



Detection and In Vivo Validation of Dichorhavirus e-Probes in Meta-Transcriptomic Data via Microbe Finder (MiFi®) Discovers a Novel Host and a Possible New Strain of Orchid Fleck Virus
 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Phylogenetic Analysis of Dichorhaviruses Using Available Genome Sequences in GenBank
2.2. Buildup of the Dichorhavirus e-Probe Sequences Infecting Different Hosts
2.3. Development, Curation, and in Silico Validation of Dichorhavirus e-Probes
2.4. Detection of Dichorhaviruses Using Real-Time RT-qPCR Assays
2.5. High Throughput Sequencing of Dichorhaviruses from Different Hosts
2.6. Dichorhavirus Detection Using MiFi® Platforms and Determination of the Diagnostic Sensitivity and Specificity of Its e-Probes
3. Results
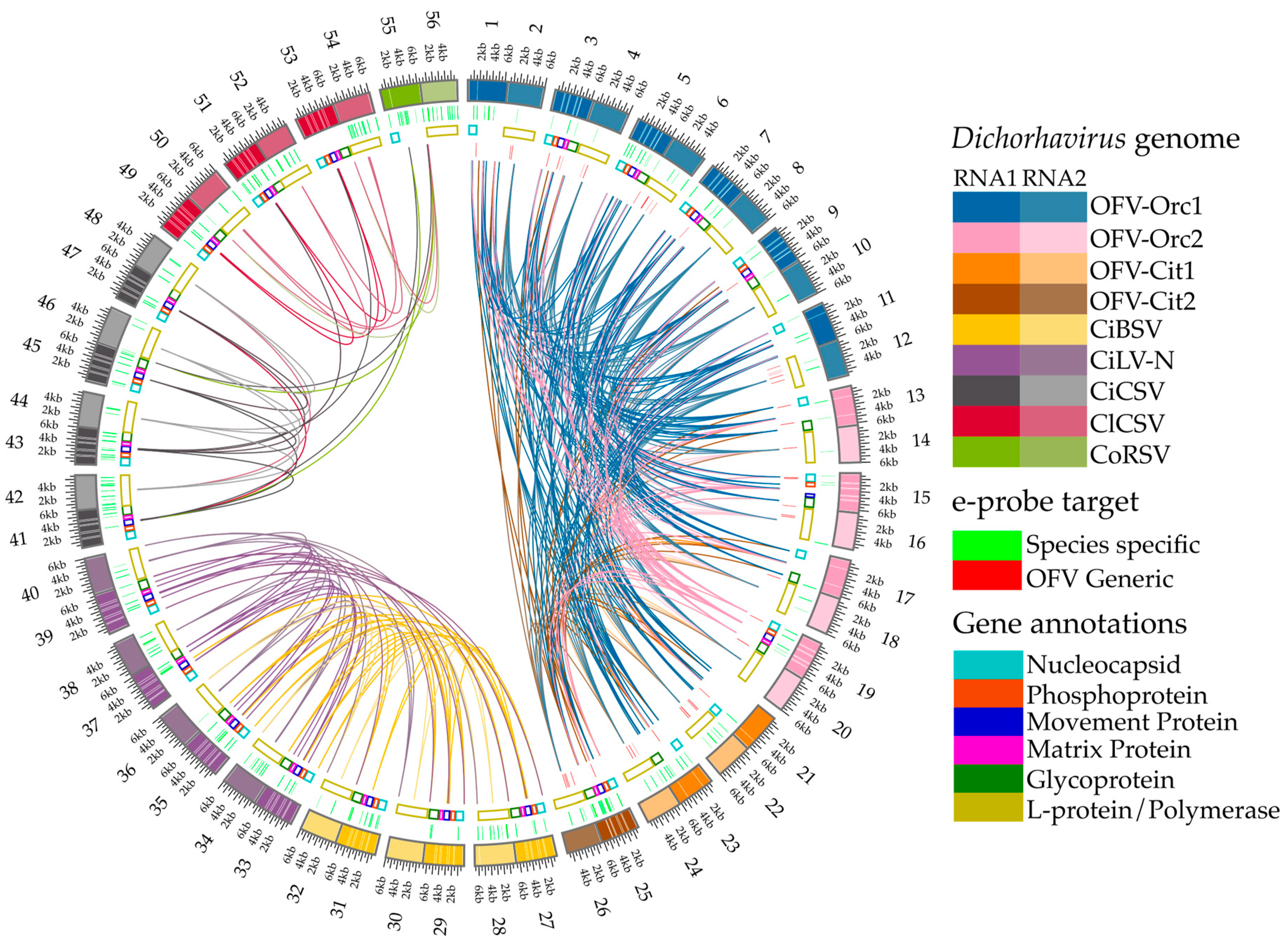
3.1. e-Probe Development and Curation
3.2. Screening of Potential Dichorhavirus-Infected Samples Using RT-qPCR Assays
3.3. Detection of Dichorhavirus Genus, Species, OFV Strains, and Variants Utilizing e-Probes in HTS Data
3.4. Diagnostic Sensitivity and Specificity for Curated Dichorhavirus e-Probes in RNA-Seq Data and Confirmation of e-Probe Diagnostics Using Bioinformatic Pipeline Analysis
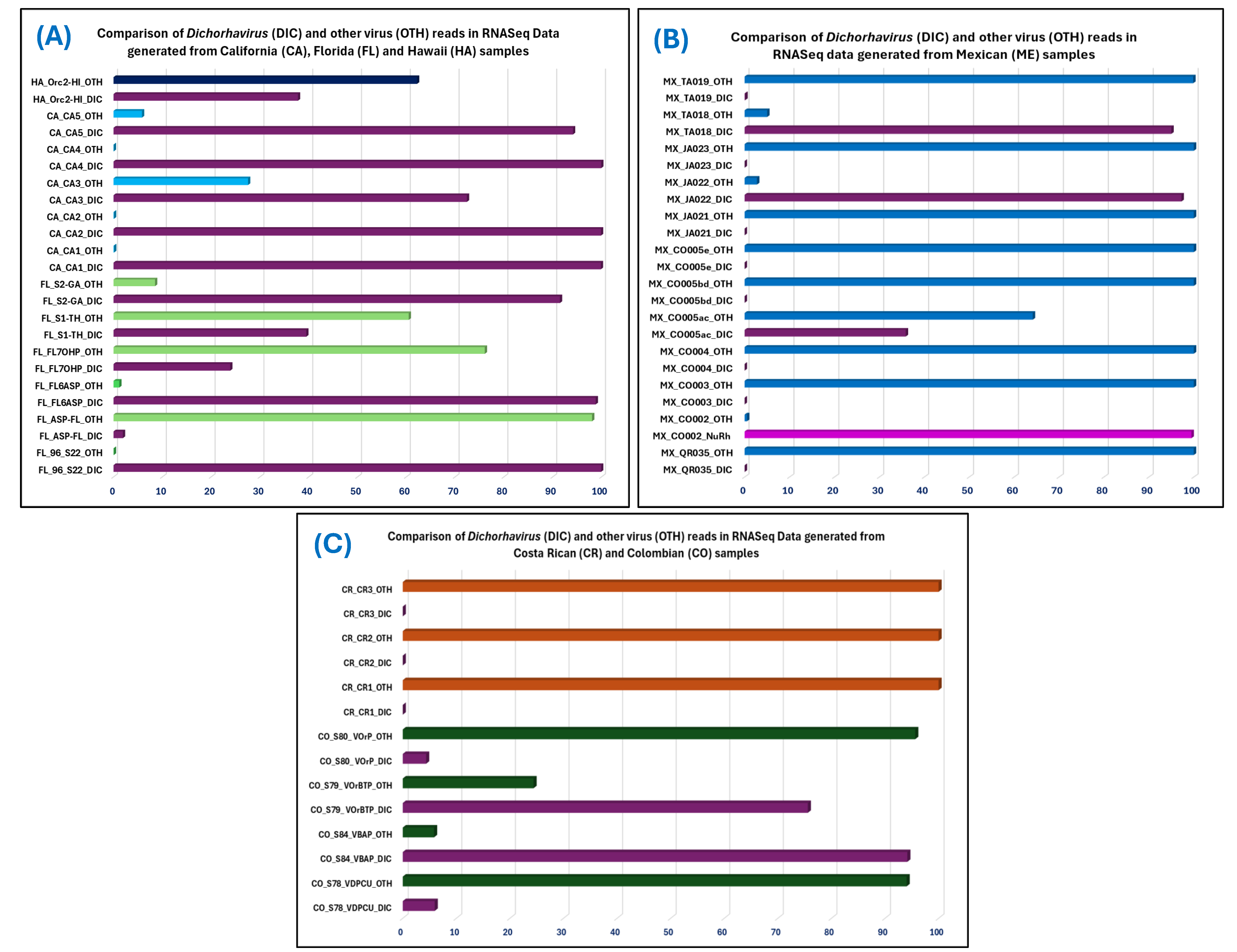
3.5. Discovery of a New Host Species of OFV and a Possible New OFV Strain in Known Hosts Utilizing e-Probe Diagnosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dietzgen, R.G.; Freitas-Astúa, J.; Chabi-Jesus, C.; Ramos-González, P.L.; Goodin, M.M.; Kondo, H.; Tassi, A.D.; Kitajima, E.W. Dichorhaviruses in Their Host Plants and Mite Vectors. In Advances in Virus Research; Academic Press Inc.: Cambridge, MA, USA, 2018; Volume 102, pp. 119–148. ISBN 9780128151945. [Google Scholar]
- Walker, P.J.; Freitas-Astua, J.; Bejerman, N.; Blasdell, K.R.; Breyta, R.; Dietzgen, R.G.; Fooks, A.R.; Kondo, H.; Kurath, G.; Kuzmin, I.V.; et al. ICTV Virus Taxonomy Profile: Rhabdoviridae 2022. J. Gen. Virol. 2022, 103, 001689. [Google Scholar] [CrossRef] [PubMed]
