
Diversity of Hepatitis E Viruses in Rats in Yunnan Province and the Inner Mongolia Autonomous Region of China

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Storage
2.2. Sample Pretreatment
2.3. NGS and Full-Length Genome Sequencing of HEV
2.4. Detection of HEV by Nested Broad-Spectrum PCR and Host Species Identification
2.5. Phylogenetic and Sequence Analyses
3. Results
3.1. Overview of NGS Data
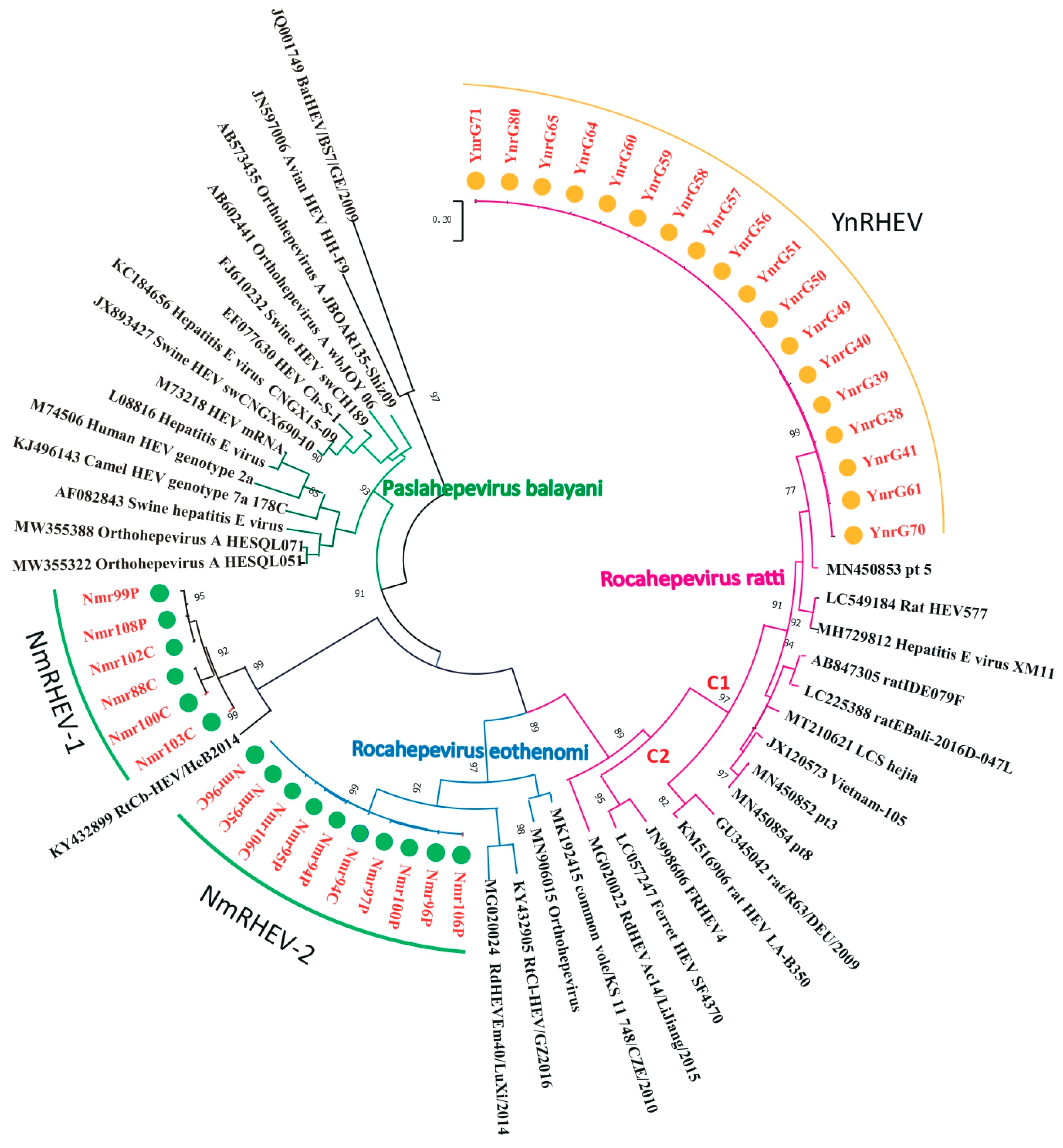
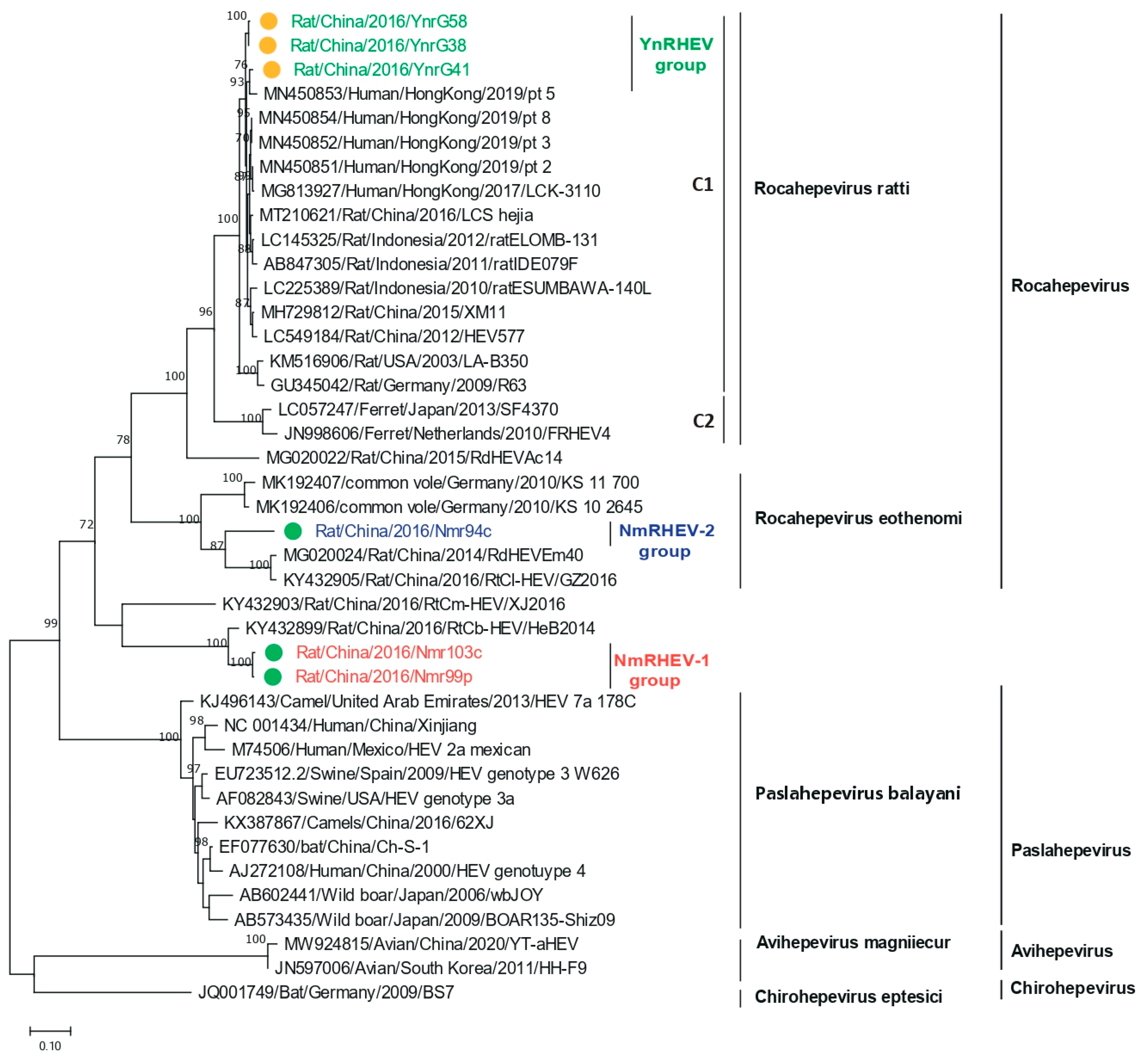
3.2. Prevalence and Phylogenetic Analysis of Hepeviridae in Rat
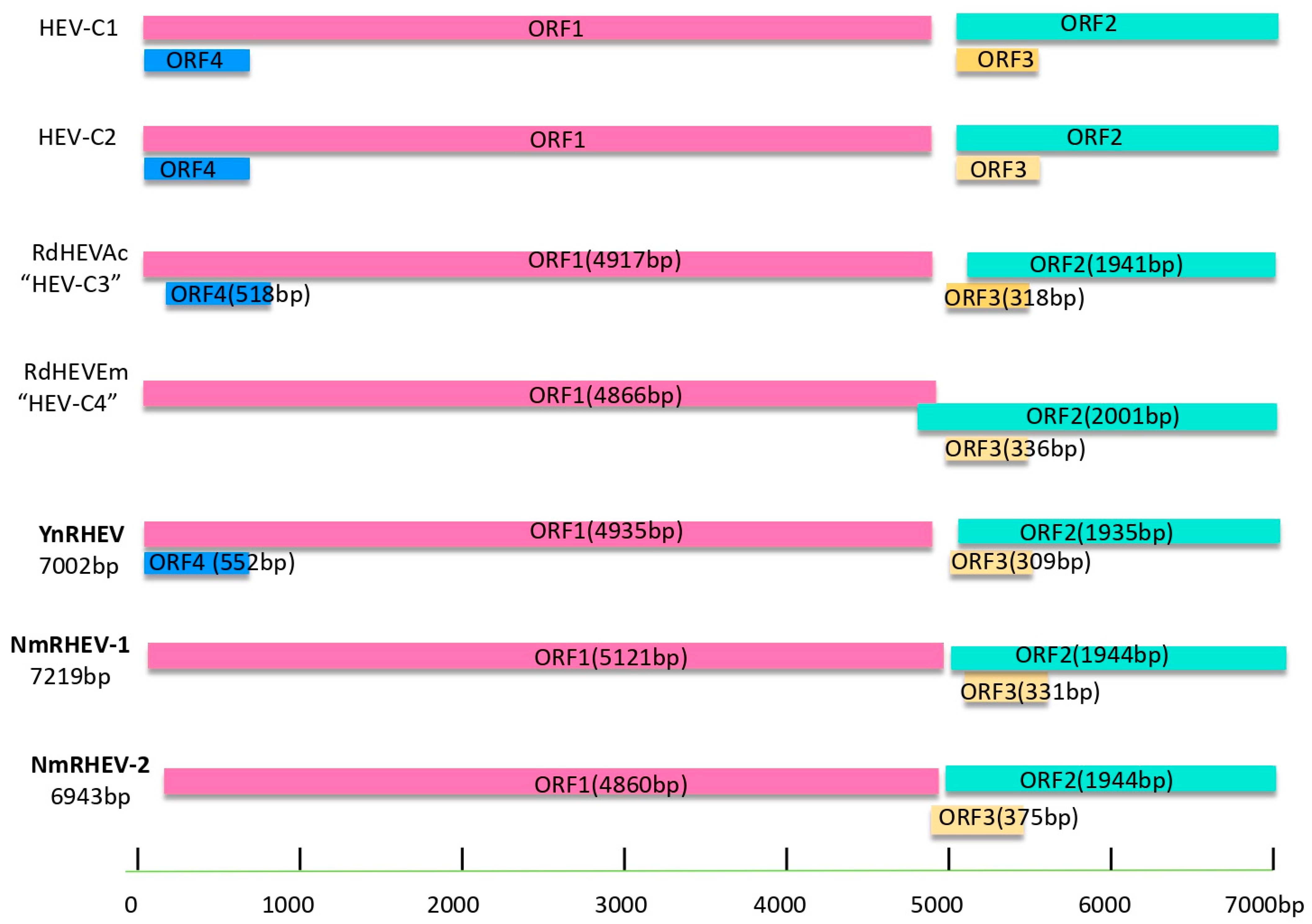
3.3. Characteristics of the HEV Genome
3.4. Comparative Analysis of Complete Genome Sequences
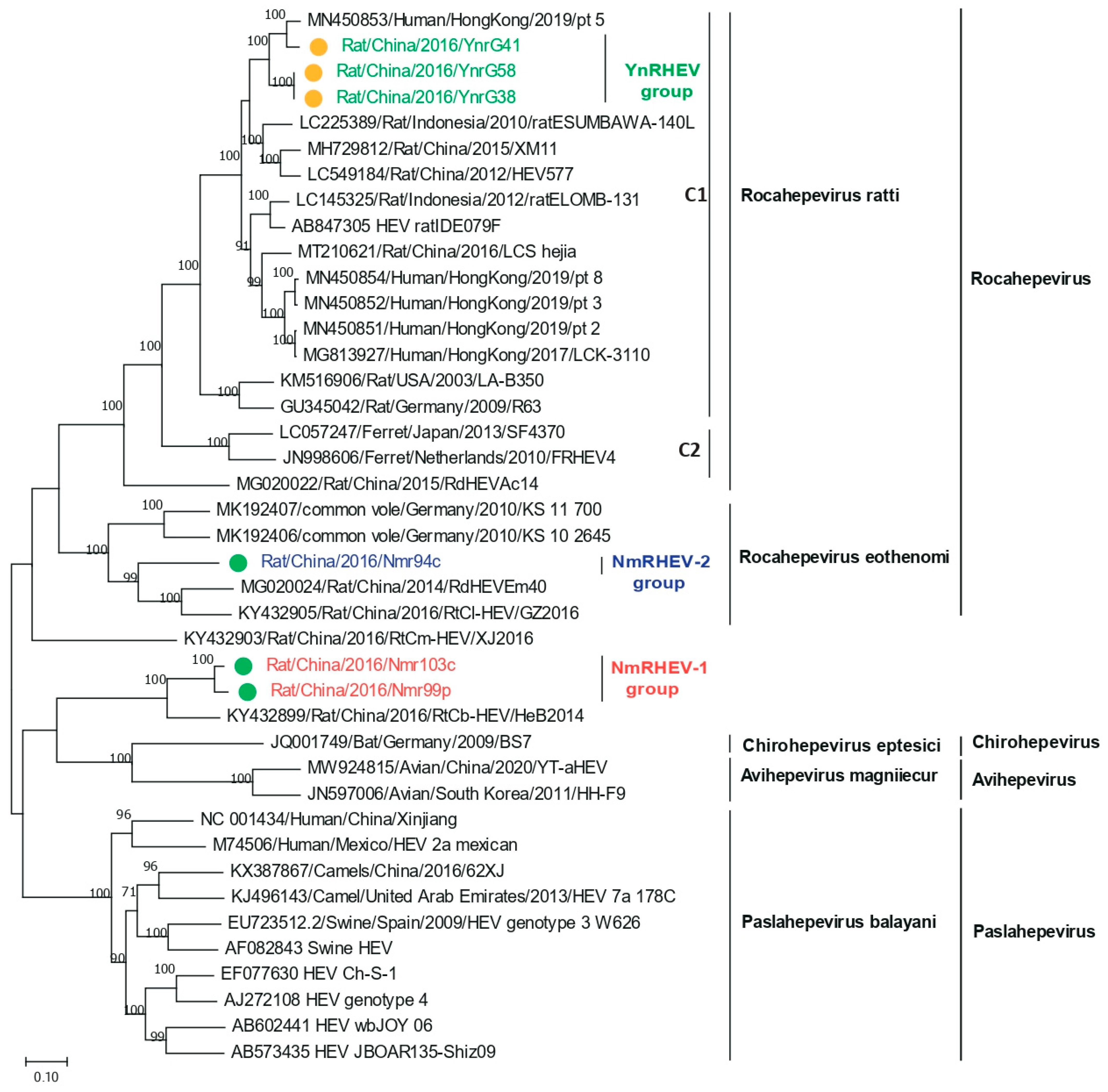
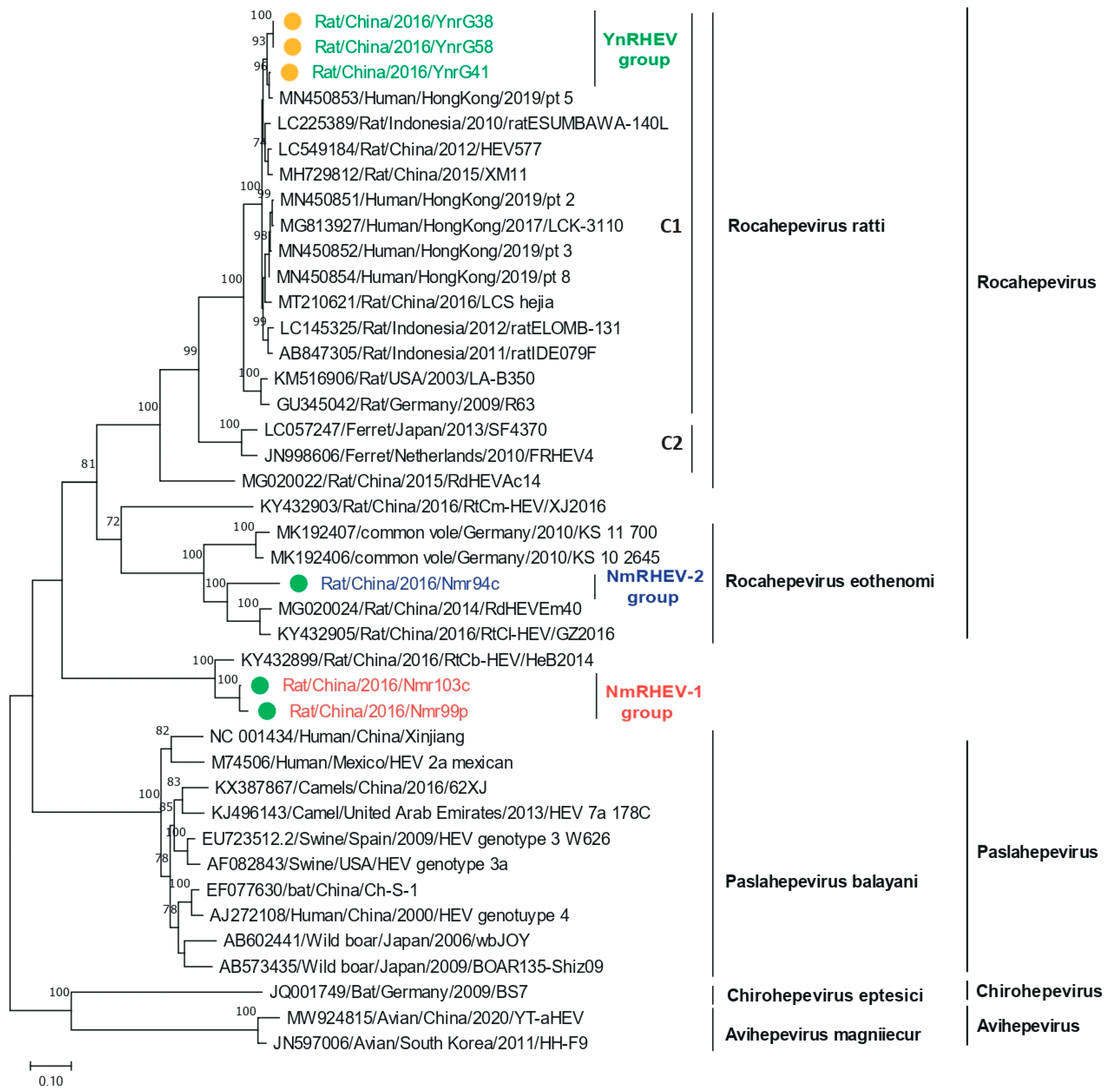
3.5. Phylogenetic Analysis of Complete Genomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, B.; Li, W.; Zhou, J.H.; Li, B.; Zhang, W.; Yang, W.H.; Pan, H.; Wang, L.X.; Bock, C.T.; Shi, Z.L.; et al. Chevrier’s Field Mouse (Apodemus chevrieri) and Père David’s Vole (Eothenomys melanogaster) in China Carry Orthohepeviruses that form Two Putative Novel Genotypes Within the Species Orthohepevirus C. Virol. Sin. 2018, 33, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Feng, Y.; Qu, F.; Zhang, L.; Yan, B.; Lv, J. Prediction of hepatitis E using machine learning models. PLoS ONE 2020, 15, e0237750. [Google Scholar] [CrossRef] [PubMed]
- Nimgaonkar, I.; Ding, Q.; Schwartz, R.E.; Ploss, A. Hepatitis E virus: Advances and challenges. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Kenney, S.P.; Meng, X.-J. Hepatitis E Virus Genome Structure and Replication Strategy. Cold Spring Harb. Perspect. Med. 2019, 9, a031724. [Google Scholar] [CrossRef]
- Wang, B.; Meng, X.-J. Structural and molecular biology of hepatitis E virus. Comput. Struct. Biotechnol. J. 2021, 19, 1907–1916. [Google Scholar] [CrossRef]
- Li, F.-L.; Wang, B.; Han, P.-Y.; Li, B.; Si, H.-R.; Zhu, Y.; Yin, H.-M.; Zong, L.-D.; Tang, Y.; Shi, Z.-L.; et al. Identification of novel rodent and shrew orthohepeviruses sheds light on hepatitis E virus evolution. Zool. Res. 2025, 46, 103–121. [Google Scholar] [CrossRef]
