
Identification of B-Cell Epitopes Located on the Surface of the S1 Protein of Infectious Bronchitis Virus M41 Strains


Abstract
1. Introduction
2. Materials and Methods
2.1. Virus and Cells
2.2. S1 Gene Cloning and Recombinant Protein Expression
2.3. Immunization of Mice
2.4. Preparation and Identification of Monoclonal Antibodies
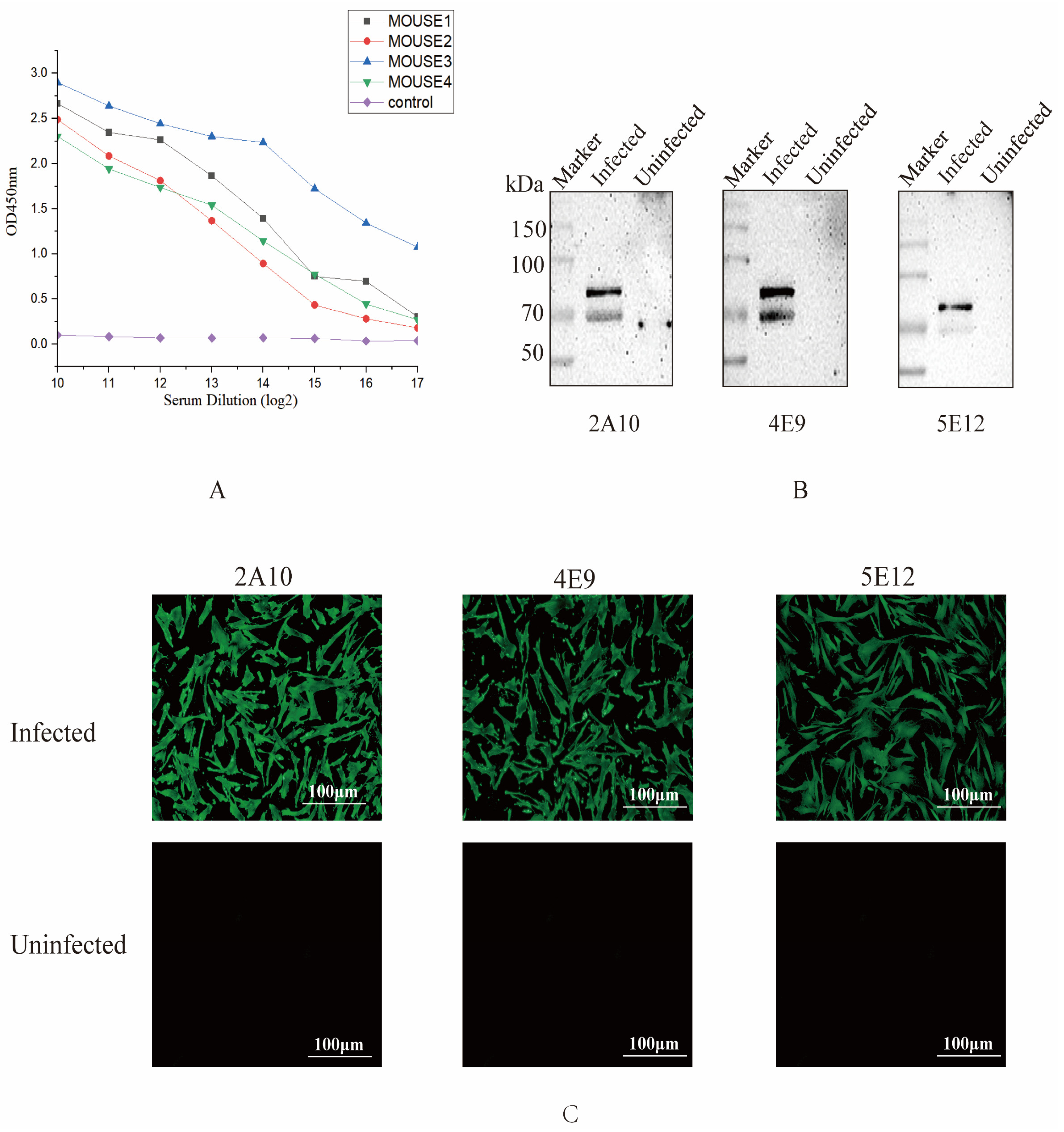
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Western Blot Analysis
2.7. Indirect Immunofluorescence Assay (IFA)
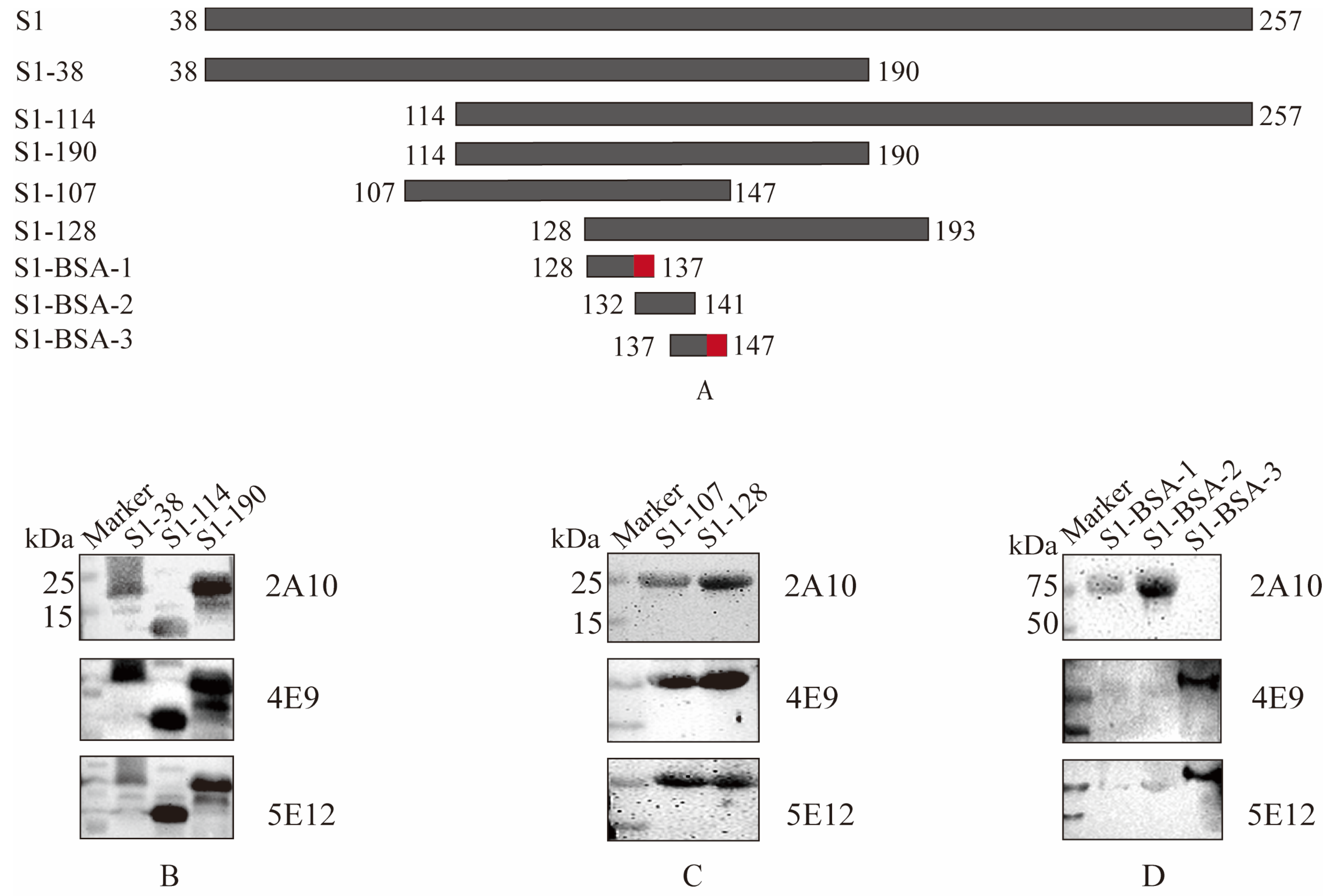
2.8. Overlapping Truncated S1 Gene Design and B-Cell Epitope Identification
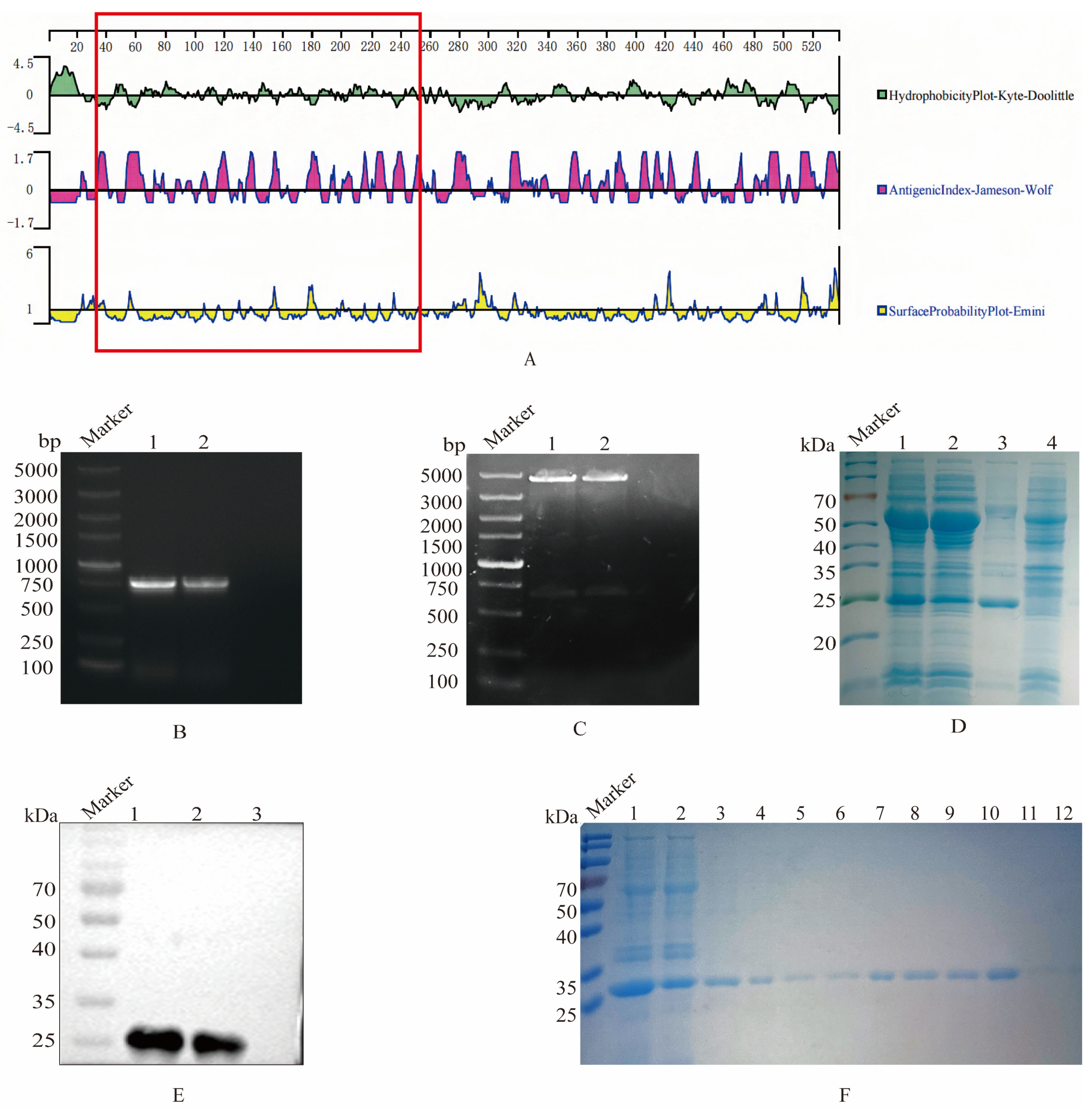
2.9. Bioinformatics Analysis
3. Results
3.1. Cloning, Expression and Purification of Recombinant S1 Protein
3.2. Screening and Characterization of Monoclonal Antibodies
3.3. Identification of B-Cell Epitopes of Monoclonal Antibodies
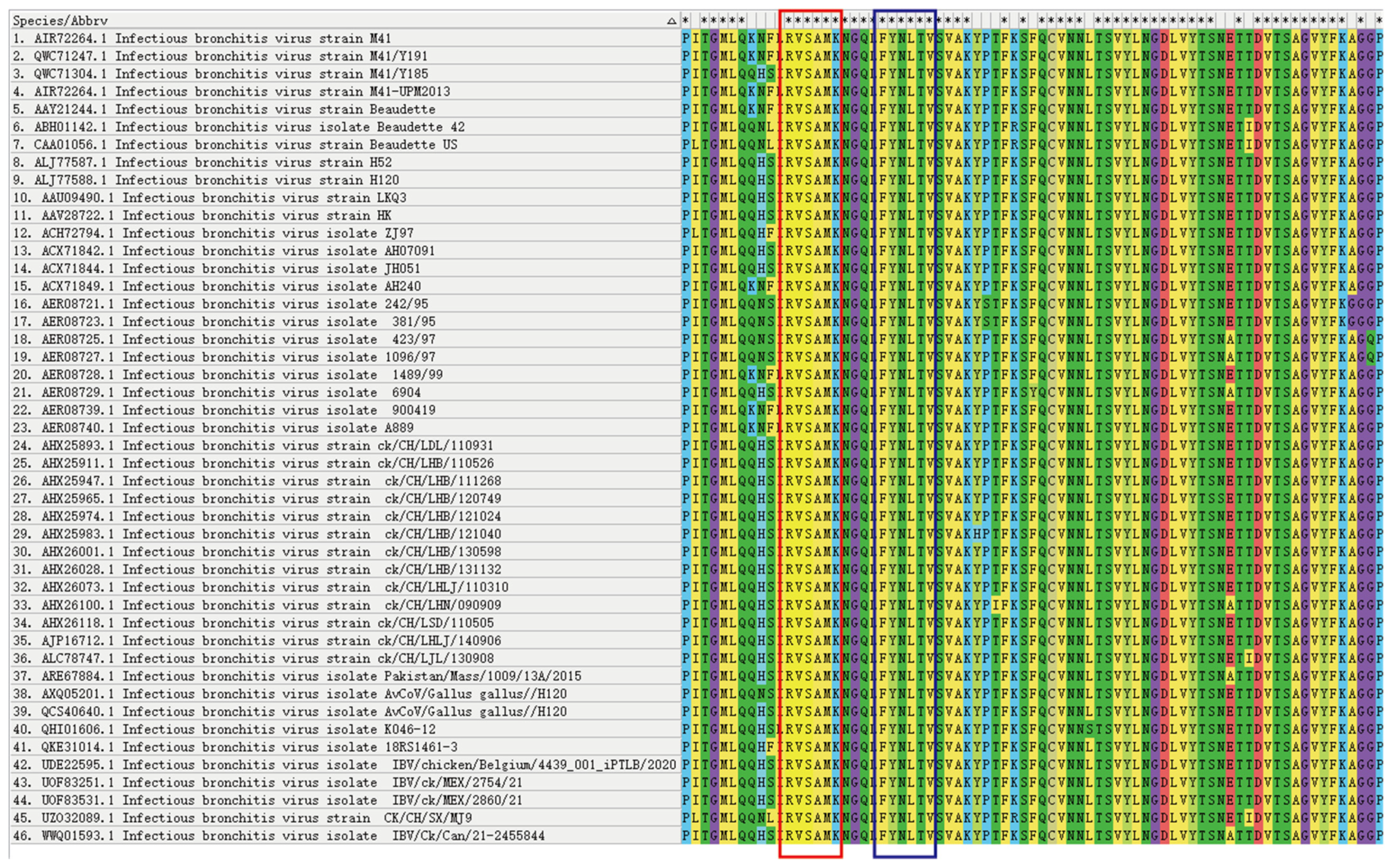
3.4. The Identified Epitopes Are Highly Conserved in IBV Strains
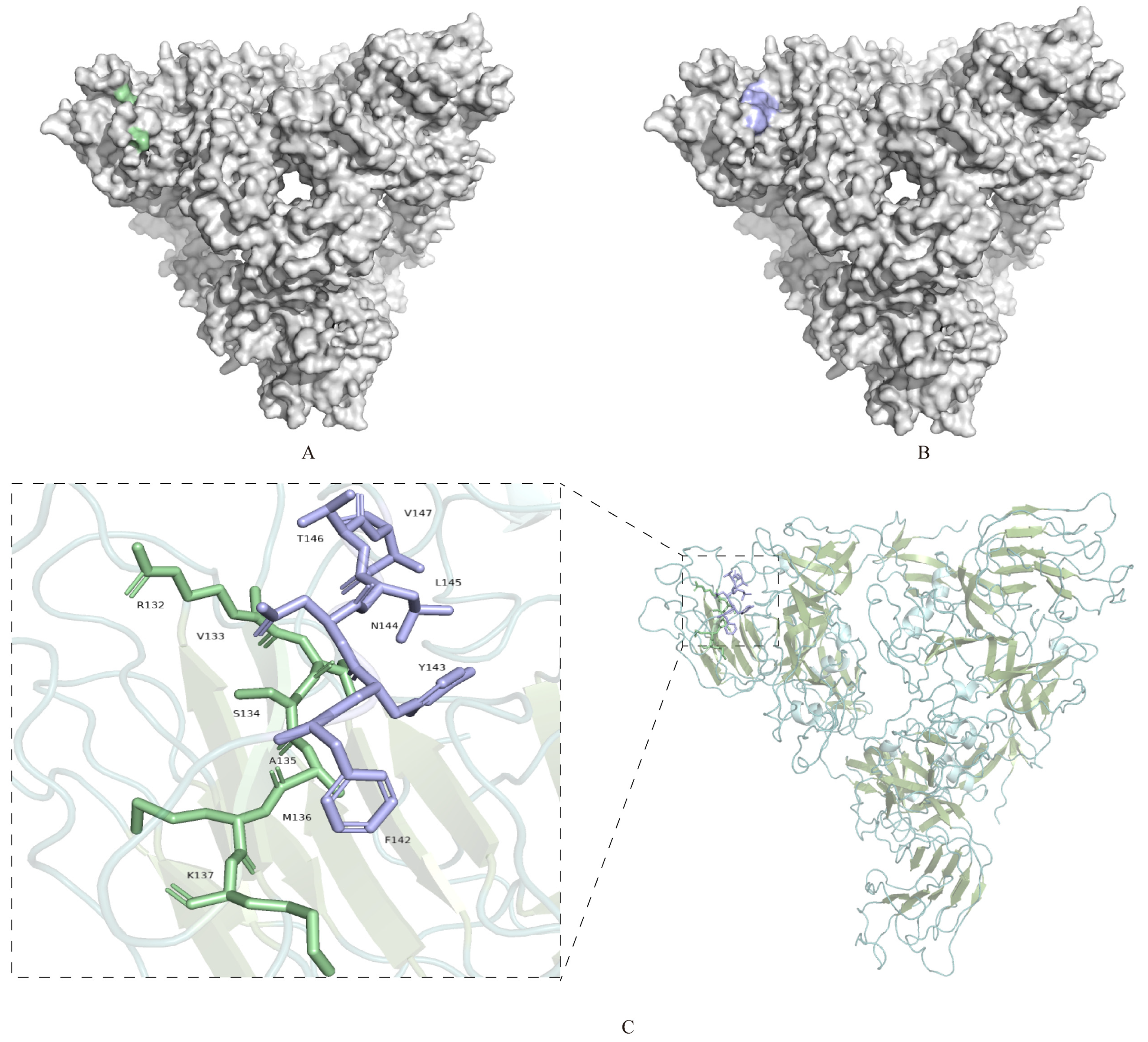
3.5. Spatial Location Prediction of Epitope Binding
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [PubMed]
- Abolnik, C.; Phiri, T.P.; van der Zel, G.; Anthony, J.; Daniell, N.; de Boni, L. Wild bird surveillance in the Gauteng Province of South Africa during the high-risk period for highly pathogenic avian influenza virus introduction. Viruses 2022, 14, 2027. [Google Scholar] [CrossRef] [PubMed]
