google Scholar] [CrossRef]
- Hsieh, C.L.; Goldsmith, J.A.; Schaub, J.M.; DiVenere, A.M.; Kuo, H.C.; Javanmardi, K.; Le, K.C.; Wrapp, D.; Lee, A.G.; Liu, Y.; et al. Structure-based design of prefusion-stabilized SARS-CoV-2 spikes. Science 2020, 369, 1501–1505. [Google Scholar] [CrossRef] [PubMed]
- Prevost, J.; Gasser, R.; Beaudoin-Bussieres, G.; Richard, J.; Duerr, R.; Laumaea, A.; Anand, S.P.; Goyette, G.; Benlarbi, M.; Ding, S.; et al. Cross-Sectional Evaluation of Humoral Responses against SARS-CoV-2 Spike. Cell Rep. Med. 2020, 1, 100126. [Google Scholar] [CrossRef]
- Beaudoin-Bussieres, G.; Richard, J.; Prevost, J.; Goyette, G.; Finzi, A. A new flow cytometry assay to measure antibody-dependent cellular cytotoxicity against SARS-CoV-2 Spike-expressing cells. STAR Protoc. 2021, 2, 100851. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.; Motoi, S.; Sugiura, M.; Kajikawa, M.; Kojima, S.; Kohroki, J.; Masuho, Y. Antibody-dependent cellular cytotoxicity and cytokine/chemokine secretion by KHYG-1 cells stably expressing FcgammaRIIIA. Immunol. Lett. 2014, 161, 59–64. [Google Scholar] [CrossRef]
- Benlarbi, M.; Kenfack, D.D.; Dionne, K.; Cote-Chenette, M.; Beaudoin-Bussieres, G.; Belanger, E.; Ding, S.; Goni, O.H.; Ngoume, Y.F.; Tauzin, A.; et al. Longitudinal humoral immunity against SARS-CoV-2 Spike following infection in individuals from Cameroon. Virology 2025, 605, 110467. [Google Scholar] [CrossRef]
- Bredholt, G.; Saevik, M.; Soyland, H.; Ueland, T.; Zhou, F.; Pathirana, R.; Madsen, A.; Vahokoski, J.; Lartey, S.; Halvorsen, B.E.; et al. Three doses of Sars-CoV-2 mRNA vaccine in older adults result in similar antibody responses but reduced cellular cytokine responses relative to younger adults. Vaccine X 2024, 20, 100564. [Google Scholar] [CrossRef]
- Jergovic, M.; Uhrlaub, J.L.; Watanabe, M.; Bradshaw, C.M.; White, L.M.; LaFleur, B.J.; Edwards, T.; Sprissler, R.; Worobey, M.; Bhattacharya, D.; et al. Competent immune responses to SARS-CoV-2 variants in older adults following two doses of mRNA vaccination. Nat. Commun. 2022, 13, 2891. [Google Scholar] [CrossRef]
- van den Hoogen, L.L.; Boer, M.; Postema, A.; de Rond, L.; de Zeeuw-Brouwer, M.L.; Pronk, I.; Wijmenga-Monsuur, A.J.; Bijvank, E.; Kruiper, C.; Beckers, L.; et al. Reduced Antibody Acquisition with Increasing Age following Vaccination with BNT162b2: Results from Two Longitudinal Cohort Studies in The Netherlands. Vaccines 2022, 10, 1480. [Google Scholar] [CrossRef]
- Hoeve, C.E.; Huiberts, A.J.; de Gier, B.; Andeweg, S.P.; den Hartog, G.; de Melker, H.E.; Hahne, S.J.M.; van de Wijgert, J.; van den Hof, S.; Knol, M.J. COVID-19 vaccination-induced antibody responses and waning by age and comorbidity status in a large population-based prospective cohort study. Vaccine 2024, 42, 126121. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin-Bussieres, G.; Tauzin, A.; Dionne, K.; El Ferri, O.; Benlarbi, M.; Bourassa, C.; Medjahed, H.; Bazin, R.; Cote, M.; Finzi, A. Multiple exposures to SARS-CoV-2 Spike enhance cross-reactive antibody-dependent cellular cytotoxicity against SARS-CoV-1. Virology 2025, 607, 110512. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Beaudoin-Bussieres, G.; Symmes, K.; Cloutier, M.; Ducas, E.; Tauzin, A.; Laumaea, A.; Grunst, M.W.; Dionne, K.; Richard, J.; et al. The Fc-effector function of COVID-19 convalescent plasma contributes to SARS-CoV-2 treatment efficacy in mice. Cell Rep. Med. 2023, 4, 100893. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Health Care Workers | |||
|---|---|---|---|
| No Recent BTI | Recent BTI | ||
| Number (n) a | 8 | 6 | |
| Age b | 53.5 (31–69) | 58 (51–62) | |
| Sex a | Female (n) | 7 | 2 |
| Male (n) | 1 | 4 | |
| First dose (n) a | Pfizer monovalent | 5 | 6 |
| Moderna monovalent | 3 | 0 | |
| Days between the first and second dose b | 97.5 | 111.5 | |
| Second dose (n) a | Pfizer monovalent | 5 | 6 |
| Moderna monovalent | 3 | 0 | |
| Days between the second and third dose b | 203 | 218 | |
| Third dose (n) a | Pfizer monovalent | 6 | 5 |
| Moderna monovalent | 2 | 0 | |
| NA | 0 | 1 | |
| Days between the third and fourth dose b | 195.5 | 186.5 | |
| Fourth dose (n) a | Pfizer monovalent | 8 | 5 |
| Moderna monovalent | 0 | 0 | |
| Moderna bivalent BA.1 | 0 | 1 | |
| Days between the fourth and fifth dose b | 111 | 120 | |
| Fifth dose (n) a | Pfizer monovalent | 0 | 1 |
| Moderna bivalent BA.1 | 2 | 1 | |
| Pfizer bivalent BA.4/5 | 4 | 3 | |
| NA | 1 | 1 | |
| Days between the fifth and sixth dose b | 350 | 378 | |
| Sixth dose (n) a | Pfizer monovalent | 1 | 0 |
| Pfizer bivalent BA.4/5 | 2 | 1 | |
| Moderna bivalent BA.1 | 1 | 0 | |
| Pfizer monovalent XBB.1.5 | 1 | 3 | |
| Moderna monovalent XBB.1.5 | 2 | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dionne, K.; Tauzin, A.; Bélanger, É.; Desfossés, Y.; Benlarbi, M.; Niu, L.; Beaudoin-Bussières, G.; Medjahed, H.; Bourassa, C.; Perreault, J.; et al. Impact of Vaccine-Elicited Anti-Spike IgG4 Antibodies on Fc-Effector Functions Against SARS-CoV-2. Viruses 2025, 17, 666. https://doi.org/10.3390/v17050666
Dionne K, Tauzin A, Bélanger É, Desfossés Y, Benlarbi M, Niu L, Beaudoin-Bussières G, Medjahed H, Bourassa C, Perreault J, et al. Impact of Vaccine-Elicited Anti-Spike IgG4 Antibodies on Fc-Effector Functions Against SARS-CoV-2. Viruses. 2025; 17(5):666. https://doi.org/10.3390/v17050666
Chicago/Turabian StyleDionne, Katrina, Alexandra Tauzin, Étienne Bélanger, Yann Desfossés, Mehdi Benlarbi, Ling Niu, Guillaume Beaudoin-Bussières, Halima Medjahed, Catherine Bourassa, Josée Perreault, and et al. 2025. "Impact of Vaccine-Elicited Anti-Spike IgG4 Antibodies on Fc-Effector Functions Against SARS-CoV-2" Viruses 17, no. 5: 666. https://doi.org/10.3390/v17050666
APA StyleDionne, K., Tauzin, A., Bélanger, É., Desfossés, Y., Benlarbi, M., Niu, L., Beaudoin-Bussières, G., Medjahed, H., Bourassa, C., Perreault, J., Pazgier, M., Bazin, R., & Finzi, A. (2025). Impact of Vaccine-Elicited Anti-Spike IgG4 Antibodies on Fc-Effector Functions Against SARS-CoV-2. Viruses, 17(5), 666. https://doi.org/10.3390/v17050666