- Purdy, M.A.; Drexler, J.F.; Meng, X.-J.; Norder, H.; Okamoto, H.; Van der Poel, W.H.M.; Reuter, G.; de Souza, W.M.; Ulrich, R.G.; Smith, D.B. ICTV Virus Taxonomy Profile: Hepeviridae 2022. J. Gen. Virol. 2022, 103, 001778. [Google Scholar] [CrossRef]
- Smith, D.B.; Simmonds, P.; Members of the International Committee on the Taxonomy of Viruses Study Group; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.M.; Purdy, M.A. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2014, 95 Pt 10, 2223–2232. [Google Scholar] [CrossRef]
- Reuter, G.; Boros, Á.; Pankovics, P. Review of Hepatitis E Virus in Rats: Evident Risk of Species Orthohepevirus C to Human Zoonotic Infection and Disease. Viruses 2020, 12, 1148. [Google Scholar] [CrossRef]
- Wang, L.; Teng, J.L.L.; Lau, S.K.P.; Sridhar, S.; Fu, H.; Gong, W.; Li, M.; Xu, Q.; He, Y.; Zhuang, H.; et al. Transmission of a Novel Genotype of Hepatitis E Virus from Bactrian Camels to Cynomolgus Macaques. J. Virol. 2019, 93, 10–1128. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Cao, K.Y.; Wernery, U.; Schountz, T.; Chiu, T.H.; Tsang, A.K.; Wong, P.C.; Wong, E.Y.; et al. New Hepatitis E Virus Genotype in Bactrian Camels, Xinjiang, China, 2013. Emerg. Infect. Dis. 2016, 22, 2219–2221. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wen, Y.; Xiong, Y.; Zhang, M.; Cheng, M.; Chen, Q. The prevalence and genomic characteristics of hepatitis E virus in murine rodents and house shrews from several regions in China. BMC Vet. Res. 2018, 14, 414. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Cai, C.L.; Li, B.; Zhang, W.; Zhu, Y.; Chen, W.H.; Zhuo, F.; Shi, Z.L.; Yang, X.L. Detection and characterization of three zoonotic viruses in wild rodents and shrews from Shenzhen city, China. Virol. Sin. 2017, 32, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Harms, D.; Yang, X.-L.; Bock, C.T. Orthohepevirus C: An Expanding Species of Emerging Hepatitis E Virus Variants. Pathogens 2020, 9, 154. [Google Scholar] [CrossRef]