- Andreychev, A.; Boyarova, E.; Brandler, O.; Tukhbatullin, A.; Kapustina, S. Terrestrial and subterranean mammals as reservoirsof zoonotic diseases in the central part of European Russia. Diversity 2023, 15, 39. [Google Scholar]
- Schalk, A.F.; Hawn, M.C. An apparently new respiratory disease of baby chicks. J. Am. Vet. Med. Assoc. 1931, 78, 19. [Google Scholar]
- Beach, J.R.; Schalm, O.W. A filterable virus, distinct from that of laryngotracheitis, the cause of a respiratory disease of chicks. Poult. Sci. 1936, 15, 199–206. [Google Scholar]
- Zhang, X.; Guo, M.; Zhao, J.; Wu, Y. Avian Infectious Bronchitis in China: Epidemiology, Vaccination, and Control. Avian Dis. 2021, 65, 652–656. [Google Scholar] [CrossRef]
- Legnardi, M.; Tucciarone, C.M.; Franzo, G.; Cecchinato, M. Infectious Bronchitis Virus Evolution, Diagnosis and Control. Vet. Sci. 2020, 7, 79. [Google Scholar] [CrossRef]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 39, 349–364. [Google Scholar] [CrossRef]
- Masters, P.S. The molecular biology of coronaviruses. Adv. Virus Res. 2006, 66, 193–292. [Google Scholar]
- Yuan, H.W.; Wen, H.L. Research progress on coronavirus S proteins and their receptors. Arch. Virol. 2021, 166, 1811–1817. [Google Scholar] [CrossRef]
- Bouwman, K.M.; Habraeken, N.; Laconi, A.; Berends, A.J.; Groenewoud, L.; Alders, M.; Kemp, V.; Verheije, M.H. N-glycosylation of infectious bronchitis virus M41 spike determines receptor specificity. J. Gen. Virol. 2020, 101, 599–608. [Google Scholar] [PubMed]
