
The Emerging Role of Circulating T Follicular Helper Cells in Dengue Virus Immunity: Balancing Protection and Pathogenesis

 , and
, and {kind=link}
{kind=link}
Abstract
1. Dengue Virus Overview
2. Dengue Clinical Manifestations
3. Contribution of CD8+ T Cells to the Immune Response Against Dengue Virus
4. CD4+ T Cells and Their Contribution to Dengue Virus Immunity
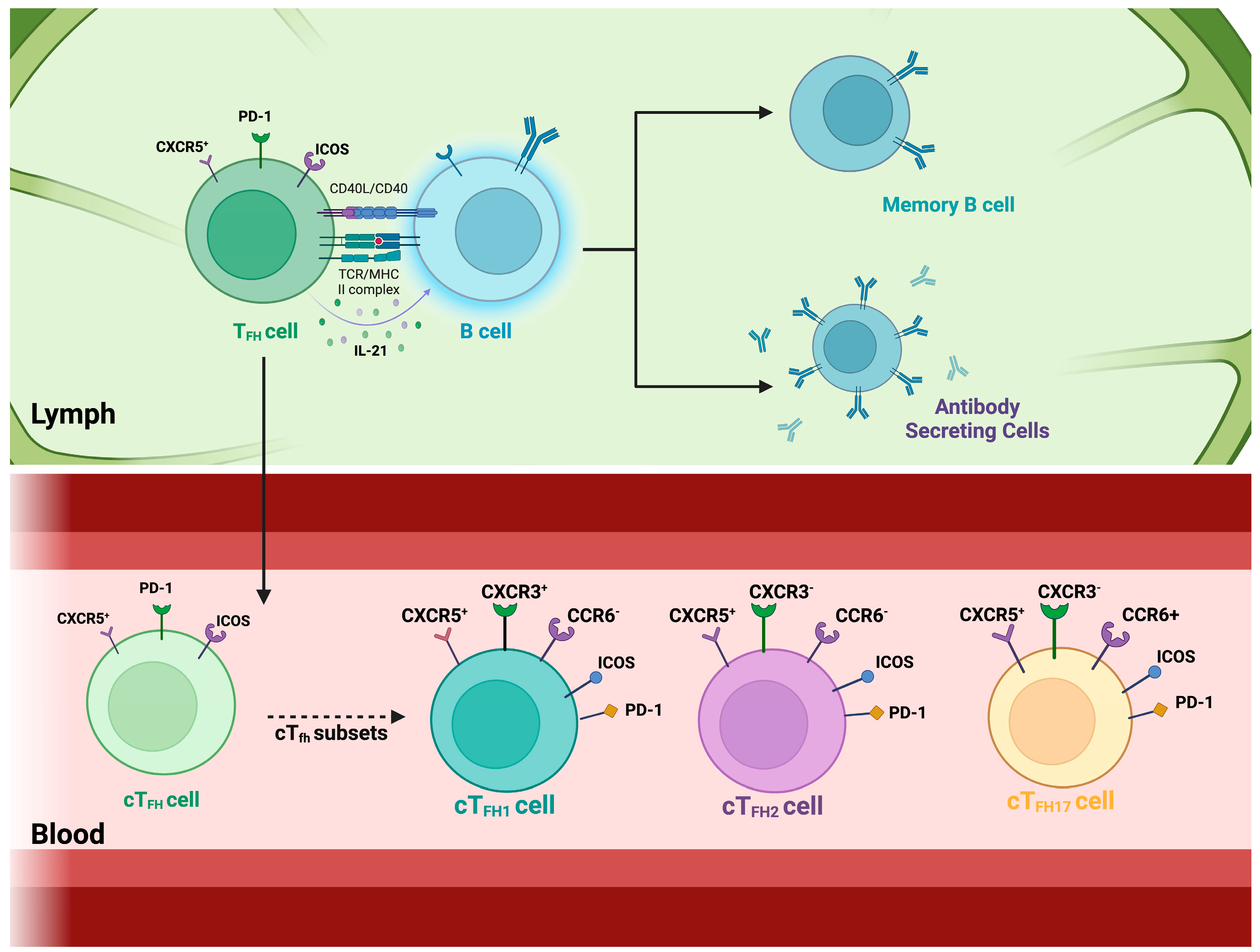
5. The Role of Circulatory T Follicular Helper Cells
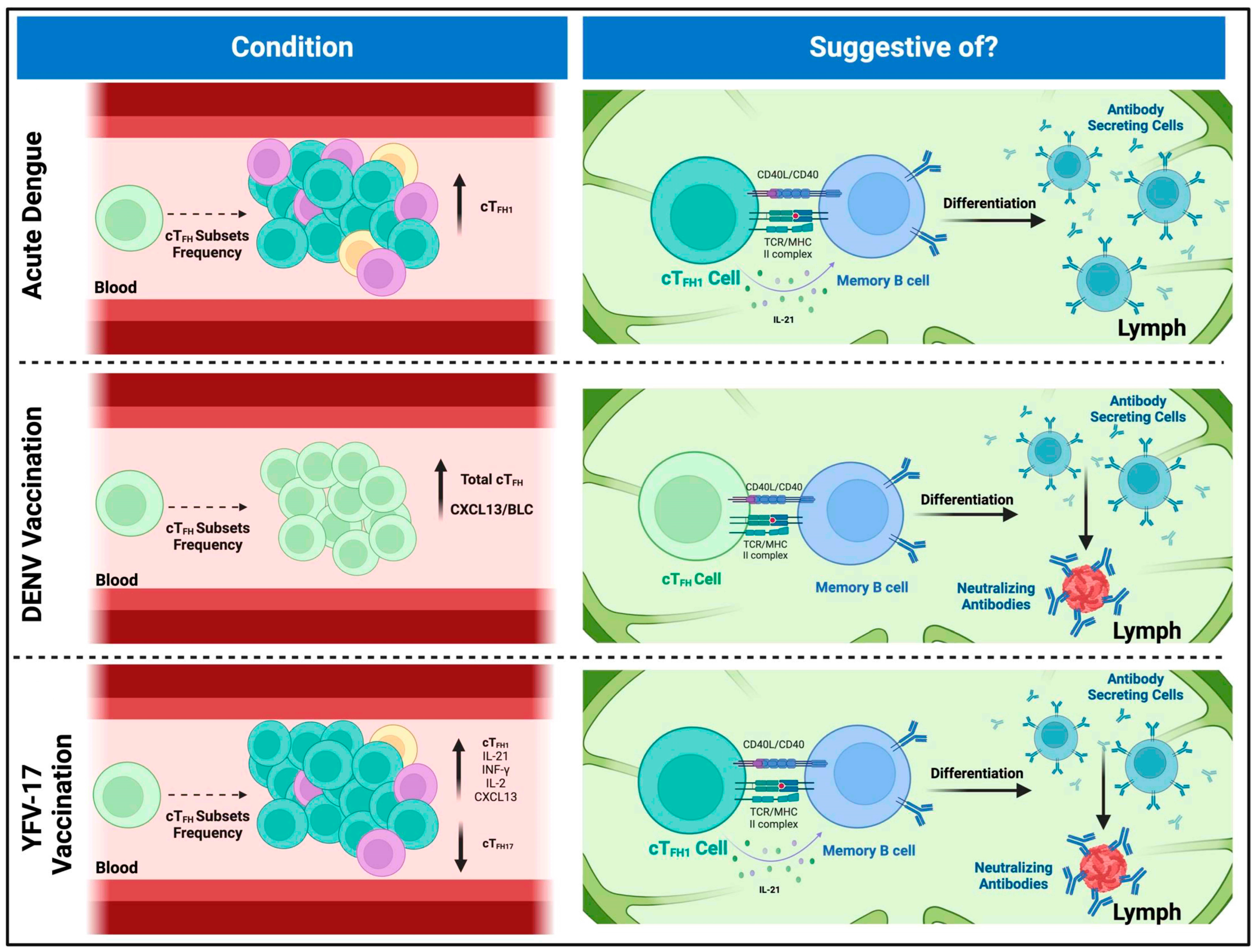
6. Characterizing Circulating T Follicular Helper Cells in Flavivirus-Induced Immune Responses
7. T Cell Function in Dengue Immune Response: Implications and Challenges for Vaccine Development
8. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holbrook, M.R. Historical Perspectives on Flavivirus Research. Viruses 2017, 9, 97. [Google Scholar] [CrossRef] [PubMed]
- Blaney, J.E.; Durbin, A.P.; Murphy, B.R.; Whitehead, S.S. Development of a live attenuated dengue virus vaccine using reverse genetics. Viral Immunol. 2006, 19, 10–32. [Google Scholar] [CrossRef] [PubMed]
- Nasar, S.; Rashid, N.; Iftikhar, S. Dengue proteins with their role in pathogenesis, and strategies for developing an effective anti-dengue treatment: A review. J. Med. Virol. 2020, 92, 941–955. [Google Scholar] [CrossRef]
- Rothman, A.L. Immunity to dengue virus: A tale of original antigenic sin and tropical cytokine storms. Nat. Rev. Immunol. 2011, 11, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Education, N. Dengue Viruses. Available online: https://www.nature.com/scitable/topicpage/dengue-viruses-22400925/#:~:text=The%20Dengue%20Serotypes&text=These%20four%20viruses%20are%20called,there%20is%20some%20genetic%20variation (accessed on 15 February 2024).