- Bejerman, N.; Dietzgen, R.; Debat, H. Novel Tri-Segmented Rhabdoviruses: A Data Mining Expedition Unveils the Cryptic Diversity of Cytorhabdoviruses. Viruses 2023, 15, 2402. [Google Scholar] [CrossRef] [PubMed]
- Nemchinov, L.G.; Irish, B.M.; Grinstead, S. First report of Medicago trirhavirus 1 infecting alfalfa in Washington State, USA. Plant Dis. 2024. Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Dietzgen, R.G.; Kuhn, J.H.; Clawson, A.N.; Freitas-Astúa, J.; Goodin, M.M.; Kitajima, E.W.; Kondo, H.; Wetzel, T.; Whitfield, A.E. Dichorhavirus: A Proposed New Genus for Brevipalpus Mite-Transmitted, Nuclear, Bacilliform, Bipartite, Negative-Strand RNA Plant Viruses. Arch. Virol. 2014, 159, 607–619. [Google Scholar] [CrossRef]
- Kondo, H.; Fujita, M.; Telengech, P.; Maruyam, K.; Hyodo, K.; Tassi, A.D.; Ochoa, R.; Andika, I.B.; Suzuki, N. Evidence for the replication of a plant rhabdovirus in its arthropod mite vector. Virus Res. 2025, 351, 199522. [Google Scholar] [CrossRef] [PubMed]
- de Lillo, E.; Freitas-Astúa, J.; Kitajima, E.W.; Ramos-González, P.L.; Simoni, S.; Tassi, A.D.; Valenzano, D. Phytophagous Mites Transmitting Plant Viruses: Update and Perspectives. Entomol. Gen. 2021, 41, 439–462. [Google Scholar] [CrossRef]
- Kondo, H.; Maeda, T.; Tamada, T. Orchid Fleck Virus: Brevipalpus californicus Mite Transmission, Biological Properties and Genome Structure. Exp. Appl. Acarol. 2003, 30, 215–223. [Google Scholar] [CrossRef]
- Roy, A.; Hartung, J.S.; Schneider, W.L.; Shao, J.; Leon, M.G.; Melzer, M.J.; Beard, J.J.; Otero-Colina, G.; Bauchan, G.R.; Ochoa, R.; et al. Role Bending: Complex Relationships between Viruses, Hosts, and Vectors Related to Citrus Leprosis, an Emerging Disease. Phytopathology 2015, 105, 872–884. [Google Scholar] [CrossRef]
- Mei, Y.; Bejerman, N.; Crew, K.S.; McCaffrey, N.; Dietzgen, R.G. First report of orchid fleck virus in Lilyturf (Liriope spicata) in Australia. Plant Dis. 2016, 100, 1028. [Google Scholar] [CrossRef]
- Harju, V.; Fowkes, A.R.; Skelton, A.; Adams, I.P.; Mcgreig, S.; Forde, S.M.D.; Pufal, H.; Conyers, C.; Frew, L.; Fox, A. First detection of orchid fleck virus on Veronica spicata and Dendrochilum magnum. New Dis. Rep. 2024, 50, e12312. [Google Scholar] [CrossRef]
