
Zika Virus: A Review of Biology, Clinical Impacts, and Coinfections


 , and
, and 
Abstract
1. Introduction
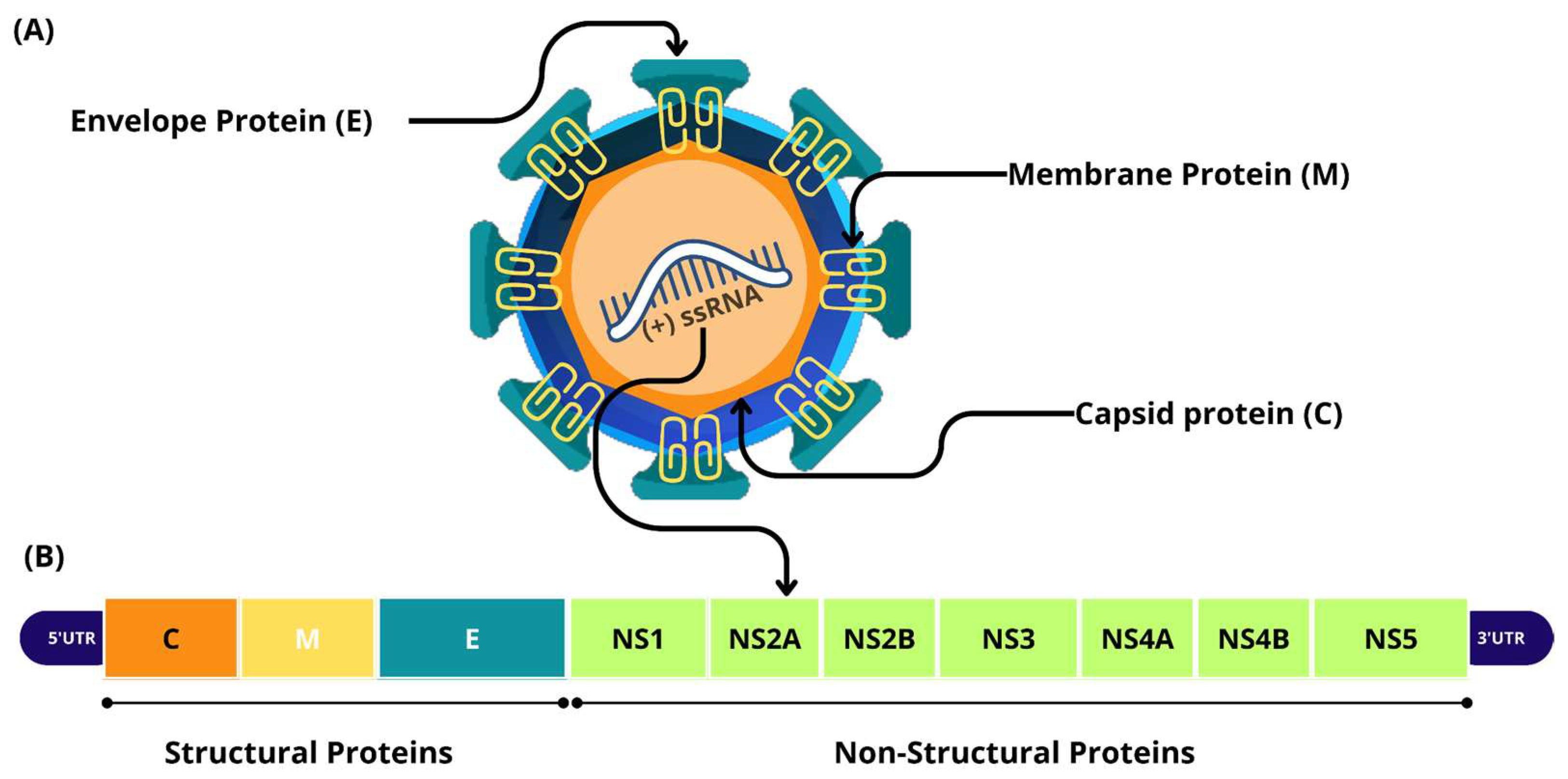
2. Biology of ZIKV
2.1. Infection and Immune Response
2.2. Blood–Brain Barrier and ZIKV Infection
2.3. Congenital Zika Syndrome
2.4. Neurological Manifestations in Adults
3. Cross-Reactive Immunity Among Primary and Secondary Infections
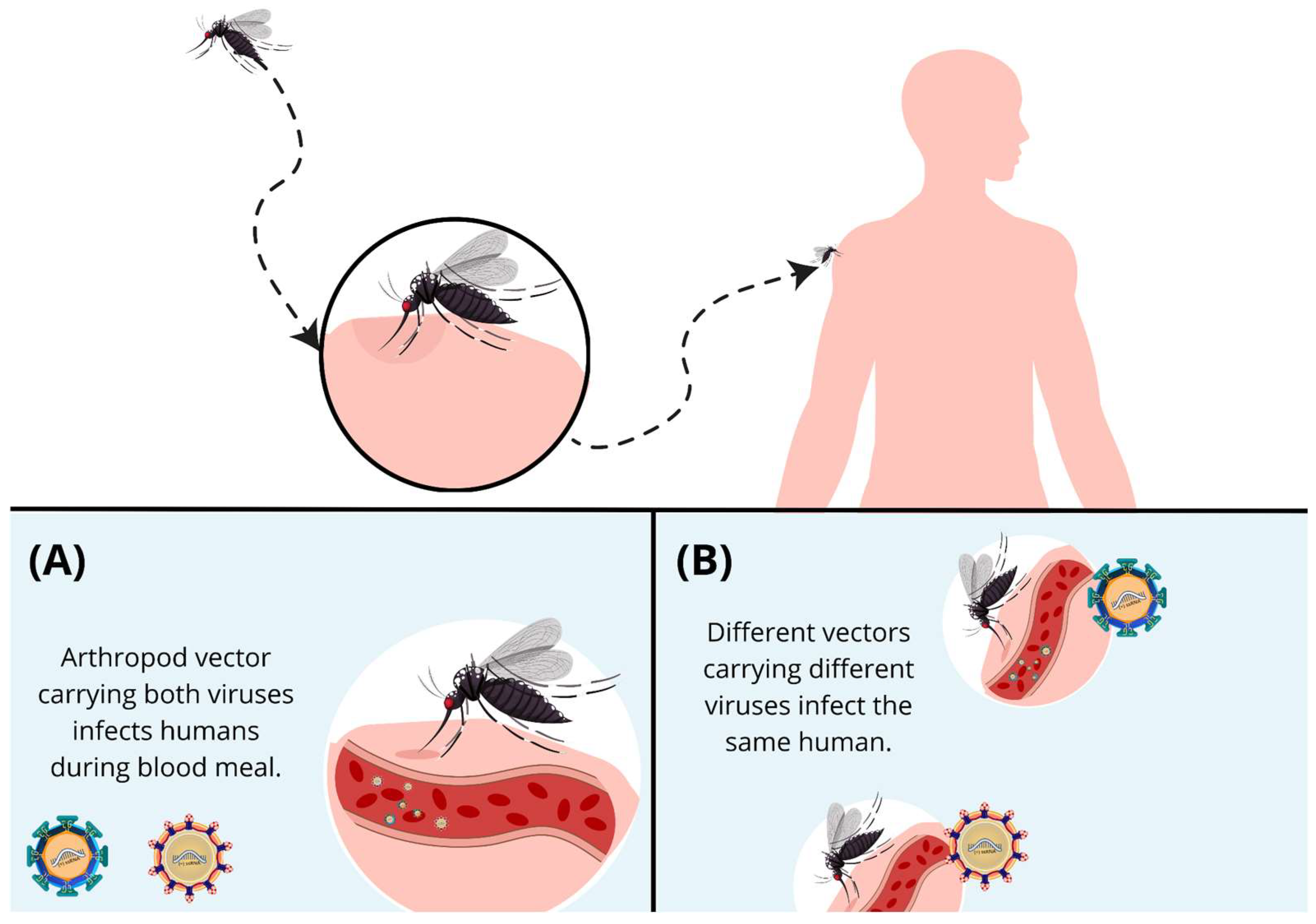
4. Coinfections
4.1. ZIKV and DENV Coinfection: Immune Response and Clinical Outcomes
4.2. ZIKV and CHIKV Coinfection
4.3. ZIKV and HIV-1 Coinfections
4.4. Herpesviridae: HSV-1, EBV, HHV6, and CMV
4.5. Other Viruses: OROV, MAYV, B19V, and SARS-CoV-2
4.6. Other Pathogens: Schistosoma mansoni, Toxoplasma gondii, Leptospira spp., Plasmodium spp., and Candida tropicalis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Souza, A.D.; Abreu, M.C.; Oliveira-Júnior, J.F. Impact of Climate Change on Human Infectious Diseases: Dengue. Braz. Arch. Biol. Technol. 2021, 64, e21190502. [Google Scholar] [CrossRef]
- Tajudeen, Y.A.; Oladunjoye, I.O.; Mustapha, M.O.; Mustapha, S.T.; Ajide-Bamigboye, N.T. Tackling the Global Health Threat of Arboviruses: An Appraisal of the Three Holistic Approaches to Health. Health Promot. Perspect. 2021, 11, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Alves, V.S.; Leite-Aguiar, R.; da Silva, J.P.; Coutinho-Silva, R.; Savio, L.E.B. Purinergic Signaling in Infectious Diseases of the Central Nervous System. Brain. Behav. Immun. 2020, 89, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Pant, P.; Bhagat, R.; Seth, P. Zika Virus E Protein Modulates Functions of Human Brain Microvascular Endothelial Cells and Astrocytes: Implications on Blood-Brain Barrier Properties. Front. Cell. Neurosci. 2023, 17, 1173120. [Google Scholar] [CrossRef]
- Dick, G.W.A.; Kitchen, S.F.; Haddow, A.J. Zika Virus (I). Isolations and Serological Specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- MacNamara, F.N. Zika Virus: A Report on Three Cases of Human Infection during an Epidemic of Jaundice in Nigeria. Trans. R. Soc. Trop. Med. Hyg. 1954, 48, 139–145. [Google Scholar] [CrossRef]
- Abrams, R.P.M.; Solis, J.; Nath, A. Therapeutic Approaches for Zika Virus Infection of the Nervous System. Neurotherapeutics 2017, 14, 1027–1048. [Google Scholar] [CrossRef]
- Counotte, M.J.; Kim, C.R.; Wang, J.; Bernstein, K.; Deal, C.D.; Broutet, N.J.N.; Low, N. Sexual Transmission of Zika Virus and Other Flaviviruses: A Living Systematic Review. PLoS Med. 2018, 15, e1002611. [Google Scholar] [CrossRef]
- Plourde, A.R.; Bloch, E.M. A Literature Review of Zika Virus. Emerg. Infect. Dis. 2016, 22, 1185–1192. [Google Scholar] [CrossRef]
- Raphael, L.M.S.; De Mello, I.S.; Gómez, M.M.; Ribeiro, I.P.; Furtado, N.D.; Lima, N.S.; Dos Santos, A.A.C.; Fernandes, D.R.; Da Cruz, S.O.D.; Damasceno, L.S.; et al. Phenotypic and Genetic Variability of Isolates of ZIKV-2016 in Brazil. Microorganisms 2022, 10, 854. [Google Scholar] [CrossRef]
- Cerbino-Neto, J.; Mesquita, E.C.; Souza, T.M.L.; Parreira, V.; Wittlin, B.B.; Durovni, B.; Lemos, M.C.F.; Vizzoni, A.; Bispo De Filippis, A.M.; Sampaio, S.A.; et al. Clinical Manifestations of Zika Virus Infection, Rio de Janeiro, Brazil, 2015. Emerg. Infect. Dis. 2016, 22, 1318–1320. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Ko, A.I.; Baud, D. Zika Virus Infection—After the Pandemic. N. Engl. J. Med. 2019, 381, 1444–1457. [Google Scholar] [CrossRef] [PubMed]
- Rawal, G.; Yadav, S.; Kumar, R. Zika Virus: An Overview. J. Fam. Med. Prim. Care 2016, 5, 523. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Oo, H.H.; Balne, P.K.; Ng, L.; Tong, L.; Leo, Y.S. Zika Virus and the Eye. Ocul. Immunol. Inflamm. 2018, 26, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.M.; Medronho, R.D.A.; Cunha, A.J.L.A.D. Zika Virus in Brazil and Worldwide: A Narrative Review. Paediatr. Int. Child Health 2021, 41, 28–35. [Google Scholar] [CrossRef]
- Wimalasiri-Yapa, B.M.C.R.; Yapa, H.E.; Huang, X.; Hafner, L.M.; Kenna, T.J.; Frentiu, F.D. Zika Virus and Arthritis/Arthralgia: A Systematic Review and Meta-Analysis. Viruses 2020, 12, 1137. [Google Scholar] [CrossRef]