- Reuter, G.; Boros, Á.; Mátics, R.; Kapusinszky, B.; Delwart, E.; Pankovics, P. Divergent hepatitis E virus in birds of prey, common kestrel (Falco tinnunculus) and red-footed falcon (F. vespertinus), Hungary. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 43, 343–346. [Google Scholar] [CrossRef]
- Lhomme, S.; Top, S.; Bertagnoli, S.; Dubois, M.; Guerin, J.L.; Izopet, J. Wildlife Reservoir for Hepatitis E Virus, Southwestern France. Emerg. Infect. Dis. 2015, 21, 1224–1226. [Google Scholar] [CrossRef]
- Johne, R.; Heckel, G.; Plenge-Bönig, A.; Kindler, E.; Maresch, C.; Reetz, J.; Schielke, A.; Ulrich, R.G. Novel hepatitis E virus genotype in Norway rats, Germany. Emerg. Infect. Dis. 2010, 16, 1452–1455. [Google Scholar] [CrossRef]
- Johne, R.; Dremsek, P.; Reetz, J.; Heckel, G.; Hess, M.; Ulrich, R.G. Hepeviridae: An expanding family of vertebrate viruses. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2014, 27, 212–229. [Google Scholar] [CrossRef]
- Murphy, E.G.; Williams, N.J.; Jennings, D.; Chantrey, J.; Verin, R.; Grierson, S.; McElhinney, L.M.; Bennett, M. First detection of Hepatitis E virus (Orthohepevirus C) in wild brown rats (Rattus norvegicus) from Great Britain. Zoonoses Public Health 2019, 66, 686–694. [Google Scholar] [CrossRef]
- Khuroo, M.S.; Khuroo, M.S.; Khuroo, N.S. Hepatitis E: Discovery, global impact, control and cure. World J. Gastroenterol. 2016, 22, 7030–7045. [Google Scholar] [CrossRef]
- de Souza, W.M.; Romeiro, M.F.; Sabino-Santos, G., Jr.; Maia, F.G.M.; Fumagalli, M.J.; Modha, S.; Nunes, M.R.T.; Murcia, P.R.; Figueiredo, L.T.M. Novel orthohepeviruses in wild rodents from Sao Paulo State, Brazil. Virology 2018, 519, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Ryll, R.; Bernstein, S.; Heuser, E.; Schlegel, M.; Dremsek, P.; Zumpe, M.; Wolf, S.; Pépin, M.; Bajomi, D.; Müller, G.; et al. Detection of rat hepatitis E virus in wild Norway rats (Rattus norvegicus) and Black rats (Rattus rattus) from 11 European countries. Vet. Microbiol. 