- Olmedo Velarde, A.; Roy, A.; Padmanabhan, C.; Nunziata, S.; Nakhla, M.K.; Melzer, M.J. First Report of Orchid Fleck Virus Associated with Citrus Leprosis Symptoms in Rough Lemon (Citrus jambhiri) and Mandarin (C. reticulata) the United States. Plant Dis. 2021, 105, 2258. [Google Scholar] [CrossRef] [PubMed]
- Dey, K.; Velez-Climent, M.; Padmanabhan, C.; Nunziata, S.; Rivera, Y.; McVay, J.; Roy, A. Smilax auriculata: A new host for Orchid fleck dichorhavirus identified in Florida, USA. Plant Dis. 2021, 106, 2271. [Google Scholar] [CrossRef] [PubMed]
- Fife, A.; Carrillo, D.; Knox, G.; Iriarte, F.; Dey, K.; Roy, A.; Ochoa, R.; Bauchan, G.; Paret, M.; Martini, X. Brevipalpus-transmitted Orchid Fleck Virus infecting three new ornamental hosts in Florida. J. Integr. Pest Manag. 2021, 12, 43. [Google Scholar] [CrossRef]
- Alvarez-Quinto, R.A.; Grinstead, S.; Rott, P.; Mollov, D. Genome characterization and complete sequence of a new badnavirus from Pandanus amaryllifolius. Arch Virol. 2022, 167, 1717–1720. [Google Scholar] [CrossRef]
- Roy, A.; Espindola, A.; Shao JNunziata, S.; Rivera, Y.; Mavrodieva, V.A.; Nakhla, M.K.; Cardwell, K.F. Evaluation of electronic probes for In-silico detection of the Dichorhavirus in High-Throughput Sequencing data using Microbe Finder (MiFi®). Phytopathology 2023, S3, 149. [Google Scholar]
- Cook, G.; Kirkman, W.; Clase, R.; Steyn, C.; Basson, E.; Fourie, P.H.; Moore, S.D.; Carstens, T.G.E.; Hattingh, V. Orchid Fleck Virus Associated with the First Case of Citrus Leprosis-N in South Africa. Plant Pathol. 2019, 155, 1373–1379. [Google Scholar] [CrossRef]
- Cruz-Jaramillo, J.L.; Ruiz-Medrano, R.; Rojas-Morales, L.; Lopez-Buenfil, J.A.; Morales- Galvan, O.; Chavarin-Palacio, C.; Ramírez-Pool, J.A.; Xoconostle-Cázares, B. Characterization of a proposed Dichorhavirus associated with the citrus leprosis disease and analysis of the host response. Viruses 2014, 6, 2602–2622. [Google Scholar] [CrossRef]