- Krauer, F.; Riesen, M.; Reveiz, L.; Oladapo, O.T.; Martínez-Vega, R.; Porgo, T.V.; Haefliger, A.; Broutet, N.J.; Low, N.; WHO Zika Causality Working Group. Zika Virus Infection as a Cause of Congenital Brain Abnormalities and Guillain–Barré Syndrome: Systematic Review. PLoS Med. 2017, 14, e1002203. [Google Scholar] [CrossRef]
- Lobkowicz, L.; Ramond, A.; Sanchez Clemente, N.; Ximenes, R.A.D.A.; Miranda-Filho, D.D.B.; Montarroyos, U.R.; Martelli, C.M.T.; De Araújo, T.V.B.; Brickley, E.B. The Frequency and Clinical Presentation of Zika Virus Coinfections: A Systematic Review. BMJ Glob. Health 2020, 5, e002350. [Google Scholar] [CrossRef]
- Beaver, J.T.; Lelutiu, N.; Habib, R.; Skountzou, I. Evolution of Two Major Zika Virus Lineages: Implications for Pathology, Immune Response, and Vaccine Development. Front. Immunol. 2018, 9, 1640. [Google Scholar] [CrossRef]
- Kazmi, S.S.; Ali, W.; Bibi, N.; Nouroz, F. A Review on Zika Virus Outbreak, Epidemiology, Transmission and Infection Dynamics. J. Biol. Res.-Thessalon. 2020, 27, 5. [Google Scholar] [CrossRef]
- Pettersson, J.H.-O.; Eldholm, V.; Seligman, S.J.; Lundkvist, Å.; Falconar, A.K.; Gaunt, M.W.; Musso, D.; Nougairède, A.; Charrel, R.; Gould, E.A.; et al. How Did Zika Virus Emerge in the Pacific Islands and Latin America? mBio 2016, 7, e01239-16. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, M.A.; Balaraman, V.; Schust, D.J.; Ezashi, T.; Roberts, R.M.; Franz, A.W.E. African and Asian Strains of Zika Virus Differ in Their Ability to Infect and Lyse Primitive Human Placental Trophoblast. PLoS ONE 2018, 13, e0200086. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, G.D.S.; Brites, C.; Drexler, J.F.; Moreira-Soto, A.; Miranda, F.; Martins, E. Expansão da circulação do vírus Zika da África à América, 1947-2018: Revisão da literatura*. Epidemiol. E Serviços Saúde 2019, 28, 1–23. [Google Scholar] [CrossRef]
- Ayres, C.F.J.; Guedes, D.R.D.; Paiva, M.H.S.; Morais-Sobral, M.C.; Krokovsky, L.; Machado, L.C.; Melo-Santos, M.A.V.; Crespo, M.; Oliveira, C.M.F.; Ribeiro, R.S.; et al. Zika Virus Detection, Isolation and Genome Sequencing through Culicidae Sampling during the Epidemic in Vitória, Espírito Santo, Brazil. Parasit. Vectors 2019, 12, 220. [Google Scholar] [CrossRef]
- White, M.K.; Wollebo, H.S.; David Beckham, J.; Tyler, K.L.; Khalili, K. Zika Virus: An Emergent Neuropathological Agent. Ann. Neurol. 2016, 80, 479–489. [Google Scholar] [CrossRef]
- Latanova, A.; Starodubova, E.; Karpov, V. Flaviviridae Nonstructural Proteins: The Role in Molecular Mechanisms of Triggering Inflammation. Viruses 2022, 14, 1808. [Google Scholar] [CrossRef]
- Nicholls, C.M.R.; Sevvana, M.; Kuhn, R.J. Structure-Guided Paradigm Shifts in Flavivirus Assembly and Maturation Mechanisms. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2020; Volume 108, pp. 33–83. ISBN 978-0-12-820761-1. [Google Scholar]
- Miner, J.J.; Diamond, M.S. Zika Virus Pathogenesis and Tissue Tropism. Cell Host Microbe 2017, 21, 134–142. [Google Scholar] [CrossRef]
- Valdespino-Vázquez, M.Y.; Sevilla-Reyes, E.E.; Lira, R.; Yocupicio-Monroy, M.; Piten-Isidro, E.; Boukadida, C.; Hernández-Pando, R.; Soriano-Jimenez, J.D.; Herrera-Salazar, A.; Figueroa-Damián, R.; et al. Congenital Zika Syndrome and Extra-Central Nervous System Detection of Zika Virus in a Pre-Term Newborn in Mexico. Clin. Infect. Dis. 2019, 68, 903–912. [Google Scholar] [CrossRef]
- Tan, L.Y.; Komarasamy, T.V.; James, W.; Balasubramaniam, V.R.M.T. Host Molecules Regulating Neural Invasion of Zika Virus and Drug Repurposing Strategy. Front. Microbiol. 2022, 13, 743147. [Google Scholar] [CrossRef]
- Berglund, G.; Lennon, C.D.; Badu, P.; Berglund, J.A.; Pager, C.T. Zika Virus Infection in a Cell Culture Model Reflects the Transcriptomic Signatures in Patients. bioRxiv Prepr. Serv. Biol. 2024, 2024.05.25.595842. [Google Scholar] [CrossRef]
- De Sales-Neto, J.M.; Madruga Carvalho, D.C.; Arruda Magalhães, D.W.; Araujo Medeiros, A.B.; Soares, M.M.; Rodrigues-Mascarenhas, S. Zika Virus: Antiviral Immune Response, Inflammation, and Cardiotonic Steroids as Antiviral Agents. Int. Immunopharmacol. 2024, 127, 111368. [Google Scholar] [CrossRef] [PubMed]
- Serman, T.M.; Gack, M.U. Evasion of Innate and Intrinsic Antiviral Pathways by the Zika Virus. Viruses 2019, 11, 970. [Google Scholar] [CrossRef] [PubMed]
