
Characterization of Nanobody Binding to Distinct Regions of the SARS-CoV-2 Spike Protein by Flow Virometry



Abstract

1. Introduction
2. Materials and Methods
2.1. FITC Conjugation to Nanobodies
2.2. Cell Culture
2.3. Flow Cytometry
2.4. Plasmids and Pseudotyped Lentivirus Production
2.5. Western Blot Analysis
2.6. Flow Virometry
3. Results
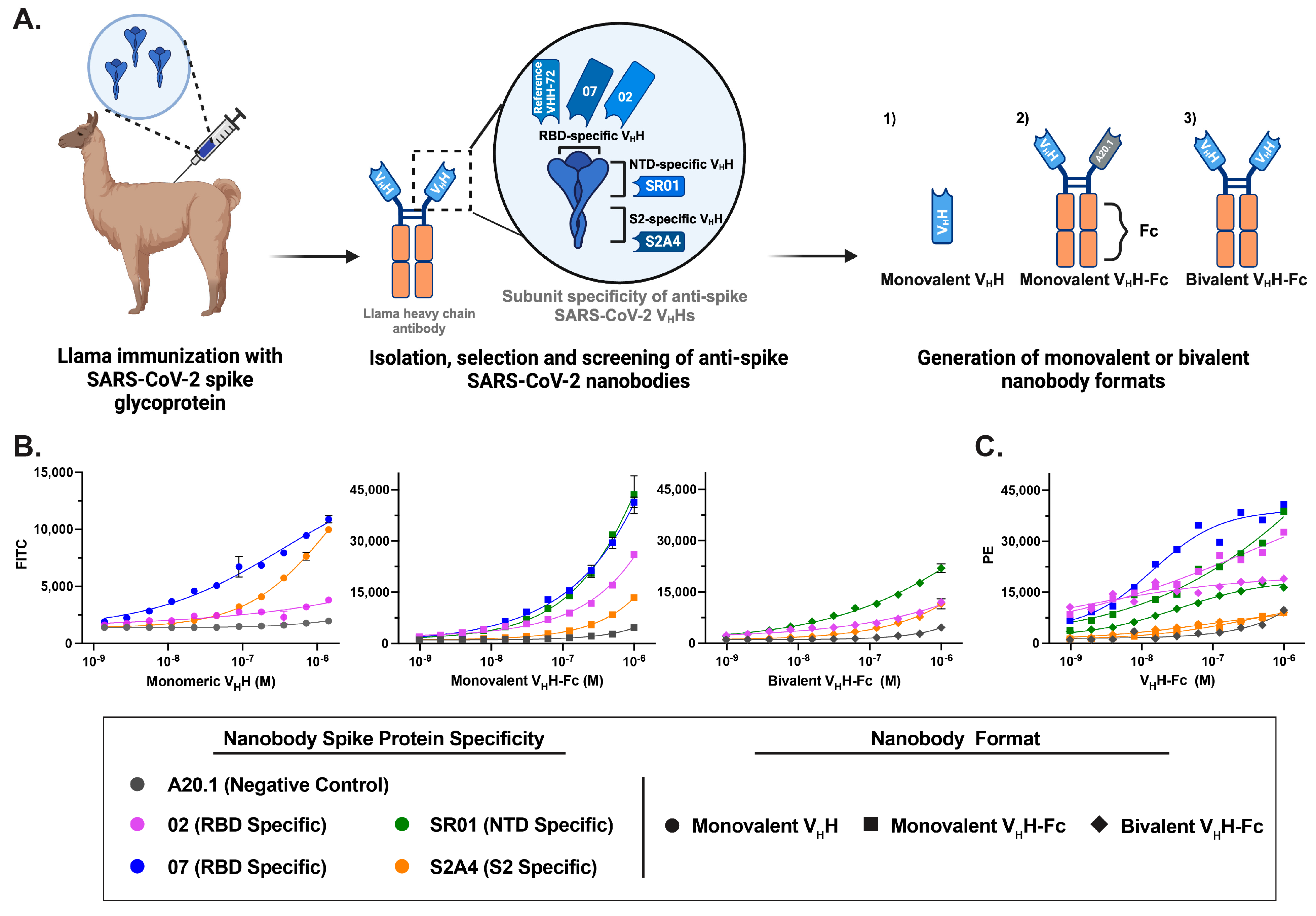
3.1. Generation of Fluorescent Anti-SARS-CoV-2 Spike Glycoprotein Nanobodies
3.2. Characterization of Fluorescent Anti-SARS-CoV-2 Spike Glycoprotein Nanobodies for Cell Binding Assays
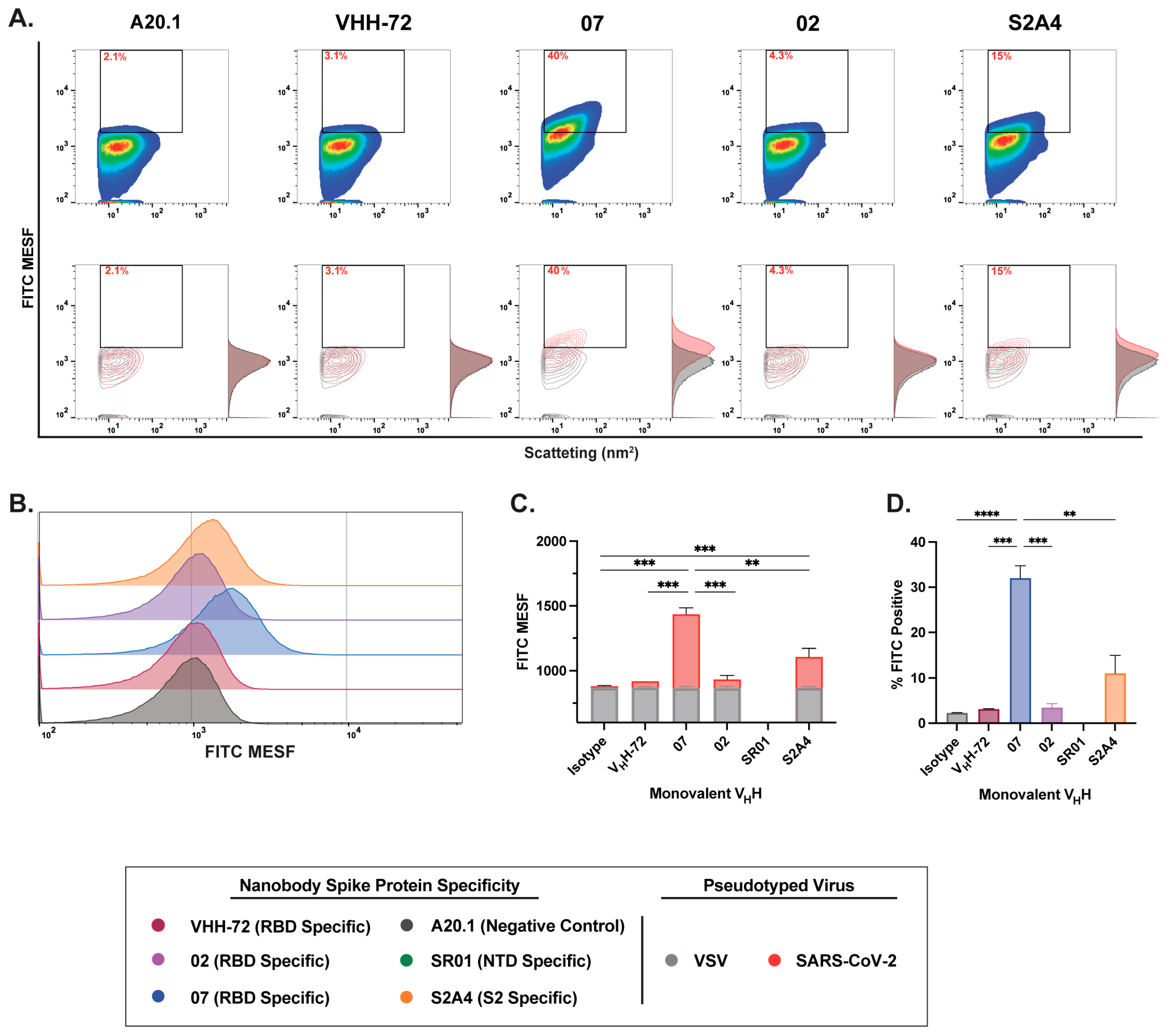
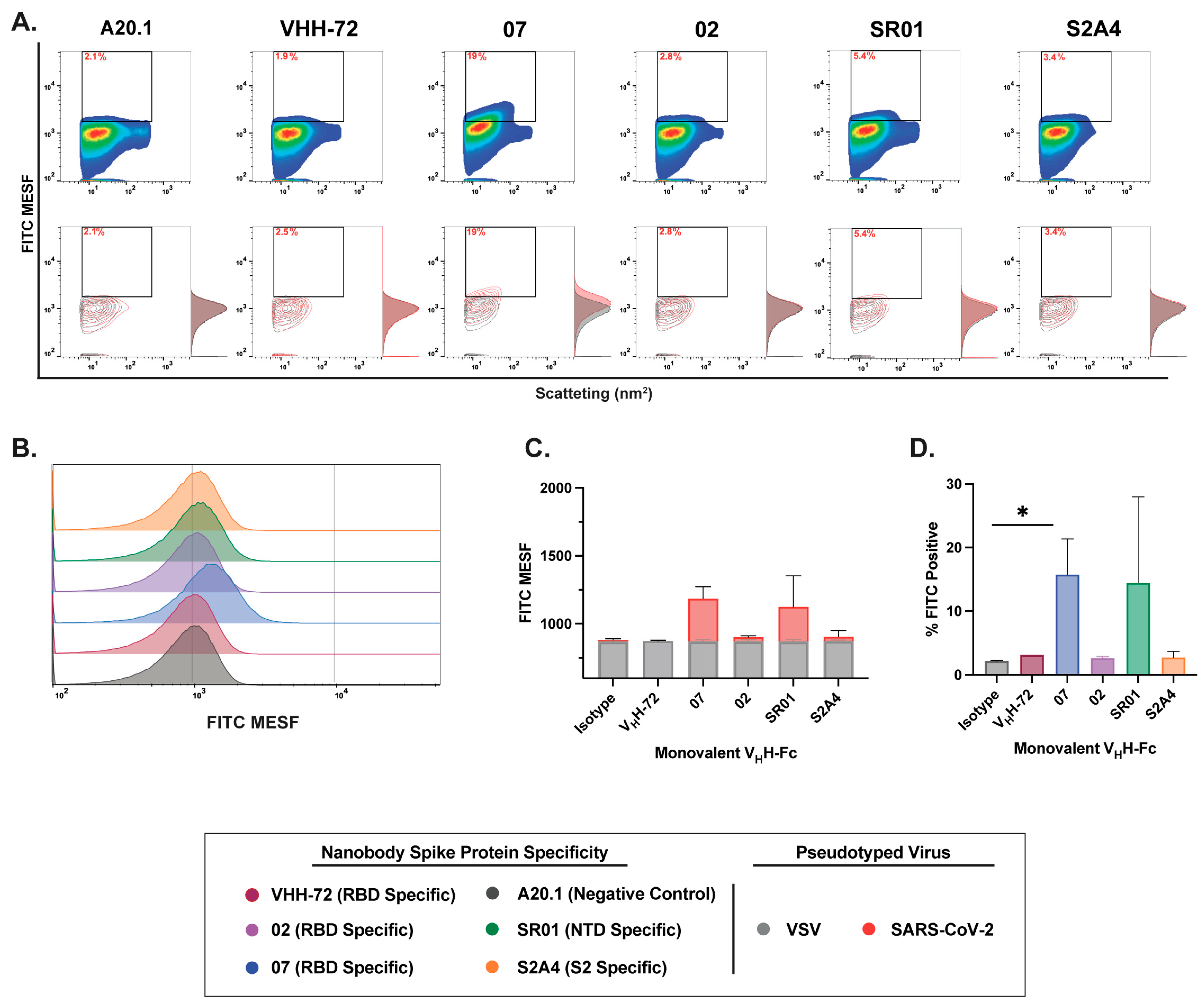
3.3. Characterization of Fluorescent Anti-SARS-CoV-2 Spike Glycoprotein Nanobodies for Flow Virometry Applications
3.4. Nanobody Labeling Enables Enhanced Detection and Resolution of SARS-CoV-2 in Flow Virometry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lippe, R. Flow Virometry: A Powerful Tool To Functionally Characterize Viruses. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Maltseva, M.; Langlois, M.-A. Influence of GlycoGag on the Incorporation of Host Membrane Proteins into the Envelope of the Moloney Murine Leukemia Virus. Front. Virol. 2021, 1, 747253. [Google Scholar] [CrossRef]
- Maltseva, M.; Langlois, M.A. Flow Virometry for Characterizing the Size, Concentration, and Surface Antigens of Viruses. Curr. Protoc. 2022, 2, e368. [Google Scholar] [CrossRef]