2017, 208, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Purcell, R.H.; Engle, R.E.; Rood, M.P.; Kabrane-Lazizi, Y.; Nguyen, H.T.; Govindarajan, S.; St Claire, M.; Emerson, S.U. Hepatitis E virus in rats, Los Angeles, California, USA. Emerg. Infect. Dis. 2011, 17, 2216–2222. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Yip, C.C.; Lo, K.H.; Wu, S.; Situ, J.; Chew, N.F.; Leung, K.H.; Chan, H.S.; Wong, S.C.; Leung, A.W.; et al. Hepatitis E virus species C infection in humans, Hong Kong. Clin. Infect. Dis. 2021, 75, 288–296. [Google Scholar] [CrossRef]
- Sridhar, S.; Yip, C.C.Y.; Wu, S.; Cai, J.; Zhang, A.J.; Leung, K.H.; Chung, T.W.H.; Chan, J.F.W.; Chan, W.M.; Teng, J.L.L.; et al. Rat Hepatitis E Virus as Cause of Persistent Hepatitis after Liver Transplant. Emerg. Infect. Dis. 2018, 24, 2241–2250. [Google Scholar] [CrossRef]
- Johne, R.; Dremsek, P.; Kindler, E.; Schielke, A.; Plenge-Bönig, A.; Gregersen, H.; Wessels, U.; Schmidt, K.; Rietschel, W.; Groschup, M.H.; et al. Rat hepatitis E virus: Geographical clustering within Germany and serological detection in wild Norway rats (Rattus norvegicus). Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2012, 12, 947–956. [Google Scholar] [CrossRef]
- Sridhar, S.; Yip, C.C.; Wu, S.; Chew, N.F.; Leung, K.H.; Chan, J.F.; Zhao, P.S.; Chan, W.M.; Poon, R.W.; Tsoi, H.W.; et al. Transmission of Rat Hepatitis E Virus Infection to Humans in Hong Kong: A Clinical and Epidemiological Analysis. Hepatology 2021, 73, 10–22. [Google Scholar] [CrossRef]
- Andonov, A.; Robbins, M.; Borlang, J.; Cao, J.; Hatchette, T.; Stueck, A.; Deschambault, Y.; Murnaghan, K.; Varga, J.; Johnston, L. Rat Hepatitis E Virus Linked to Severe Acute Hepatitis in an Immunocompetent Patient. J. Infect. Dis. 2019, 220, 951–955. [Google Scholar] [CrossRef]
- Li, L.L.; Liu, M.M.; Shen, S.; Zhang, Y.J.; Xu, Y.L.; Deng, H.Y.; Deng, F.; Duan, Z.J. Detection and characterization of a novel hepacivirus in long-tailed ground squirrels (Spermophilus undulatus) in China. Arch. Virol. 2019, 164, 2401–2410. [Google Scholar] [CrossRef]