- Plociennikowska, A.; Frankish, J.; Moraes, T.; Del Prete, D.; Kahnt, F.; Acuna, C.; Slezak, M.; Binder, M.; Bartenschlager, R. TLR3 Activation by Zika Virus Stimulates Inflammatory Cytokine Production Which Dampens the Antiviral Response Induced by RIG-I-like Receptors. J. Virol. 2021, 95, e01050-20. [Google Scholar] [CrossRef] [PubMed]
- Komarasamy, T.V.; Adnan, N.A.A.; James, W.; Balasubramaniam, V.R.M.T. Zika Virus Neuropathogenesis: The Different Brain Cells, Host Factors and Mechanisms Involved. Front. Immunol. 2022, 13, 773191. [Google Scholar] [CrossRef]
- Lee, J.K.; Kim, J.-A.; Oh, S.-J.; Lee, E.-W.; Shin, O.S. Zika Virus Induces Tumor Necrosis Factor-Related Apoptosis Inducing Ligand (TRAIL)-Mediated Apoptosis in Human Neural Progenitor Cells. Cells 2020, 9, 2487. [Google Scholar] [CrossRef]
- Beys-da-Silva, W.O.; Rosa, R.L.; Santi, L.; Berger, M.; Park, S.K.; Campos, A.R.; Terraciano, P.; Varela, A.P.M.; Teixeira, T.F.; Roehe, P.M.; et al. Zika Virus Infection of Human Mesenchymal Stem Cells Promotes Differential Expression of Proteins Linked to Several Neurological Diseases. Mol. Neurobiol. 2019, 56, 4708–4717. [Google Scholar] [CrossRef]
- Mufrrih, M.; Chen, B.; Chan, S.-W. Zika Virus Induces an Atypical Tripartite Unfolded Protein Response with Sustained Sensor and Transient Effector Activation and a Blunted BiP Response. mSphere 2021, 6, e00361-21. [Google Scholar] [CrossRef]
- Hoffmann, H.-H.; Schneider, W.M.; Blomen, V.A.; Scull, M.A.; Hovnanian, A.; Brummelkamp, T.R.; Rice, C.M. Diverse Viruses Require the Calcium Transporter SPCA1 for Maturation and Spread. Cell Host Microbe 2017, 22, 460–470.e5. [Google Scholar] [CrossRef]
- Chen, X.; Yan, Y.; Liu, Z.; Yang, S.; Li, W.; Wang, Z.; Wang, M.; Guo, J.; Li, Z.; Zhu, W.; et al. In Vitro and in Vivo Inhibition of the Host TRPC4 Channel Attenuates Zika Virus Infection. EMBO Mol. Med. 2024, 16, 1817–1839. [Google Scholar] [CrossRef]
- Helms, H.C.; Abbott, N.J.; Burek, M.; Cecchelli, R.; Couraud, P.-O.; Deli, M.A.; Förster, C.; Galla, H.J.; Romero, I.A.; Shusta, E.V.; et al. In Vitro Models of the Blood–Brain Barrier: An Overview of Commonly Used Brain Endothelial Cell Culture Models and Guidelines for Their Use. J. Cereb. Blood Flow Metab. 2016, 36, 862–890. [Google Scholar] [CrossRef]
- Ayala-Nunez, N.V.; Gaudin, R. A Viral Journey to the Brain: Current Considerations and Future Developments. PLoS Pathog. 2020, 16, e1008434. [Google Scholar] [CrossRef] [PubMed]
- Vhp, L.; Aragão, M.; Pinho, R.; Hazin, A.; Paciorkowski, A.; Penalva De Oliveira, A.; Masruha, M.R. Congenital Zika Virus Infection: A Review with Emphasis on the Spectrum of Brain Abnormalities. Curr. Neurol. Neurosci. Rep. 2020, 20, 49. [Google Scholar] [CrossRef] [PubMed]
- Ades, A.E.; Soriano-Arandes, A.; Alarcon, A.; Bonfante, F.; Thorne, C.; Peckham, C.S.; Giaquinto, C. Vertical Transmission of Zika Virus and Its Outcomes: A Bayesian Synthesis of Prospective Studies. Lancet Infect. Dis. 2021, 21, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Mendoza, C.K.; Rice, M.E.; Galang, R.R.; Fulton, A.C.; VanMaldeghem, K.; Prado, M.V.; Ellis, E.; Anesi, M.S.; Simeone, R.M.; Petersen, E.E.; et al. Pregnancy Outcomes After Maternal Zika Virus Infection During Pregnancy—U.S. Territories, January 1, 2016–April 25, 2017. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 615–621. [Google Scholar] [CrossRef]
- Coutinho, C.; Negrini, S.; Araujo, D.; Teixeira, S.; Amaral, F.; Moro, M.; Fernandes, J.; Da Motta, M.; Negrini, B.; Caldas, C.; et al. Early Maternal Zika Infection Predicts Severe Neonatal Neurological Damage: Results from the Prospective Natural History of Zika Virus Infection in Gestation Cohort Study. BJOG Int. J. Obstet. Gynaecol. 2021, 128, 317–326. [Google Scholar] [CrossRef]
- Cunha, A.J.L.A.D.; De Magalhães-Barbosa, M.C.; Lima-Setta, F.; Medronho, R.D.A.; Prata-Barbosa, A. Microcephaly Case Fatality Rate Associated with Zika Virus Infection in Brazil: Current Estimates. Pediatr. Infect. Dis. J. 2017, 36, 528–530. [Google Scholar] [CrossRef]
- Antoniou, E.; Orovou, E.; Andronikidi, P.E.; Orovas, C.; Rigas, N.; Palaska, E.; Sarella, A.; Iatrakis, G.; Voyiatzaki, C. Congenital Zika Infection and the Risk of Neurodevelopmental, Neurological, and Urinary Track Disorders in Early Childhood. A Systematic Review. Viruses 2021, 13, 1671. [Google Scholar] [CrossRef]