- Renner, T.M.; Tang, V.A.; Burger, D.; Langlois, M.A. Intact Viral Particle Counts Measured by Flow Virometry Provide Insight into the Infectivity and Genome Packaging Efficiency of Moloney Murine Leukemia Virus. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Burnie, J.; Guzzo, C. The Incorporation of Host Proteins into the External HIV-1 Envelope. Viruses 2019, 11, 85. [Google Scholar] [CrossRef] [PubMed]
- Zicari, S.; Arakelyan, A.; Fitzgerald, W.; Zaitseva, E.; Chernomordik, L.V.; Margolis, L.; Grivel, J.C. Evaluation of the Maturation of Individual Dengue Virions with Flow Virometry. Virology 2016, 488, 20–27. [Google Scholar] [CrossRef]
- Gaudin, R.; Barteneva, N.S. Sorting of Small Infectious Virus Particles by Flow Virometry Reveals Distinct Infectivity Profiles. Nat. Commun. 2015, 6, 6022. [Google Scholar] [CrossRef]
- Landowski, M.; Dabundo, J.; Liu, Q.; Nicola, A.V.; Aguilar, H.C. Nipah Virion Entry Kinetics, Composition, and Conformational Changes Determined by Enzymatic Virus-Like Particles and New Flow Virometry Tools. J. Virol. 2014, 88, 14197–14206. [Google Scholar] [CrossRef]
- Arakelyan, A.; Fitzgerald, W.; Margolis, L.; Grivel, J.C. Nanoparticle-Based Flow Virometry for the Analysis of Individual Virions. J. Clin. Investig. 2013, 123, 3716–3727. [Google Scholar] [CrossRef]
- Chaudhuri, R.; Lindwasser, O.W.; Smith, W.J.; Hurley, J.H.; Bonifacino, J.S. Downregulation of CD4 by Human Immunodeficiency Virus Type 1 Nef is Dependent on Clathrin and Involves Direct Interaction of Nef with the AP2 Clathrin Adaptor. J. Virol. 2007, 81, 3877–3890. [Google Scholar] [CrossRef]