- Zerfu, B.; Kassa, T.; Legesse, M. Epidemiology, biology, pathogenesis, clinical manifestations, and diagnosis of dengue virus infection, and its trend in Ethiopia: A comprehensive literature review. Trop. Med. Health 2023, 51, 11. [Google Scholar] [CrossRef] [PubMed]
- Chng, M.H.Y.; Lim, M.Q.; Rouers, A.; Becht, E.; Lee, B.; MacAry, P.A.; Lye, D.C.; Leo, Y.S.; Chen, J.; Fink, K.; et al. Large-Scale HLA Tetramer Tracking of T Cells during Dengue Infection Reveals Broad Acute Activation and Differentiation into Two Memory Cell Fates. Immunity 2019, 51, 1119–1135. [Google Scholar] [CrossRef]
- Guzman, M.G.; Gubler, D.J.; Izquierdo, A.; Martinez, E.; Halstead, S.B. Dengue infection. Nat. Rev. Dis. Primers 2016, 2, 16055. [Google Scholar] [CrossRef]
- Khan, M.B.; Yang, Z.S.; Lin, C.Y.; Hsu, M.C.; Urbina, A.N.; Assavalapsakul, W.; Wang, W.H.; Chen, Y.H.; Wang, S.F. Dengue overview: An updated systemic review. J. Infect. Public Health 2023, 16, 1625–1642. [Google Scholar] [CrossRef]
- Izmirly, A.M.; Alturki, S.O.; Alturki, S.O.; Connors, J.; Haddad, E.K. Challenges in Dengue Vaccines Development: Pre-existing Infections and Cross-Reactivity. Front. Immunol. 2020, 11, 1055. [Google Scholar] [CrossRef] [PubMed]
- Huber, J.E.; Ahlfeld, J.; Scheck, M.K.; Zaucha, M.; Witter, K.; Lehmann, L.; Karimzadeh, H.; Pritsch, M.; Hoelscher, M.; von Sonnenburg, F.; et al. Dynamic changes in circulating T follicular helper cell composition predict neutralising antibody responses after yellow fever vaccination. Clin. Transl. Immunol. 2020, 9, e1129. [Google Scholar] [CrossRef] [PubMed]
- Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control: New Edition; WHO: Geneva, Switzerland, 2009.