- Johne, R.; Plenge-Bönig, A.; Hess, M.; Ulrich, R.G.; Reetz, J.; Schielke, A. Detection of a novel hepatitis E-like virus in faeces of wild rats using a nested broad-spectrum RT-PCR. J. Gen. Virol. 2010, 91 Pt 3, 750–758. [Google Scholar] [CrossRef]
- Nicolas, V.; Querouil, S.; Verheyen, E.; Verheyen, W.; Mboumba, J.F.; Dillen, M.; Colyn, M. Mitochondrial phylogeny of African wood mice, genus Hylomyscus (Rodentia, Muridae): Implications for their taxonomy and biogeography. Mol. Phylogenetics Evol. 2006, 38, 779–793. [Google Scholar] [CrossRef]
- Sooryanarain, H.; Meng, X.J. Hepatitis E virus: Reasons for emergence in humans. Curr. Opin. Virol. 2019, 34, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.J. Expanding Host Range and Cross-Species Infection of Hepatitis E Virus. PLoS Pathog. 2016, 12, e1005695. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Juarez, A.; Frias, M.; Perez, A.B.; Pineda, J.A.; Reina, G.; Fuentes-Lopez, A.; Freyre-Carrillo, C.; Ramirez-Arellano, E.; Alados, J.C.; Rivero, A.; et al. Orthohepevirus C infection as an emerging cause of acute hepatitis in Spain: First report in Europe. J. Hepatol. 2022, 77, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Hao, X.; Xia, Y.; Yu, W.; Huang, F. Rat Hepatitis E virus is a potential zoonotic pathogen to humans. J. Hepatol. 2022, 77, 868–870. [Google Scholar] [CrossRef]
- Wang, B.; Meng, X.J. Hepatitis E virus: Host tropism and zoonotic infection. Curr. Opin. Microbiol. 2021, 59, 8–15. [Google Scholar] [CrossRef]
- Su, Q.; Chen, Y.; Wang, B.; Huang, C.; Han, S.; Yuan, G.; Zhang, Q.; He, H. Epidemiology and genetic diversity of zoonotic pathogens in urban rats (Rattus spp.) from a subtropical city, Guangzhou, southern China. Zoonoses Public Health 2020, 67, 534–545. [Google Scholar] [CrossRef]
- Li, W.; Guan, D.; Su, J.; Takeda, N.; Wakita, T.; Li, T.-C.; Ke, C.W. High prevalence of rat hepatitis E virus in wild rats in China. Vet. Microbiol. 2013, 165, 275–280. [Google Scholar] [CrossRef]
- Robinson, S.J.; Borlang, J.; Himsworth, C.G.; Pearl, D.L.; Weese, J.S.; Dibernardo, A.; Osiowy, C.; Nasheri, N.; Jardine, C.M. Rat Hepatitis E Virus in Norway Rats, Ontario, Canada, 2018–2021. Emerg. Infect. Dis. 2023, 29, 1890–1894. [Google Scholar] [CrossRef]