- Pereira, H.V.F.S.; Dos Santos, S.P.; Amâncio, A.P.R.L.; De Oliveira-Szejnfeld, P.S.; Flor, E.O.; De Sales Tavares, J.; Ferreira, R.V.B.; Tovar-Moll, F.; De Amorim, M.M.R.; Melo, A. Neurological Outcomes of Congenital Zika Syndrome in Toddlers and Preschoolers: A Case Series. Lancet Child Adolesc. Health 2020, 4, 378–387. [Google Scholar] [CrossRef]
- Cicuto Ferreira Rocha, N.A.; De Campos, A.C.; Cicuto Ferreira Rocha, F.; Pereira Dos Santos Silva, F. Microcephaly and Zika Virus: Neuroradiological Aspects, Clinical Findings and a Proposed Framework for Early Evaluation of Child Development. Infant Behav. Dev. 2017, 49, 70–82. [Google Scholar] [CrossRef]
- Coffey, L.L.; Keesler, R.I.; Pesavento, P.A.; Woolard, K.; Singapuri, A.; Watanabe, J.; Cruzen, C.; Christe, K.L.; Usachenko, J.; Yee, J.; et al. Intraamniotic Zika Virus Inoculation of Pregnant Rhesus Macaques Produces Fetal Neurologic Disease. Nat. Commun. 2018, 9, 2414. [Google Scholar] [CrossRef]
- Janssens, S.; Schotsaert, M.; Karnik, R.; Balasubramaniam, V.; Dejosez, M.; Meissner, A.; García-Sastre, A.; Zwaka, T.P. Zika Virus Alters DNA Methylation of Neural Genes in an Organoid Model of the Developing Human Brain. mSystems 2018, 3, e00219-17. [Google Scholar] [CrossRef] [PubMed]
- Halani, S.; Tombindo, P.E.; O’Reilly, R.; Miranda, R.N.; Erdman, L.K.; Whitehead, C.; Bielecki, J.M.; Ramsay, L.; Ximenes, R.; Boyle, J.; et al. Clinical Manifestations and Health Outcomes Associated with Zika Virus Infections in Adults: A Systematic Review. PLoS Negl. Trop. Dis. 2021, 15, e0009516. [Google Scholar] [CrossRef] [PubMed]
- Filgueiras, I.S.; Torrentes De Carvalho, A.; Cunha, D.P.; Mathias Da Fonseca, D.L.; El Khawanky, N.; Freire, P.P.; Cabral-Miranda, G.; Schimke, L.F.; Camara, N.O.S.; Ochs, H.D.; et al. The Clinical Spectrum and Immunopathological Mechanisms Underlying ZIKV-Induced Neurological Manifestations. PLoS Negl. Trop. Dis. 2021, 15, e0009575. [Google Scholar] [CrossRef]
- Muñoz, L.S.; Parra, B.; Pardo, C.A. Neuroviruses Emerging in the Americas Study Neurological Implications of Zika Virus Infection in Adults. J. Infect. Dis. 2017, 216, S897–S905. [Google Scholar] [CrossRef]
- Robbiani, D.F.; Olsen, P.C.; Costa, F.; Wang, Q.; Oliveira, T.Y.; Nery, N.; Aromolaran, A.; Do Rosário, M.S.; Sacramento, G.A.; Cruz, J.S.; et al. Risk of Zika Microcephaly Correlates with Features of Maternal Antibodies. J. Exp. Med. 2019, 216, 2302–2315. [Google Scholar] [CrossRef] [PubMed]
- Charniga, K.; Cucunubá, Z.M.; Walteros, D.M.; Mercado, M.; Prieto, F.; Ospina, M.; Nouvellet, P.; Donnelly, C.A. Descriptive Analysis of Surveillance Data for Zika Virus Disease and Zika Virus-Associated Neurological Complications in Colombia, 2015–2017. PLoS ONE 2021, 16, e0252236. [Google Scholar] [CrossRef]
- Akrami, K.M.; de Nogueira, B.M.F.; do Rosário, M.S.; de Moraes, L.; Cordeiro, M.T.; Haddad, R.; Gomes, L.N.; de Pádua Carvalho, I.; Dos Reis Pimentel, E.; de Jesus Silva, J.; et al. The Re-Emergence of Zika in Brazil in 2020: A Case of Guillain Barré Syndrome during the Low Season for Arboviral Infections. J. Travel Med. 2020, 27, taaa165. [Google Scholar] [CrossRef]
- Berry, N.; Ferguson, D.; Ham, C.; Hall, J.; Jenkins, A.; Giles, E.; Devshi, D.; Kempster, S.; Rose, N.; Dowall, S.; et al. High Susceptibility, Viral Dynamics and Persistence of South American Zika Virus in New World Monkey Species. Sci. Rep. 2019, 9, 14495. [Google Scholar] [CrossRef]
- Pantoja, P.; Pérez-Guzmán, E.X.; Rodríguez, I.V.; White, L.J.; González, O.; Serrano, C.; Giavedoni, L.; Hodara, V.; Cruz, L.; Arana, T.; et al. Zika Virus Pathogenesis in Rhesus Macaques Is Unaffected by Pre-Existing Immunity to Dengue Virus. Nat. Commun. 2017, 8, 15674. [Google Scholar] [CrossRef]
- Oliveira, R.A.; De Oliveira-Filho, E.F.; Fernandes, A.I.; Brito, C.A.; Marques, E.T.; Tenório, M.C.; Gil, L.H. Previous Dengue or Zika Virus Exposure Can Drive to Infection Enhancement or Neutralisation of Other Flaviviruses. Mem. Inst. Oswaldo Cruz 2019, 114, e190098. [Google Scholar] [CrossRef]