- Kalayanarooj, S. Clinical Manifestations and Management of Dengue/DHF/DSS. Trop. Med. Health 2011, 39, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Trofa, A.F.; DeFraites, R.F.; Smoak, B.L.; Kanesa-thasan, N.; King, A.D.; Burrous, J.M.; MacArthy, P.O.; Rossi, C.; Hoke, C.H. Dengue fever in US military personnel in Haiti. JAMA 1997, 277, 1546–1548. [Google Scholar] [CrossRef] [PubMed]
- Ajlan, B.A.; Alafif, M.M.; Alawi, M.M.; Akbar, N.A.; Aldigs, E.K.; Madani, T.A. Assessment of the new World Health Organization’s dengue classification for predicting severity of illness and level of healthcare required. PLoS Negl. Trop. Dis. 2019, 13, e0007144. [Google Scholar] [CrossRef] [PubMed]
- Kalayanarooj, S.; Vaughn, D.W.; Nimmannitya, S.; Green, S.; Suntayakorn, S.; Kunentrasai, N.; Viramitrachai, W.; Ratanachu-eke, S.; Kiatpolpoj, S.; Innis, B.L.; et al. Early clinical and laboratory indicators of acute dengue illness. J. Infect. Dis. 1997, 176, 313–321. [Google Scholar] [CrossRef]
- Fukusumi, M.; Arashiro, T.; Arima, Y.; Matsui, T.; Shimada, T.; Kinoshita, H.; Arashiro, A.; Takasaki, T.; Sunagawa, T.; Oishi, K. Dengue Sentinel Traveler Surveillance: Monthly and Yearly Notification Trends among Japanese Travelers, 2006–2014. PLoS Negl. Trop. Dis. 2016, 10, e0004924. [Google Scholar] [CrossRef]
- Leder, K.; Torresi, J.; Brownstein, J.S.; Wilson, M.E.; Keystone, J.S.; Barnett, E.; Schwartz, E.; Schlagenhauf, P.; Wilder-Smith, A.; Castelli, F.; et al. Travel-associated illness trends and clusters, 2000–2010. Emerg. Infect. Dis. 2013, 19, 1049–1073. [Google Scholar] [CrossRef]
- Cao, X.T.; Ngo, T.N.; Wills, B.; Kneen, R.; Nguyen, T.T.; Ta, T.T.; Tran, T.T.; Doan, T.K.; Solomon, T.; Simpson, J.A.; et al. Evaluation of the World Health Organization standard tourniquet test and a modified tourniquet test in the diagnosis of dengue infection in Viet Nam. Trop. Med. Int. Health 2002, 7, 125–132. [Google Scholar]
- Wilder-Smith, A.; Schwartz, E. Dengue in travelers. N. Engl. J. Med. 2005, 353, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, B.D.C.; Correia, D. Ultrasound Assessment of Hepatobiliary and Splenic Changes in Patients with Dengue and Warning Signs During the Acute and Recovery Phases. J. Ultrasound Med. 2019, 38, 2015–2024. [Google Scholar] [CrossRef]
- Srikiatkhachorn, A.; Krautrachue, A.; Ratanaprakarn, W.; Wongtapradit, L.; Nithipanya, N.; Kalayanarooj, S.; Nisalak, A.; Thomas, S.J.; Gibbons, R.V.; Mammen, M.P.; et al. Natural history of plasma leakage in dengue hemorrhagic fever: A serial ultrasonographic study. Pediatr. Infect. Dis. J. 2007, 26, 283–290; discussion 291–292. [Google Scholar] [CrossRef]
- Rivino, L. T cell immunity to dengue virus and implications for vaccine design. Expert Rev. Vaccines 2016, 15, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Travers, P.; Walport, M.; Shlomchik, M.J. Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Weiskopf, D.; Angelo, A.M.; Azeredo, D.L.E.; Sidney, J.; Greenbaum, A.J.; Fernando, N.A.; Broadwater, A.; Kolla, V.R.; Silva, D.D.A.; Silva, D.M.A.; et al. Comprehensive analysis of dengue virus-specific responses supports an HLA-linked protective role for CD8+ T cells. Proc. Natl. Acad. Sci. USA 2013, 110, E2046–E2053. [Google Scholar] [CrossRef] [PubMed]
- Zellweger, M.R.L.; Shresta, S. Mouse Models to Study Dengue Virus Immunology and Pathogenesis. Front. Immunol. 2014, 5, 151. [Google Scholar]
- Rivino, L. Understanding the Human T Cell Response to Dengue Virus. In Advances in Experimental Medicine and Biology; Springer Nature: London, UK, 2018; Volume 1062, pp. 241–250. [Google Scholar]