- Raman, R.; Tharakaraman, K.; Sasisekharan, V.; Sasisekharan, R. Glycan-protein interactions in viral pathogenesis. Curr. Opin. Struct. Biol. 2016, 40, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Zheng, Y.; Yang, Y.; Liu, C.; Geng, Q.; Luo, C.; Zhang, W.; Li, F. Cryo-EM structure of infectious bronchitis coronavirus spike protein reveals structural and functional evolution of coronavirus spike proteins. PLoS Pathog. 2018, 14, e1007009. [Google Scholar]
- Yamada, Y.; Liu, D.X. Proteolytic Activation of the Spike Protein at a Novel RRRR/S Motif Is Implicated in Furin-Dependent Entry, Syncytium Formation, and Infectivity of Coronavirus Infectious Bronchitis Virus in Cultured Cells. J. Virol. 2009, 83, 8744–8758. [Google Scholar]
- Gombold, J.L.; Hingley, S.T.; Weiss, S.R. Fusion-defective mutants of mouse hepatitis virus A59 contain a mutation in the spike protein cleavage signal. J. Virol. 1993, 67, 4504–4512. [Google Scholar]
- Wickramasinghe, I.N.; de Vries, R.P.; Gröne, A.; de Haan, C.A.; Verheije, M.H. Binding of avian coronavirus spike proteins to host factors reflects virus tropism and pathogenicity. J. Virol. 2011, 85, 8903–8912. [Google Scholar] [CrossRef]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.; Rottier, P.J. The coronavirus spike protein is a class I virus fusion protein: Structural and functional characterization of the fusion core complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef]