- Khadivjam, B.; El Bilali, N.; Lippe, R. Analysis and Sorting of Individual HSV-1 Particles by Flow Virometry. Methods Mol. Biol. 2020, 2060, 289–303. [Google Scholar] [CrossRef] [PubMed]
- El Bilali, N.; Duron, J.; Gingras, D.; Lippe, R. Quantitative Evaluation of Protein Heterogeneity within Herpes Simplex Virus 1 Particles. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Loret, S.; El Bilali, N.; Lippe, R. Analysis of Herpes Simplex Virus Type I Nuclear Particles by Flow Cytometry. Cytometry A 2012, 81, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Chaphekar, D.; Fernandes, C.; Persaud, A.T.; Guzzo, C. Comparing Methods to Detect Cellular Proteins on the Surface of HIV-1 Virions. J. Virol. Methods 2025, 333, 115096. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Persaud, A.T.; Chaphekhar, D.; Burnie, J.; Belanger, C.; Tang, V.A.; Guzzo, C. Flow Virometry: Recent Advancements, Best Practices, and Future Frontiers. J. Virol. 2025, e0171724. [Google Scholar] [CrossRef]
- Burnie, J.; Fernandes, C.; Patel, A.; Persaud, A.T.; Chaphekar, D.; Wei, D.; Lee, T.K.H.; Tang, V.A.; Cicala, C.; Arthos, J.; et al. Applying Flow Virometry to Study the HIV Envelope Glycoprotein and Differences Across HIV Model Systems. Viruses 2024, 16, 935. [Google Scholar] [CrossRef]
- Persaud, A.T.; Khela, J.; Fernandes, C.; Chaphekar, D.; Burnie, J.; Tang, V.A.; Colpitts, C.C.; Guzzo, C. Virion-Incorporated CD14 Enables HIV-1 to Bind LPS and Initiate TLR4 Signaling in Immune Cells. J. Virol. 2024, 98, e0036324. [Google Scholar] [CrossRef]