- Ngono, E.A.; Shresta, S. Immune Response to Dengue and Zika. Annu. Rev. Immunol. 2018, 36, 279–308. [Google Scholar] [CrossRef]
- Fisher, G.R.; James, C.E., Jr.; Johnson, T.R.; Tang, Y.W.; Graham, B.S. Passive IgA Monoclonal Antibody Is No More Effective Than IgG at Protecting Mice from Mucosal Challenge with Respiratory Syncytial Virus. J. Infect. Dis. 1999, 180, 1324–1327. [Google Scholar] [CrossRef]
- Weiskopf, D.; Bangs, D.J.; Sidney, J.; Kolla, R.V.; Silva, D.D.A.; Silva, D.M.A.; Crotty, S.; Peters, B.; Sette, A. Dengue virus infection elicits highly polarized CX3CR1 + cytotoxic CD4 + T cells associated with protective immunity. Proc. Natl. Acad. Sci. USA 2015, 112, E4256–E4263. [Google Scholar]
- Grifoni, A.; Pham, J.; Sidney, J.; O’Rourke, H.P.; Paul, S.; Peters, B.; Martini, R.S.; Silva, D.D.A.; Ricciardi, J.M.; Magnani, M.D.; et al. Prior Dengue Virus Exposure Shapes T Cell Immunity to Zika Virus in Humans. J. Virol. 2017, 91, e01469-17. [Google Scholar] [CrossRef]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef]
- St John, A.L.; Rathore, A.P.S. Adaptive immune responses to primary and secondary dengue virus infections. Nat. Rev. Immunol. 2019, 19, 218–230. [Google Scholar]
- Stebegg, M.; Kumar, S.D.; Silva-Cayetano, A.; Fonseca, V.R.; Linterman, M.A.; Graca, L. Regulation of the Germinal Center Response. Front. Immunol. 2018, 9, 2469. [Google Scholar] [CrossRef] [PubMed]
- Jogdand, G.M.; Mohanty, S.; Devadas, S. Regulators of Tfh Cell Differentiation. Front. Immunol. 2016, 7, 520. [Google Scholar]
- Vinuesa, C.G.; Cyster, J.G. How T cells earn the follicular rite of passage. Immunity 2011, 35, 671–680. [Google Scholar] [PubMed]
- Vinuesa, C.G.; Linterman, M.A.; Yu, D.; MacLennan, I.C. Follicular Helper T Cells. Annu. Rev. Immunol. 2016, 34, 335–368. [Google Scholar]
- Crotty, S. T Follicular Helper Cell Biology: A Decade of Discovery and Diseases. Immunity 2019, 50, 1132–1148. [Google Scholar]
- Locci, M.; Havenar-Daughton, C.; Landais, E.; Wu, J.; Kroenke, M.A.; Arlehamn, C.L.; Su, L.F.; Cubas, R.; Davis, M.M.; Sette, A.; et al. Human circulating PD-1 + CXCR3-CXCR5 + memory Tfh cells are highly functional and correlate with broadly neutralizing HIV antibody responses. Immunity 2013, 39, 758–769. [Google Scholar] [PubMed]
- Haltaufderhyde, K.; Srikiatkhachorn, A.; Green, S.; Macareo, L.; Park, S.; Kalayanarooj, S.; Rothman, A.L.; Mathew, A. Activation of Peripheral T Follicular Helper Cells During Acute Dengue Virus Infection. J. Infect. Dis. 2018, 218, 1675–1685. [Google Scholar]
- Gong, F.; Zheng, T.; Zhou, P. T Follicular Helper Cell Subsets and the Associated Cytokine IL-21 in the Pathogenesis and Therapy of Asthma. Front. Immunol. 2019, 10, 2918. [Google Scholar]
- Bentebibel, S.E.; Khurana, S.; Schmitt, N.; Kurup, P.; Mueller, C.; Obermoser, G.; Palucka, A.K.; Albrecht, R.A.; Garcia-Sastre, A.; Golding, H.; et al. ICOS(+)PD-1(+)CXCR3(+) T follicular helper cells contribute to the generation of high-avidity antibodies following influenza vaccination. Sci. Rep. 2016, 6, 26494. [Google Scholar]
- Bentebibel, S.E.; Lopez, S.; Obermoser, G.; Schmitt, N.; Mueller, C.; Harrod, C.; Flano, E.; Mejias, A.; Albrecht, R.A.; Blankenship, D.; et al. Induction of ICOS+CXCR3+CXCR5+ TH cells correlates with antibody responses to influenza vaccination. Sci. Transl. Med. 2013, 5, 176ra32. [Google Scholar]