- Mou, H.; Raj, V.S.; van Kuppeveld, F.J.; Rottier, P.J.; Haagmans, B.L.; Bosch, B.J. The receptor binding domain of the new Middle East respiratory syndrome coronavirus maps to a 231-residue region in the spike protein that efficiently elicits neutralizing antibodies. J. Virol. 2013, 87, 9379–9383. [Google Scholar]
- You, R.; Liu, K.; Huang, M.; Tang, L.; Zhang, X.; Huang, Y.; Zhao, J.; Zhao, Y.; Ye, L.; Zhang, G. Identification and Comparison of the Sialic Acid-Binding Domain Characteristics of Avian Coronavirus Infectious Bronchitis Virus Spike Protein. J. Virol. 2023, 97, e00489-23. [Google Scholar]
- Parsons, L.M.; Bouwman, K.M.; Azurmendi, H.; de Vries, R.P.; Cipollo, J.F.; Verheije, M.H. Glycosylation of the viral attachment protein of avian coronavirus is essential for host cell and receptor binding. J. Biol. Chem. 2019, 294, 7797–7809. [Google Scholar]
- Zhang, Y.; Xu, Z.; Cao, Y. Host Antiviral Responses against Avian Infectious Bronchitis Virus (IBV): Focus on Innate Immunity. Viruses 2021, 13, 1698. [Google Scholar] [CrossRef] [PubMed]
- Promkuntod, N.; van Eijndhoven, R.E.; de Vrieze, G.; Gröne, A.; Verheije, M.H. Mapping of the receptor-binding domain and amino acids critical for attachment in the spike protein of avian coronavirus infectious bronchitis virus. Virology 2014, 448, 26–32. [Google Scholar]
- Cook, J.K.; Jackwood, M.; Jones, R.C. The long view: 40 years of infectious bronchitis research. Avian Pathol. 2012, 41, 239–250. [Google Scholar]