- Thomas, S.; Smatti, M.K.; Ouhtit, A.; Cyprian, F.S.; Almaslamani, M.A.; Thani, A.A.; Yassine, H.M. Antibody-Dependent Enhancement (ADE) and the Role of Complement System in Disease Pathogenesis. Mol. Immunol. 2022, 152, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Masel, J.; McCracken, M.K.; Gleeson, T.; Morrison, B.; Rutherford, G.; Imrie, A.; Jarman, R.G.; Koren, M.; Pollett, S. Does Prior Dengue Virus Exposure Worsen Clinical Outcomes of Zika Virus Infection? A Systematic Review, Pooled Analysis and Lessons Learned. PLoS Negl. Trop. Dis. 2019, 13, e0007060. [Google Scholar] [CrossRef] [PubMed]
- Tonnerre, P.; Melgaço, J.G.; Torres-Cornejo, A.; Pinto, M.A.; Yue, C.; Blümel, J.; De Sousa, P.S.F.; De Mello, V.D.M.; Moran, J.; De Filippis, A.M.B.; et al. Evolution of the Innate and Adaptive Immune Response in Women with Acute Zika Virus Infection. Nat. Microbiol. 2019, 5, 76–83. [Google Scholar] [CrossRef]
- Estofolete, C.F.; Terzian, A.C.B.; Colombo, T.E.; De Freitas Guimarães, G.; Ferraz, H.C.; Da Silva, R.A.; Greque, G.V.; Nogueira, M.L. Co-Infection between Zika and Different Dengue Serotypes during DENV Outbreak in Brazil. J. Infect. Public Health 2019, 12, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Zambrana, J.V.; Hasund, C.M.; Aogo, R.A.; Bos, S.; Arguello, S.; Gonzalez, K.; Collado, D.; Miranda, T.; Kuan, G.; Gordon, A.; et al. Primary Exposure to Zika Virus Is Linked with Increased Risk of Symptomatic Dengue Virus Infection with Serotypes 2, 3, and 4, but Not 1. Sci. Transl. Med. 2024, 16, eadn2199. [Google Scholar] [CrossRef]
- Breitbach, M.E.; Newman, C.M.; Dudley, D.M.; Stewart, L.M.; Aliota, M.T.; Koenig, M.R.; Shepherd, P.M.; Yamamoto, K.; Crooks, C.M.; Young, G.; et al. Primary Infection with Dengue or Zika Virus Does Not Affect the Severity of Heterologous Secondary Infection in Macaques. PLOS Pathog. 2019, 15, e1007766. [Google Scholar] [CrossRef]
- Hassert, M.; Scroggins, S.; Coleman, A.K.; Shacham, E.; Brien, J.D.; Pinto, A.K. Heterologous Flavivirus Exposure Provides Varying Degrees of Cross-Protection from Zika Virus in a Mouse Model of Infection. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ahmed, S.; Sultana, S.; Kundu, S.; Alam, S.S.; Hossan, T.; Islam, M.A. Global Prevalence of Zika and Chikungunya Coinfection: A Systematic Review and Meta-Analysis. Diseases 2024, 12, 31. [Google Scholar] [CrossRef]
- Wu, H.; Zhou, H.-Y.; Zheng, H.; Wu, A. Towards Understanding and Identification of Human Viral Co-Infections. Viruses 2024, 16, 673. [Google Scholar] [CrossRef]
- Cherabuddi, K.; Iovine, N.M.; Shah, K.; White, S.K.; Paisie, T.; Salemi, M.; Morris, J.G.; Lednicky, J.A. Zika and Chikungunya Virus Co-Infection in a Traveller Returning from Colombia, 2016: Virus Isolation and Genetic Analysis. JMM Case Rep. 2016, 3, e005072. [Google Scholar] [CrossRef]
- Alves, R.P.D.S.; Amorim, J.H. Editorial: Arboviruses: Co-Circulation, Co-Transmission, and Co-Infection. Front. Microbiol. 2023, 14, 1321166. [Google Scholar] [CrossRef] [PubMed]
- Rothan, H.A.; Bidokhti, M.R.M.; Byrareddy, S.N. Current Concerns and Perspectives on Zika Virus Co-Infection with Arboviruses and HIV. J. Autoimmun. 2018, 89, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Zhang, M.; Wang, G.; Zhang, D.; Zheng, X.; Li, Y. Biased Virus Transmission Following Sequential Coinfection of Aedes Aegypti with Dengue and Zika Viruses. PLoS Negl. Trop. Dis. 2024, 18, e0012053. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.C.-D.; Weng, S.-C.; Tsao, P.-N.; Chu, J.J.H.; Shiao, S.-H. Co-Infection of Dengue and Zika Viruses Mutually Enhances Viral Replication in the Mosquito Aedes Aegypti. Parasit. Vectors 2023, 16, 160. [Google Scholar] [CrossRef]
- Brustolin, M.; Pujhari, S.; Terradas, G.; Werling, K.; Asad, S.; Metz, H.C.; Henderson, C.A.; Kim, D.; Rasgon, J.L. In Vitro and In Vivo Coinfection and Superinfection Dynamics of Mayaro and Zika Viruses in Mosquito and Vertebrate Backgrounds. J. Virol. 2023, 97, e01778-22. [Google Scholar] [CrossRef]
- Frota, C.C.; Correia, F.G.S.; Alves Vasconcelos, L.R.; De Sousa, P.R.C.; Ferreira, M.L.D.S.; Saraiva, S.P.; Mota Ferreira, R.; Romcy, K.A.M.; Pinheiro, R.F.; De Oliveira, R.T.G.; et al. Positivity of Dengue, Chikungunya, and Zika Infections in Women in Northeast Brazil Post-Zika Epidemic. Pathog. Glob. Health 2023, 117, 485–492. [Google Scholar] [CrossRef]
- ELmojtaba, I.M.; Al-Maqrashi, K.; Al-Musalhi, F.; Al-Salti, N. Optimal Control and Cost Effectiveness Analysis of a Zika–Malaria Co-Infection Model. Partial Differ. Equ. Appl. Math. 2024, 11, 100754. [Google Scholar] [CrossRef]