- Burnie, J.; Fernandes, C.; Chaphekar, D.; Wei, D.; Ahmed, S.; Persaud, A.T.; Khader, N.; Cicala, C.; Arthos, J.; Tang, V.A.; et al. Identification of CD38, CD97, and CD278 on the HIV Surface using a Novel Flow Virometry Screening assay. Sci. Rep. 2023, 13, 23025. [Google Scholar] [CrossRef]
- Bonar, M.M.; Tilton, J.C. High Sensitivity Detection and Sorting of Infectious Human Immunodeficiency Virus (HIV-1) Particles by Flow Virometry. Virology 2017, 505, 80–90. [Google Scholar] [CrossRef]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahman, N.; Hamers, R. Naturally Occurring Antibodies Devoid of Light Chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef]
- Barakat, S.; Berksoz, M.; Zahedimaram, P.; Piepoli, S.; Erman, B. Nanobodies as Molecular Imaging Probes. Free Radic. Biol. Med. 2022, 182, 260–275. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S. Applications of Nanobodies. Annu. Rev. Anim. Biosci. 2021, 9, 401–421. [Google Scholar] [CrossRef]
- De Meyer, T.; Muyldermans, S.; Depicker, A. Nanobody-Based Products as Research and Diagnostic tools. Trends Biotechnol. 2014, 32, 263–270. [Google Scholar] [CrossRef]
- Rossotti, M.A.; van Faassen, H.; Tran, A.T.; Sheff, J.; Sandhu, J.K.; Duque, D.; Hewitt, M.; Wen, X.; Bavananthasivam, J.; Beitari, S.; et al. Arsenal of Nanobodies Shows Broad-Spectrum Neutralization Against SARS-CoV-2 Variants of Concern in vitro and in vivo in Hamster Models. Commun. Biol. 2022, 5, 933. [Google Scholar] [CrossRef] [PubMed]