- Takahashi, M.; Kunita, S.; Kawakami, M.; Kadosaka, T.; Fujita, H.; Takada, N.; Miyake, M.; Kobayashi, T.; Ohnishi, H.; Nagashima, S.; et al. First detection and characterization of rat hepatitis E Virus (HEV-C1) in Japan. Virus Res. 2022, 314, 198766. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Sample ID | Site | Date | Host Species | Virus Classification |
|---|---|---|---|---|---|
| YnRHEV | 16Ynr-38 | YN | June 2016 | Rattus norvegicus | Rocahepevirus ratti |
| 16Ynr-39 | YN | June 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-40 | YN | June 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-41 | YN | June 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-49 | YN | September 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-50 | YN | September 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-51 | YN | September 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-56 | YN | September 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-57 | YN | September 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-58 | YN | September 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-59 | YN | September 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-60 | YN | September 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-61 | YN | October 2013 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-64 | YN | November 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-65 | YN | November 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-70 | YN | November 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-71 | YN | November 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| 16Ynr-80 | YN | November 2016 | Rattus norvegicus | Rocahepevirus ratti | |
| NmRHEV-1 | 17Nmr-103C | IMAR | October 2017 | Apodemus agrarius | Novel orthohepeviruses |
| 17Nmr-99P | IMAR | June 2017 | Cricetulus sokolovi | Novel orthohepeviruses | |
| 17Nmr-100C | IMAR | October 2017 | Cricetulus barabensis | Novel orthohepeviruses | |
| 17Nmr-88C | IMAR | June 2017 | Mus musculus | Novel orthohepeviruses | |