- Li, S.; Chen, W.; Shen, Y.; Xia, J.; Fan, S.; Li, N.; Luo, Y.; Han, X.; Cui, M.; Zhao, Y.; et al. Molecular characterization of infectious bronchitis virus in Southwestern China for the protective efficacy evaluation of four live vaccine strains. Vaccine 2022, 40, 255–265. [Google Scholar]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef]
- Casais, R.; Dove, B.; Cavanagh, D.; Britton, P. Recombinant avian infectious bronchitis virus expressing a heterologous spike gene demonstrates that the spike protein is a determinant of cell tropism. J. Virol. 2003, 77, 9084–9089. [Google Scholar]
- Huo, J.; Zhang, G.; Wang, L.; Sun, W.; Jia, L.; Yang, X.; Liu, Y. Combining the advantages of prokaryotic expression and T7 phage display systems to obtain antigens for antibody preparation. Protein Expr. Purif. 2021, 184, 105808. [Google Scholar]
- Shirvani, E.; Paldurai, A.; Manoharan, V.K.; Varghese, B.P.; Samal, S.K. A Recombinant Newcastle Disease Virus (NDV) Expressing S Protein of Infectious Bronchitis Virus (IBV) Protects Chickens against IBV and NDV. Sci. Rep. 2018, 8, 11951. [Google Scholar]
- Keep, S.; Stevenson-Leggett, P.; Webb, I.; Fones, A.; Kirk, J.; Britton, P.; Bickerton, E. The spike protein of the apathogenic Beaudette strain of avian coronavirus can elicit a protective immune response against a virulent M41 challenge. PLoS ONE 2024, 19, e0297516. [Google Scholar]
- Wang, L.; Parr, R.L.; King, D.J.; Collisson, E.W. A highly conserved epitope on the spike protein of infectious bronchitis virus. Arch. Virol. 1995, 140, 2201–2213. [Google Scholar]