- Koenig, P.A.; Das, H.; Liu, H.; Kummerer, B.M.; Gohr, F.N.; Jenster, L.M.; Schiffelers, L.D.J.; Tesfamariam, Y.M.; Uchima, M.; Wuerth, J.D.; et al. Structure-Guided Multivalent Nanobodies Block SARS-CoV-2 Infection and Suppress Mutational Escape. Science 2021, 371, eabe6230. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Nambulli, S.; Xiao, Z.; Liu, H.; Sang, Z.; Duprex, W.P.; Schneidman-Duhovny, D.; Zhang, C.; Shi, Y. Versatile and Multivalent Nanobodies Efficiently Neutralize SARS-CoV-2. Science 2020, 370, 1479–1484. [Google Scholar] [CrossRef]
- Wrapp, D.; De Vlieger, D.; Corbett, K.S.; Torres, G.M.; Wang, N.; Van Breedam, W.; Roose, K.; van Schie, L.; Team, V.-C.C.-R.; Hoffmann, M.; et al. Structural Basis for Potent Neutralization of Betacoronaviruses by Single-Domain Camelid Antibodies. Cell 2020, 181, 1436–1441. [Google Scholar] [CrossRef]
- Schepens, B.; van Schie, L.; Nerinckx, W.; Roose, K.; Van Breedam, W.; Fijalkowska, D.; Devos, S.; Weyts, W.; De Cae, S.; Vanmarcke, S.; et al. An affinity-enhanced, Broadly Neutralizing Heavy Chain-Only Antibody Protects Against SARS-CoV-2 Infection in Animal Models. Sci. Transl. Med. 2021, 13, eabi7826. [Google Scholar] [CrossRef]
- NIH Clinical Trials, N.C. Dose-Finding, Safety, and Efficacy Study of XVR011 Added to Standard of Care in Patients Hospitalised for COVID-19. 15 April 2022. Available online: https://clinicaltrials.gov/study/NCT04884295#study-plan (accessed on 21 January 2025).