| 17Nmr-102C | IMAR | October 2017 | Cricetulus barabensis | Novel orthohepeviruses | |
| 17Nmr-108P | IMAR | October 2017 | unidentified | Novel orthohepeviruses | |
| NmRHEV-2 | 17Nmr-96C | IMAR | June 2017 | Microtus fortis | Rocahepevirus Eothenomi |
| 17Nmr-95C | IMAR | June 2017 | Microtus fortis | Rocahepevirus Eothenomi | |
| 17Nmr-95P | IMAR | June 2017 | Microtus fortis | Rocahepevirus Eothenomi | |
| 17Nmr-94P | IMAR | June 2017 | Microtus fortis | Rocahepevirus Eothenomi | |
| 17Nmr-106C | IMAR | October 2017 | Microtus fortis | Rocahepevirus Eothenomi | |
| 17Nmr-94C | IMAR | June 2017 | Microtus fortis | Rocahepevirus Eothenomi | |
| 17Nmr-97P | IMAR | June 2017 | Microtus fortis | Rocahepevirus Eothenomi | |
| 17Nmr-100P | IMAR | October 2017 | Cricetulus barabensis | Rocahepevirus Eothenomi | |
| 17Nmr-96P | IMAR | June 2017 | Microtus fortis | Rocahepevirus Eothenomi | |
| 17Nmr-106P | IMAR | October 2017 | Microtus fortis | Rocahepevirus Eothenomi |
| Nucleotide Sequences (%) | Amino Acid Sequences (%) | ||||||
|---|---|---|---|---|---|---|---|
| Genotype/Strain | Complete | ORF1 | ORF2 | ORF3 | ORF1 | ORF2 | ORF3 |
| YnRHEV | |||||||
| YnrG38/G58/G41 | 89.3–99.0 | 88.8–99.0 | 84.6–98.1 | 93.8–100 | 92.6–96.3 | 97.6 | 94.5 |
| NmRHEV-1 | 50.2–50.5 | 45.5–48.5 | 57.2–57.6 | 46.4–48.8 | 48.9–51.6 | 56.7–57.3 | 45–46.1 |
| NmRHEV-2 | 54.6–56.7 | 48.9–49.0 | 58.9–62.1 | 48.9–51.2 | 54.6–56.7 | 57.3–59.2 | 45.2–65.3 |
| HEV-C1 | 88.9–94.3 | 88.6–94.3 | 90.1–94.4 | 94.4–97.4 | 90.1–98.0 | 96.8–97.5 | 94.9 |
| HEV-C2 | 63.2–63.6 | 64.5–64.8 | 71.9–72 | 58.5–59.7 | 73.6–77.4 | 80.4–80.7 | 62.5–63.4 |
| HEV-C3 | 64.5–64.8 | 61.3–61.8 | 68.1–68.2 | 58.7 | 67.6–70.5 | 74.2 | 59.0–59.6 |
| HEV-C4 | 61.3–61.8 | 50.1–50.2 | 61.5–62.0 | 39.7–39.8 | 54.5–56.8 | 61.5–61.8 | 41.6–43.1 |
| HEV-A | 52.1–53.0 | 49.4–50.8 | 56.1–56.4 | 47.9–51.3 | 48.4–51.4 | 55.8–56.6 | 20.1–21.4 |
| HEV-B | 45.8–46.4 | 45.9–46.1 | 47.2–47.3 | 34.1–35.0 | 47.1–49.4 | 43.4–44.0 | 25.9–26.3 |
| HEV-D | 46.5–46.8 | 45.0–45.1 | 49.9–50.3 | 35.6–36.2 | 47.6–49.3 | 48.8–49.1 | 11.8–13.1 |