- Byford, E.; Carr, M.; Piñon, L.; Ahearne, M.J.; Wagner, S.D. Isolation of CD4+ T-cells and Analysis of Circulating T-follicular Helper (cTfh) Cell Subsets from Peripheral Blood Using 6-color Flow Cytometry. J. Vis. Exp. 2019, e58431. [Google Scholar] [CrossRef]
- Izmirly, A.M.; Pelletier, A.N.; Connors, J.; Taramangalam, B.; Alturki, S.O.; Gordon, E.A.; Alturki, S.O.; Mell, J.C.; Swaminathan, G.; Karthik, V.; et al. Pre-vaccination frequency of circulatory Tfh is associated with robust immune response to TV003 dengue vaccine. PLoS Pathog. 2022, 18, e1009903. [Google Scholar] [CrossRef]
- Kudryavtsev, I.V.; Arsentieva, N.A.; Batsunov, O.K.; Korobova, Z.R.; Khamitova, I.V.; Isakov, D.V.; Kuznetsova, R.N.; Rubinstein, A.A.; Stanevich, O.V.; Lebedeva, A.A.; et al. Alterations in B Cell and Follicular T-Helper Cell Subsets in Patients with Acute COVID-19 and COVID-19 Convalescents. Curr. Issues Mol. Biol. 2021, 44, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, N.; Bentebibel, S.E.; Ueno, H. Phenotype and functions of memory Tfh cells in human blood. Trends Immunol. 2014, 35, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Byford, E.T.; Carr, M.; Ladikou, E.; Ahearne, M.J.; Wagner, S.D. Circulating Tfh1 (cTfh1) cell numbers and PD1 expression are elevated in low-grade B-cell non-Hodgkin’s lymphoma and cTfh gene expression is perturbed in marginal zone lymphoma. PLoS ONE 2018, 13, e0190468. [Google Scholar] [CrossRef] [PubMed]
- Ueno, H. Tfh cell response in influenza vaccines in humans: What is visible and what is invisible. Curr. Opin. Immunol. 2019, 59, 9–14. [Google Scholar] [CrossRef]
- Ghamar Talepoor, A.; Khosropanah, S.; Doroudchi, M. Functional subsets of circulating follicular helper T cells in patients with atherosclerosis. Physiol. Rep. 2020, 8, e14637. [Google Scholar] [CrossRef]
- Morita, R.; Schmitt, N.; Bentebibel, S.E.; Ranganathan, R.; Bourdery, L.; Zurawski, G.; Foucat, E.; Dullaers, M.; Oh, S.; Sabzghabaei, N.; et al. Human blood CXCR5(+)CD4(+) T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity 2011, 34, 108–121. [Google Scholar] [CrossRef]
- Johnston, R.J.; Poholek, A.C.; DiToro, D.; Yusuf, I.; Eto, D.; Barnett, B.; Dent, A.L.; Craft, J.; Crotty, S. Bcl6 and Blimp-1 are reciprocal and antagonistic regulators of T follicular helper cell differentiation. Science 2009, 325, 1006–1010. [Google Scholar] [CrossRef]
- Kroenke, M.A.; Eto, D.; Locci, M.; Cho, M.; Davidson, T.; Haddad, E.K.; Crotty, S. Bcl6 and Maf cooperate to instruct human follicular helper CD4 T cell differentiation. J. Immunol. 2012, 188, 3734–3744. [Google Scholar] [CrossRef]
- Boswell, K.L.; Paris, R.; Boritz, E.; Ambrozak, D.; Yamamoto, T.; Darko, S.; Wloka, K.; Wheatley, A.; Narpala, S.; McDermott, A.; et al. Loss of circulating CD4 T cells with B cell helper function during chronic HIV infection. PLoS Pathog. 2014, 10, e1003853. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.S.; Suryani, S.; Avery, D.T.; Chan, A.; Nanan, R.; Santner-Nanan, B.; Deenick, E.K.; Tangye, S.G. Early commitment of naïve human CD4(+) T cells to the T follicular helper (T(FH)) cell lineage is induced by IL-12. Immunol. Cell Biol. 2009, 87, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Tang, Y.; Jiang, Q.; Jiang, D.; Zhang, Y.; Lv, Y.; Xu, D.; Wu, J.; Xie, J.; Wen, C.; et al. Follicular Helper T Cells in the Immunopathogenesis of SARS-CoV-2 Infection. Front. Immunol. 2021, 12, 731100. [Google Scholar] [CrossRef] [PubMed]