- ExeVIr. ExeVir Pipeline for Antiviral Nanonbodies. 2025. Available online: https://exevir.com/pipeline/ (accessed on 11 January 2025).
- Rujas, E.; Kucharska, I.; Tan, Y.Z.; Benlekbir, S.; Cui, H.; Zhao, T.; Wasney, G.A.; Budylowski, P.; Guvenc, F.; Newton, J.C.; et al. Multivalency Transforms SARS-CoV-2 Antibodies into Ultrapotent Neutralizers. Nat. Commun. 2021, 12, 3661. [Google Scholar] [CrossRef]
- Hussack, G.; Arbabi-Ghahroudi, M.; van Faassen, H.; Songer, J.G.; Ng, K.K.; MacKenzie, R.; Tanha, J. Neutralization of Toxin A with Single-domain Antibodies Targeting the Cell Receptor Binding Domain. J. Biol. Chem. 2011, 286, 8961–8976. [Google Scholar] [CrossRef]
- Poulain, A.; Perret, S.; Malenfant, F.; Mullick, A.; Massie, B.; Durocher, Y. Rapid Protein Production from Stable CHO Cell Pools using Plasmid Vector and the Cumate Gene-Switch. J. Biotechnol. 2017, 255, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Rocheleau, L.; Laroche, G.; Fu, K.; Stewart, C.M.; Mohamud, A.O.; Cote, M.; Giguere, P.M.; Langlois, M.A.; Pelchat, M. Identification of a High-Frequency Intrahost SARS-CoV-2 Spike Variant with Enhanced Cytopathic and Fusogenic Effects. mBio 2021, 12, e0078821. [Google Scholar] [CrossRef]
- Welsh, J.A.; Jones, J.C. Small Particle Fluorescence and Light Scatter Calibration Using FCMPASS Software. Curr. Protoc. Cytom. 2020, 94, e79. [Google Scholar] [CrossRef]
- Welsh, J.A.; Jones, J.C.; Tang, V.A. Fluorescence and Light Scatter Calibration Allow Comparisons of Small Particle Data in Standard Units across Different Flow Cytometry Platforms and Detector Settings. Cytometry A. 2020, 97, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.A.; Van Der Pol, E.; Arkesteijn, G.J.A.; Bremer, M.; Brisson, A.; Coumans, F.; Dignat-George, F.; Duggan, E.; Ghiran, I.; Giebel, B.; et al. MIFlowCyt-EV: A Framework for Standardized Reporting of Extracellular Vesicle Flow Cytometry Experiments. J. Extracell Vesicles 2020, 9, 1713526. [Google Scholar] [CrossRef]
- Rossotti, M.A.; Henry, K.A.; van Faassen, H.; Tanha, J.; Callaghan, D.; Hussack, G.; Arbabi-Ghahroudi, M.; MacKenzie, C.R. Camelid Single-Domain Antibodies Raised by DNA Immunization are Potent Inhibitors of EGFR signaling. Biochem. J. 2019, 476, 39–50. [Google Scholar] [CrossRef]
- Gettemans, J. Site-Specific Fluorescent Labeling, Single-Step Immunocytochemistry, and Delivery of Nanobodies into Living Cells. Methods Mol. Biol. 2022, 2446, 373–393. [Google Scholar] [CrossRef]
- Pleiner, T.; Bates, M.; Trakhanov, S.; Lee, C.T.; Schliep, J.E.; Chug, H.; Bohning, M.; Stark, H.; Urlaub, H.; Gorlich, D. Nanobodies: Site-Specific Labeling for Super-Resolution Imaging, Rapid Epitope-Mapping and Native Protein Complex Isolation. Elife 2015, 4, e11349. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, M.P.; Pommie, C.; Ruiz, M.; Giudicelli, V.; Foulquier, E.; Truong, L.; Thouvenin-Contet, V.; Lefranc, G. IMGT Unique Numbering for Immunoglobulin and T cell Receptor Variable Domains and Ig Superfamily V-like Domains. Dev. Comp. Immunol. 2003, 27, 55–77. [Google Scholar] [CrossRef]