- Fontenille, D.; Powell, J.R. From Anonymous to Public Enemy: How Does a Mosquito Become a Feared Arbovirus Vector? Pathogens 2020, 9, 265. [Google Scholar] [CrossRef]
- Kuno, G.; Mackenzie, J.; Junglen, S.; Hubálek, Z.; Plyusnin, A.; Gubler, D. Vertebrate Reservoirs of Arboviruses: Myth, Synonym of Amplifier, or Reality? Viruses 2017, 9, 185. [Google Scholar] [CrossRef]
- García-Romero, C.; Carrillo Bilbao, G.A.; Navarro, J.-C.; Martin-Solano, S.; Saegerman, C. Arboviruses in Mammals in the Neotropics: A Systematic Review to Strengthen Epidemiological Monitoring Strategies and Conservation Medicine. Viruses 2023, 15, 417. [Google Scholar] [CrossRef]
- Liang, G.; Gao, X.; Gould, E.A. Factors Responsible for the Emergence of Arboviruses; Strategies, Challenges and Limitations for Their Control. Emerg. Microbes Infect. 2015, 4, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Khongwichit, S.; Chuchaona, W.; Vongpunsawad, S.; Poovorawan, Y. Molecular Surveillance of Arboviruses Circulation and Co-Infection during a Large Chikungunya Virus Outbreak in Thailand, October 2018 to February 2020. Sci. Rep. 2022, 12, 22323. [Google Scholar] [CrossRef] [PubMed]
- Sardi, S.I.; Somasekar, S.; Naccache, S.N.; Bandeira, A.C.; Tauro, L.B.; Campos, G.S.; Chiu, C.Y. Coinfections of Zika and Chikungunya Viruses in Bahia, Brazil, Identified by Metagenomic Next-Generation Sequencing. J. Clin. Microbiol. 2016, 54, 2348–2353. [Google Scholar] [CrossRef]
- Calvet, G.A.; Filippis, A.M.B.; Mendonça, M.C.L.; Sequeira, P.C.; Siqueira, A.M.; Veloso, V.G.; Nogueira, R.M.; Brasil, P. First Detection of Autochthonous Zika Virus Transmission in a HIV-Infected Patient in Rio de Janeiro, Brazil. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2016, 74, 1–3. [Google Scholar] [CrossRef]
- de Araújo, P.S.R.; de Mélo Silva, M.L., Jr.; Tenório, M.; Santos, F.G.T.D. Co-Infection ZIKV and HSV-1 Associated with Meningoencephalitis: Case Report and Literature Review. J. Infect. Public Health 2019, 12, 97–100. [Google Scholar] [CrossRef]
- Martins-Luna, J.; Del Valle-Mendoza, J.; Silva-Caso, W.; Sandoval, I.; Del Valle, L.J.; Palomares-Reyes, C.; Carrillo-Ng, H.; Peña-Tuesta, I.; Aguilar-Luis, M.A. Oropouche Infection a Neglected Arbovirus in Patients with Acute Febrile Illness from the Peruvian Coast. BMC Res. Notes 2020, 13, 67. [Google Scholar] [CrossRef]
- De Souza Costa, M.C.; Siqueira Maia, L.M.; Costa De Souza, V.; Gonzaga, A.M.; Correa De Azevedo, V.; Ramos Martins, L.; Chavez Pavoni, J.H.; Gomes Naveca, F.; Dezengrini Slhessarenko, R. Arbovirus Investigation in Patients from Mato Grosso during Zika and Chikungunya Virus Introdution in Brazil, 2015–2016. Acta Trop. 2019, 190, 395–402. [Google Scholar] [CrossRef]
- Morais, V.D.S.; Reis Santana, L.M.; Bezerra, J.F.; Cruz, F.E.; Rocha De Souza, T.; Tahmasebi, R.; Alves Raposo, R.A.; Marcatti, R.; Garcia Barbosa, E.M.; Hefford, P.M.; et al. Detection of Coinfection with Primate Erythroparvovirus 1 and Arboviruses (DENV, CHIKV and ZIKV) in Individuals with Acute Febrile Illness in the State of Rio Grande Do Norte in 2016. PLoS Negl. Trop. Dis. 2023, 17, e0011701. [Google Scholar] [CrossRef]
- Gunturiz, M.L.; Cortés, L.; Cuevas, E.L.; Chaparro, P.; Ospina, M.L. Toxoplasmosis cerebral congénita e infección por el virus del Zika y del chikunguña: Reporte de un caso. Biomédica 2018, 38, 144–152. [Google Scholar] [CrossRef]
- Alves, L.S.; Estanislau, C.; Barreto, L.; Batista, F.; Toppa, N. Concomitant Testicular Infection by Zika Virus and Schistosoma Mansoni in a Brazilian Young Boy. Rev. Assoc. Médica Bras. 2017, 63, 500–503. [Google Scholar] [CrossRef]
- Otu, A.A.; Udoh, U.A.; Ita, O.I.; Hicks, J.P.; Ukpeh, I.; Walley, J. Prevalence of Zika and Malaria in Patients with Fever in Secondary Healthcare Facilities in South-Eastern Nigeria. Trop. Doct. 2020, 50, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Neaterour, P.; Rivera, A.; Galloway, R.L.; Negrón, M.G.; Rivera-Garcia, B.; Sharp, T.M. Fatal Leptospira Spp./Zika Virus Coinfection-Puerto Rico, 2016. Am. J. Trop. Med. Hyg. 2017, 97, 1085–1087. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.J.; Othman, M.S.; Hiu, J.; Wong, K.T.; Lai, S.K. Idiopathic Ileal Volvulus with Multiple Concomitant Infections in a Starving Man. Malays. J. Pathol. 2021, 43, 81–85. [Google Scholar]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.D.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of Simultaneous Exposure to Arboviruses on Infection and Transmission by Aedes Aegypti Mosquitoes. Nat. Commun. 2017, 8, 15412. [Google Scholar] [CrossRef]