- Kant, A.; Koch, G.; van Roozelaar, D.J.; Kusters, J.G.; Poelwijk, F.A.; van der Zeijst, B.A. Location of antigenic sites defined by neutralizing monoclonal antibodies on the S1 avian infectious bronchitis virus glycopolypeptide. J. Gen. Virol. 1992, 73, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Parr, R.L.; Collissor, E.W. Epitopes on the spike protein of a nephropathogenic strain of infectious bronchitis virus. Arch. Virol. 1993, 133, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Bande, F.; Arshad, S.S.; Hair Bejo, M.; Kadkhodaei, S.; Omar, A.R. Prediction and In Silico Identification of Novel B-Cells and T-Cells Epitopes in the S1-Spike Glycoprotein of M41 and CR88 (793/B) Infectious Bronchitis Virus Serotypes for Application in Peptide Vaccines. Adv. Bioinform. 2016, 2016, 5484972. [Google Scholar] [CrossRef] [PubMed]
- Ignjatovic, J.; Sapats, S. Identification of previously unknown antigenic epitopes on the S and N proteins of avian infectious bronchitis virus. Arch. Virol. 2005, 150, 1813–1831. [Google Scholar] [CrossRef]
- Zou, N.; Xia, J.; Wang, F.; Duan, Z.; Miao, D.; Yan, Q.; Cao, S.; Wen, X.; Liu, P.; Huang, Y. Two novel neutralizing antigenic epitopes of the s1 subunit protein of a QX-like avian infectious bronchitis virus strain Sczy3 as revealed using a phage display peptide library. Vet. Immunol. Immunopathol. 2015, 168, 49–55. [Google Scholar] [CrossRef]