- Hansen, S.B.; Andersen, K.R. Introducing Cysteines into Nanobodies for Site-Specific Labeling. Methods Mol. Biol. 2022, 2446, 327–343. [Google Scholar] [CrossRef]
- Brittain, G.C.t.; Chen, Y.Q.; Martinez, E.; Tang, V.A.; Renner, T.M.; Langlois, M.A.; Gulnik, S. A Novel Semiconductor-Based Flow Cytometer with Enhanced Light-Scatter Sensitivity for the Analysis of Biological Nanoparticles. Sci. Rep. 2019, 9, 16039. [Google Scholar] [CrossRef] [PubMed]
- Tang, V.A.; Renner, T.M.; Fritzsche, A.K.; Burger, D.; Langlois, M.A. Single-Particle Discrimination of Retroviruses from Extracellular Vesicles by Nanoscale Flow Cytometry. Sci. Rep. 2017, 7, 17769. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.L.; Yuan, T.Z.; Chan, K.Y.; Ding, L.; Han, Z.; Franco, H.; Holliday, C.; Kannan, S.; Davidson, E.; Doranz, B.J.; et al. A VHH Single-Domain Platform Enabling Discovery and Development of Monospecific Antibodies and Modular Neutralizing Bispecifics Against SARS-CoV-2 Variants. Antib. Ther. 2024, 7, 164–176. [Google Scholar] [CrossRef]
- Titong, A.; Gallolu Kankanamalage, S.; Dong, J.; Huang, B.; Spadoni, N.; Wang, B.; Wright, M.; Pham, K.L.J.; Le, A.H.; Liu, Y. First-in-Class Trispecific VHH-Fc Based Antibody with Potent Prophylactic and Therapeutic Efficacy Against SARS-CoV-2 and Variants. Sci. Rep. 2022, 12, 4163. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Format of Nanobody | |||
|---|---|---|---|
| Spike Protein Domain/Subunit | Nanobody Name | Monovalent VHH SPR (KD, nM) | Bivalent VHH-Fc Flow Cytometry (EC50, nM) |
| RBD | VHH-72 2 | 86.2 | 0.2 |
| 07 | 0.94 | 0.3 | |
| 02 | 0.62 | 1 | |
| NTD | SR01 | 0.56 | 3.4 |
| S2 | S2A4 | 12.8 | 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maltseva, M.; Rossotti, M.A.; Tanha, J.; Langlois, M.-A. Characterization of Nanobody Binding to Distinct Regions of the SARS-CoV-2 Spike Protein by Flow Virometry. Viruses 2025, 17, 571. https://doi.org/10.3390/v17040571
Maltseva M, Rossotti MA, Tanha J, Langlois M-A. Characterization of Nanobody Binding to Distinct Regions of the SARS-CoV-2 Spike Protein by Flow Virometry. Viruses. 2025; 17(4):571. https://doi.org/10.3390/v17040571
Chicago/Turabian StyleMaltseva, Mariam, Martin A. Rossotti, Jamshid Tanha, and Marc-André Langlois. 2025. "Characterization of Nanobody Binding to Distinct Regions of the SARS-CoV-2 Spike Protein by Flow Virometry" Viruses 17, no. 4: 571. https://doi.org/10.3390/v17040571
APA StyleMaltseva, M., Rossotti, M. A., Tanha, J., & Langlois, M.-A. (2025). Characterization of Nanobody Binding to Distinct Regions of the SARS-CoV-2 Spike Protein by Flow Virometry. Viruses, 17(4), 571. https://doi.org/10.3390/v17040571