- Villamil-Gómez, W.E.; Rodríguez-Morales, A.J.; Uribe-García, A.M.; González-Arismendy, E.; Castellanos, J.E.; Calvo, E.P.; Álvarez-Mon, M.; Musso, D. Zika, Dengue, and Chikungunya Co-Infection in a Pregnant Woman from Colombia. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2016, 51, 135–138. [Google Scholar] [CrossRef]
- Iovine, N.M.; Lednicky, J.; Cherabuddi, K.; Crooke, H.; White, S.K.; Loeb, J.C.; Cella, E.; Ciccozzi, M.; Salemi, M.; Morris, J.G. Coinfection With Zika and Dengue-2 Viruses in a Traveler Returning from Haiti, 2016: Clinical Presentation and Genetic Analysis. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 64, 72–75. [Google Scholar] [CrossRef]
- Siqueira, C.; Féres, V.; Coutinho, L.; Junqueira, I.; Bento, L.; Montes, L.; Siqueira, J.B. Six Cases of Zika/Dengue Coinfection in a Brazilian Cohort, 2015–2019. Viruses 2020, 12, 1201. [Google Scholar] [CrossRef]
- Ngwe Tun, M.M.; Raini, S.K.; Fernando, L.; Gunawardene, Y.; Inoue, S.; Takamatsu, Y.; Urano, T.; Muthugala, R.; Hapugoda, M.; Morita, K. Epidemiological Evidence of Acute Transmission of Zika Virus Infection in Dengue Suspected Patients in Sri-Lanka. J. Infect. Public Health 2023, 16, 1435–1442. [Google Scholar] [CrossRef]
- Mapalagamage, M.; Weiskopf, D.; Sette, A.; De Silva, A.D. Current Understanding of the Role of T Cells in Chikungunya, Dengue and Zika Infections. Viruses 2022, 14, 242. [Google Scholar] [CrossRef]
- Valiant, W.G.; Mattapallil, M.J.; Higgs, S.; Huang, Y.-J.S.; Vanlandingham, D.L.; Lewis, M.G.; Mattapallil, J.J. Simultaneous Coinfection of Macaques with Zika and Dengue Viruses Does Not Enhance Acute Plasma Viremia but Leads to Activation of Monocyte Subsets and Biphasic Release of Pro-Inflammatory Cytokines. Sci. Rep. 2019, 9, 7877. [Google Scholar] [CrossRef]
- Wichit, S.; Gumpangseth, N.; Hamel, R.; Yainoy, S.; Arikit, S.; Punsawad, C.; Missé, D. Chikungunya and Zika Viruses: Co-Circulation and the Interplay between Viral Proteins and Host Factors. Pathogens 2021, 10, 448. [Google Scholar] [CrossRef] [PubMed]
- Silva, K.R.; Bica, B.E.R.G.; Pimenta, E.S.; Serafim, R.B.; Abreu, M.M.; Gonçalves, J.L.S.; Santana, L.d.S.; Cabral-Castro, M.J.; Peralta, J.M.; Cavalcanti, M.G. Fatal Human Case of Zika and Chikungunya Virus Co-Infection with Prolonged Viremia and Viruria. Diseases 2018, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Prata-Barbosa, A.; Cleto-Yamane, T.L.; Robaina, J.R.; Guastavino, A.B.; De Magalhães-Barbosa, M.C.; Brindeiro, R.D.M.; Medronho, R.A.; Da Cunha, A.J.L.A. Co-Infection with Zika and Chikungunya Viruses Associated with Fetal Death—A Case Report. Int. J. Infect. Dis. 2018, 72, 25–27. [Google Scholar] [CrossRef]
- Brito, C.A.A.; Azevedo, F.; Cordeiro, M.T.; Marques, E.T.A.; Franca, R.F.O. Central and Peripheral Nervous System Involvement Caused by Zika and Chikungunya Coinfection. PLoS Negl. Trop. Dis. 2017, 11, e0005583. [Google Scholar] [CrossRef]
- Villamil-Gómez, W.E.; Sánchez-Herrera, Á.R.; Hernández-Prado, H.; Hernández-Iriarte, J.; Díaz-Ricardo, K.; Vergara-Serpa, O.; Castellanos, J.; Portela-Gaviria, J.E.; Patiño-Valencia, S.; Vargas-Bedoya, D.C.; et al. Zika Virus and HIV Co-Infection in Five Patients from Two Areas of Colombia. J. Formos. Med. Assoc. 2018, 117, 856–858. [Google Scholar] [CrossRef]
- Bidokhti, M.R.M.; Dutta, D.; Madduri, L.S.V.; Woollard, S.M.; Norgren, R.; Giavedoni, L.; Byrareddy, S.N. SIV/SHIV-Zika Co-Infection Does Not Alter Disease Pathogenesis in Adult Non-Pregnant Rhesus Macaque Model. PLoS Negl. Trop. Dis. 2018, 12, e0006811. [Google Scholar] [CrossRef]
- Torres, L.R.; Capobianco, L.R.P.L.; De Souza, A.A.A.; De Almeida Ribeiro, C.R.; Cascabulho, C.; Garzoni, L.R.; Portari, E.A.; Gardel, M.A.; Meuser-Batista, M.; De Paula, V.S.; et al. ZIKV Replication Is Differential in Explants and Cells of Human Placental Which Is Suppressed by HSV-2 Coinfection. Virology 2022, 570, 45–56. [Google Scholar] [CrossRef]