- Bashir, A.; Li, S.; Ye, Y.; Zheng, Q.; Knanghat, R.; Bashir, F.; Shah, N.N.; Yang, D.; Xue, M.; Wang, H.; et al. SARS-CoV-2 S protein harbors furin cleavage site located in a short loop between antiparallel β-strand. Int. J. Biol. Macromol. 2024, 281, 136020. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Name | Primer Sequence (5′–3′) | Primer Size | Product Size (bp) |
|---|---|---|---|---|
| S1 | S1-F | TCGGTACCGATGGTTGGCATTTACATGG | 28 | 660 |
| S1-R | ACAAGCTTGACAATAAACTTCTGCTTAAC | 29 | ||
| S1-38 | S1-38-F | TCGGTACCGATGGTTGGCATTTACATGG | 28 | 459 |
| S1-38-R | CTATCTAGAAAAATAAACACCTGCAGATGT | 30 | ||
| S1-114 | S1-114-F | CTGGTACCCATGTTGGGTGTCCTATAAC | 28 | 423 |
| S1-114-R | ACAAGCTTGACAATAAACTTCTGCTTAAC | 29 | ||
| S1-190 | S1-190-F | CTGGTACCCATGTTGGGTGTCCTATAAC | 28 | 231 |
| S1-190-R | CTATCTAGAAAAATAAACACCTGCAGATGT | 30 | ||
| S1-107 | S1-107-F | TCGGATCCACTACAGTGTTTGTTACACATT | 30 | 198 |
| S1-107-R | GCAAGCTTATCACCATTTAAATATACGGATGT | 32 | ||
| S1-128 | S1-128-F | TCGGATCCCAACAGCATTCTATACGTGTT | 29 | 198 |
| S1-128-R | GCAAGCTTAGCTTTAAAATAAACACCTGC | 29 |
| Component | Volume (µL) | Concentration |
|---|---|---|
| PrimeSTAR HS Premix | 25 | 2× |
| Primer-F | 1 | 10 µM |
| Primer-R | 1 | 10 µM |
| cDNA | 1 | 100 ng/µL |
| ddH2O | Add to 50 |
| Reaction Basic Steps | Temperature (°C) | Time | Cycles |
|---|---|---|---|
| Initial denaturation | 98 | 2 min | 1 |
| Denaturation | 98 | 10 s | 35 |
| Annealing | 56 | 5 s | 35 |
| Extension | 72 | 1 min | 35 |
| Final Extension | 72 | 5 min | 1 |
| mAbs | Antibody Potency of Supernatant | Heavy Chain | Light Chain |
|---|---|---|---|
| 2A10 | 1:6400 | IgG1 | Kappa |