- Baiyegunhi, O.; Ndlovu, B.; Ogunshola, F.; Ismail, N.; Walker, B.D.; Ndung’u, T.; Ndhlovu, Z.M. Frequencies of Circulating Th1-Biased T Follicular Helper Cells in Acute HIV-1 Infection Correlate with the Development of HIV-Specific Antibody Responses and Lower Set Point Viral Load. J. Virol. 2018, 92, e00659-18. [Google Scholar] [CrossRef] [PubMed]
- Bos, S.; Graber, A.L.; Cardona-Ospina, J.A.; Duarte, E.M.; Zambrana, J.V.; Ruíz Salinas, J.A.; Mercado-Hernandez, R.; Singh, T.; Katzelnick, L.C.; de Silva, A.; et al. The association of neutralizing antibodies with protection against symptomatic dengue virus infection varies by serotype, prior infection history, and assay condition. medRxiv 2023. [Google Scholar] [CrossRef]
- Sandberg, J.T.; Ols, S.; Löfling, M.; Varnaitė, R.; Lindgren, G.; Nilsson, O.; Rombo, L.; Kalén, M.; Loré, K.; Blom, K.; et al. Activation and Kinetics of Circulating T Follicular Helper Cells, Specific Plasmablast Response, and Development of Neutralizing Antibodies following Yellow Fever Virus Vaccination. J. Immunol. 2021, 207, 1033–1043. [Google Scholar] [CrossRef]
- Wijesinghe, A.; Gamage, J.; Goonewardena, H.; Gomes, L.; Jayathilaka, D.; Wijeratne, D.T.; de Alwis, R.; Jeewandara, C.; Wijewickrama, A.; Ogg, G.S.; et al. Phenotype and functionality of follicular helper T cells in patients with acute dengue infection. J. Biomed. Sci. 2020, 27, 50. [Google Scholar] [CrossRef]
- Halstead, B.S. Dengue Virus–Mosquito Interactions. Annu. Rev. Entomol. 2008, 53, 273–291. [Google Scholar] [CrossRef]
- Dempsey, A.L. Zika-neutralizing antibodies. Nat. Immunol. 2017, 18, 603. [Google Scholar] [CrossRef]
- Wilder-Smith, A.; Hombach, J.; Ferguson, N.; Selgelid, M.; O’Brien, K.; Vannice, K.; Barrett, A.; Ferdinand, E.; Flasche, S.; Guzman, M.; et al. Deliberations of the Strategic Advisory Group of Experts on Immunization on the use of CYD-TDV dengue vaccine. Lancet Infect. Dis. 2019, 19, e31–e38. [Google Scholar] [CrossRef]
- Shukla, R.; Ramasamy, V.; Shanmugam, R.K.; Ahuja, R.; Khanna, N. Antibody-Dependent Enhancement: A Challenge for Developing a Safe Dengue Vaccine. Front. Cell. Infect. Microbiol. 2020, 10, 572681. [Google Scholar] [CrossRef]
- Henein, S.; Swanstrom, J.; Byers, A.M.; Moser, J.M.; Shaik, S.F.; Bonaparte, M.; Jackson, N.; Guy, B.; Baric, R.; de Silva, A.M. Dissecting Antibodies Induced by a Chimeric Yellow Fever-Dengue, Live-Attenuated, Tetravalent Dengue Vaccine (CYD-TDV) in Naive and Dengue-Exposed Individuals. J. Infect. Dis. 2017, 215, 351–358. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Pérez, P.N.; Collazo-Llera, J.A.; Rodríguez-Alvarado, F.A.; Rivera-Amill, V. The Emerging Role of Circulating T Follicular Helper Cells in Dengue Virus Immunity: Balancing Protection and Pathogenesis. Viruses 2025, 17, 652. https://doi.org/10.3390/v17050652
Flores-Pérez PN, Collazo-Llera JA, Rodríguez-Alvarado FA, Rivera-Amill V. The Emerging Role of Circulating T Follicular Helper Cells in Dengue Virus Immunity: Balancing Protection and Pathogenesis. Viruses. 2025; 17(5):652. https://doi.org/10.3390/v17050652
Chicago/Turabian StyleFlores-Pérez, Paola N., José A. Collazo-Llera, Fabiola A. Rodríguez-Alvarado, and Vanessa Rivera-Amill. 2025. "The Emerging Role of Circulating T Follicular Helper Cells in Dengue Virus Immunity: Balancing Protection and Pathogenesis" Viruses 17, no. 5: 652. https://doi.org/10.3390/v17050652
APA StyleFlores-Pérez, P. N., Collazo-Llera, J. A., Rodríguez-Alvarado, F. A., & Rivera-Amill, V. (2025). The Emerging Role of Circulating T Follicular Helper Cells in Dengue Virus Immunity: Balancing Protection and Pathogenesis. Viruses, 17(5), 652. https://doi.org/10.3390/v17050652