| NmRHEV-1 | |||||||
| Nmr103c/Nmr99p | 95.4 | 94.8 | 96.9 | 98.5 | 97.8 | 99.2 | 98.1 |
| NmRHEV-2 | 51.8–52.3 | 47.7–47.9 | 58.1–59.2 | 52.1–52.9 | 51.8–52.3 | 56.2–58.3 | 48.2–49.5 |
| RtCb-HEV/HeB2014 | 77.6–77.7 | 75.5–75.8 | 83.2–83.6 | 83.9–84.6 | 87.5–88.5 | 91.3–91.4 | 92.3 |
| HEV-C1 | 50.3–50.6 | 48.1–48.2 | 56.5–56.8 | 43.1–43.8 | 48.3–49.0 | 56.6–56.7 | 36.3 |
| HEV-C2 | 51.3–51.3 | 47.3–47.5 | 56.5–57 | 37.2–37.6 | 48.7–49.2 | 57.1–57.2 | 34.1 |
| HEV-C3 | 51.7 | 47.8 | 56.3–56.4 | 42.9–43.6 | 48–48.5 | 55.0–55.1 | 32.6 |
| HEV-C4 | 52.6 | 54.2–54.4 | 58.1–58.3 | 37.6–38.1 | 47.6–48.5 | 59.1–59.3 | 26.5 |
| HEV-A | 52.8–53.5 | 52.5–53.4 | 53.9–54.9 | 35.6–38.2 | 44.3–49.2 | 54.6–55.7 | 21.5–24.4 |
| HEV-B | 43.4–43.7 | 42.4–42.6 | 46.3–47.0 | 27.9–38.3 | 41.9–42.1 | 42.2–42.3 | 22.2 |
| HEV-D | 46.1 | 44.9–45.0 | 48.9–49.0 | 30.2–30.6 | 41.4–41.8 | 49.2–49.5 | 21.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.-L.; Ma, X.-H.; Nan, X.-W.; Wang, J.-L.; Zhao, J.; Sun, X.-M.; Li, J.-S.; Zheng, G.-S.; Duan, Z.-J. Diversity of Hepatitis E Viruses in Rats in Yunnan Province and the Inner Mongolia Autonomous Region of China. Viruses 2025, 17, 490. https://doi.org/10.3390/v17040490
Li L-L, Ma X-H, Nan X-W, Wang J-L, Zhao J, Sun X-M, Li J-S, Zheng G-S, Duan Z-J. Diversity of Hepatitis E Viruses in Rats in Yunnan Province and the Inner Mongolia Autonomous Region of China. Viruses. 2025; 17(4):490. https://doi.org/10.3390/v17040490
Chicago/Turabian StyleLi, Li-Li, Xiao-Hua Ma, Xiao-Wei Nan, Jing-Lin Wang, Jing Zhao, Xiao-Man Sun, Jin-Song Li, Gui-Sen Zheng, and Zhao-Jun Duan. 2025. "Diversity of Hepatitis E Viruses in Rats in Yunnan Province and the Inner Mongolia Autonomous Region of China" Viruses 17, no. 4: 490. https://doi.org/10.3390/v17040490
APA StyleLi, L.-L., Ma, X.-H., Nan, X.-W., Wang, J.-L., Zhao, J., Sun, X.-M., Li, J.-S., Zheng, G.-S., & Duan, Z.-J. (2025). Diversity of Hepatitis E Viruses in Rats in Yunnan Province and the Inner Mongolia Autonomous Region of China. Viruses, 17(4), 490. https://doi.org/10.3390/v17040490