- Rosenstierne, M.W.; Schaltz-Buchholzer, F.; Bruzadelli, F.; Có, A.; Cardoso, P.; Jørgensen, C.S.; Michiels, J.; Heyndrickx, L.; Ariën, K.K.; Fischer, T.K.; et al. Zika Virus IgG in Infants with Microcephaly, Guinea-Bissau, 2016. Emerg. Infect. Dis. 2018, 24, 948–950. [Google Scholar] [CrossRef]
- Lichs, G.G.C.; Fernandez, Z.D.C.; Nascimento, V.A.D.; Alcantara, D.M.C.; Lemos, E.F.; Carvalho, C.M.E.; Demarchi, L.H.F.; Gonçalves, C.C.M.; Naveca, F.G.; Favacho, A.R.D.M. Surveillance of Erythrovirus B19 (B19V) in Patients with Acute Febrile Illness Suspected of Arboviruses in Mato Grosso Do Sul State, Brazil. Front. Microbiol. 2024, 15, 1417434. [Google Scholar] [CrossRef]
- Fantinato, F.F.S.T.; Araújo, E.L.L.; Ribeiro, I.G.; Andrade, M.R.D.; Dantas, A.L.D.M.; Rios, J.M.T.; Silva, O.M.V.D.; Silva, M.D.S.D.; Nóbrega, R.V.; Batista, D.D.A.; et al. Descrição Dos Primeiros Casos de Febre Pelo Vírus Zika Investigados Em Municípios Da Região Nordeste Do Brasil, 2015. Epidemiol. E Serviços Saúde 2016, 25, 683–690. [Google Scholar] [CrossRef]
- Grayo, S. Is the ZIKV Congenital Syndrome and Microcephaly Due to Syndemism with Latent Virus Coinfection? Viruses 2021, 13, 669. [Google Scholar] [CrossRef] [PubMed]
- Vouga, M.; Baud, D.; Jolivet, E.; Najioullah, F.; Monthieux, A.; Schaub, B. Congenital Zika Virus Syndrome… what Else? Two Case Reports of Severe Combined Fetal Pathologies. BMC Pregnancy Childbirth 2018, 18, 356. [Google Scholar] [CrossRef] [PubMed]
- Nunez-Avellaneda, D.; Villagómez, F.R.; Villegas-Pineda, J.C.; Barrios-Palacios, J.; Salazar, M.I.; Machain-Williams, C.; Blitvich, B.J. Evidence of Coinfections between SARS-CoV-2 and Select Arboviruses in Guerrero, Mexico, 2020–2021. Am. J. Trop. Med. Hyg. 2022, 106, 896–899. [Google Scholar] [CrossRef]
- Silva, S.J.R.D.; Magalhães, J.J.F.D.; Pena, L. Simultaneous Circulation of DENV, CHIKV, ZIKV and SARS-CoV-2 in Brazil: An Inconvenient Truth. One Health 2021, 12, 100205. [Google Scholar] [CrossRef]
- Biron, A.; Cazorla, C.; Amar, J.; Pfannstiel, A.; Dupont-Rouzeyrol, M.; Goarant, C. Zika Virus Infection as an Unexpected Finding in a Leptospirosis Patient. JMM Case Rep. 2016, 3, e005033. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Sample/ZIKV Detection Method | Co-Infecting Pathogen (CP) | Sample/Detection Method of the CP | References |
|---|---|---|---|
| Serum/RT-qPCR | DENV-1 and DENV-2 | Serum/RT qPCR | [65] |
| Serum/RT-qPCR | DENV-3 and DENV-4 | Serum/RT qPCR | [83] |
| Serum/mNGS and RT qPCR | CHIKV | Serum/mNGS and RT PCR | [84] |
| Serum/RT-PCR and sequencing | HIV | NI * | [85] |
| Serum/RT-PCR | HSV-1 | Serum/RT-qPCR | [86] |
| Brain, thymus, lungs, kidneys, adrenal glands, and spleen/RT-qPCR | EBV | Brain and Liver/qPCR | [29] |
| HHV-6 | Thymus, kidneys, adrenal glands, and liver/qPCR | ||
| Serum/RT-qPCR | OROV | Serum/RT-PCR | [87] |
| Serum/RT-PCR | MAYV | Serum/RT-PCR and Nested PCR | [88] |
| Serum/RT-qPCR | B19V | Serum/RT-qPCR | [89] |
| Amniotic fluid/RT-qPCR | Toxoplasma gondii | Amniotic fluid/PCR | [90] |
| Blood/RT-PCR | Schistosoma mansoni | Testicle/biopsy | [91] |
| Serum/qualitative lateral flow immuno-chromographic cassettes for IgM and IgG | Plasmodium spp. | Blood/Microscopy of EDTA blood sample pieces | [92] |
| Serum/RT-PCR | Leptospira spp. | Serum/PCR | [93] |
| Blood, liver, lung, and tissue/RT PCR | Candida tropicalis | Blood, pus, peritoneal and pericardial fluid/NI * | [94] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana, L.M.B.; de Moura, I.A.; Mouzinho Ramos Tanaka, Y.; França, R.F.d.O. Zika Virus: A Review of Biology, Clinical Impacts, and Coinfections. Viruses 2025, 17, 637. https://doi.org/10.3390/v17050637
Santana LMB, de Moura IA, Mouzinho Ramos Tanaka Y, França RFdO. Zika Virus: A Review of Biology, Clinical Impacts, and Coinfections. Viruses. 2025; 17(5):637. https://doi.org/10.3390/v17050637
Chicago/Turabian StyleSantana, Lucas Matheus Barreto, Ingrid Andrêssa de Moura, Yuri Mouzinho Ramos Tanaka, and Rafael Freitas de Oliveira França. 2025. "Zika Virus: A Review of Biology, Clinical Impacts, and Coinfections" Viruses 17, no. 5: 637. https://doi.org/10.3390/v17050637
APA StyleSantana, L. M. B., de Moura, I. A., Mouzinho Ramos Tanaka, Y., & França, R. F. d. O. (2025). Zika Virus: A Review of Biology, Clinical Impacts, and Coinfections. Viruses, 17(5), 637. https://doi.org/10.3390/v17050637