| 4E9 | 1:12,800 | IgG1 | Kappa |
| 5E12 | 1:1600 | IgG1 | Kappa |
| Vector Truncation | mAbs | ||
|---|---|---|---|
| 2A10 | 4E9 | 5E12 | |
| S1-38 | + | + | + |
| S1-114 | + | + | + |
| S1-190 | + | + | + |
| S1-107 | + | + | + |
| S1-128 | + | + | + |
| Peptide S1-128 | + | - | - |
| Peptide S1-132 | + | - | - |
| Peptide S1-137 | - | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Z.; Hu, J.; Cai, Y.; Liu, Y.; Yin, G.; Guo, X.; Wang, R.; Zhong, M.; Liu, Q.; Feng, X. Identification of B-Cell Epitopes Located on the Surface of the S1 Protein of Infectious Bronchitis Virus M41 Strains. Viruses 2025, 17, 464. https://doi.org/10.3390/v17040464
Gao Z, Hu J, Cai Y, Liu Y, Yin G, Guo X, Wang R, Zhong M, Liu Q, Feng X. Identification of B-Cell Epitopes Located on the Surface of the S1 Protein of Infectious Bronchitis Virus M41 Strains. Viruses. 2025; 17(4):464. https://doi.org/10.3390/v17040464
Chicago/Turabian StyleGao, Zichen, Jianing Hu, Yiqin Cai, Ye Liu, Guihu Yin, Xinyu Guo, Ruiying Wang, Meng Zhong, Qingtao Liu, and Xiuli Feng. 2025. "Identification of B-Cell Epitopes Located on the Surface of the S1 Protein of Infectious Bronchitis Virus M41 Strains" Viruses 17, no. 4: 464. https://doi.org/10.3390/v17040464
APA StyleGao, Z., Hu, J., Cai, Y., Liu, Y., Yin, G., Guo, X., Wang, R., Zhong, M., Liu, Q., & Feng, X. (2025). Identification of B-Cell Epitopes Located on the Surface of the S1 Protein of Infectious Bronchitis Virus M41 Strains. Viruses, 17(4), 464. https://doi.org/10.3390/v17040464