- Roy, A.; Leon, M.G.; Stone, A.L.; Schneider, W.L.; Hartung, J.S.; Brlansky, R.H. First report of citrus leprosis virus nuclear type in sweet orange in Colombia. Plant Dis. 2014, 98, 1162. [Google Scholar] [CrossRef]
- Roy, A.; Stone, A.L.; Shao, J.; Otero-Colina, G.; Wei, G.; Choudhary, N.; Achor, D.; Levy, L.; Nakhla, M.K.; Hartung, J.S.; et al. Identification and molecular characterization of nuclear citrus leprosis virus, a member of the proposed Dichorhavirus genus infecting multiple Citrus species in Mexico. Phytopathology. 2015, 105, 564–575. [Google Scholar] [CrossRef]
- Roy, A.; Stone, A.L.; Otero-Colina, G.; Wei, G.; Brlansky, R.H.; Ochoa, R. Reassortment of genome segments creates stable lineages among strains of orchid fleck virus infecting citrus in Mexico. Phytopathology 2020, 110, 106–120. [Google Scholar] [CrossRef]
- Chabi-Jesus, C.; Ramos-González, P.L.; Tassi, A.D.; Guerra-Peraza, O.; Kitajima, E.W.; Harakava, R.; Beserra, J.E.A.; Salaroli, R.B.; Freitas-Astúa, J. Identification and Characterization of Citrus Chlorotic Spot Virus, a New Dichorhavirus Associated with Citrus Leprosis-like Symptoms. Plant Dis. 2018, 102, 1588–1598. [Google Scholar] [CrossRef] [PubMed]
- Chabi-Jesus, C.; Ramos-González, P.L.; Tassi, A.D.; Rossetto Pereira, L.; Bastianel, M.; Lau, D.; Canale, M.C.; Harakava, R.; Novelli, V.M.; Kitajima, E.W.; et al. Citrus Bright Spot Virus: A New Dichorhavirus, Transmitted by Brevipalpus azores, Causing Citrus Leprosis Disease in Brazil. Plants 2023, 12, 1371. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, E.W.; Chagas, C.M.; Braghini, M.T.; Fazuoli, L.C.; Locali-Fabris, E.C.; Salaroli, R.B. Natural Infection of Several Coffea Species and Hybrids and Psilanthus ebracteolatus by the Coffee Ringspot Virus (CoRSV). Sci. Agric. 2011, 68, 503–507. [Google Scholar] [CrossRef]
- Kubo, K.S.; Freitas-Astúa, J.; Machado, M.A.; Kitajima, E.W. Orchid Fleck Symptoms May Be Caused Naturally by Two Different Viruses Transmitted by Brevipalpus. J. Gen. Plant Pathol. 2009, 75, 250–255. [Google Scholar] [CrossRef]
- Kubo, K.S.; Novelli, V.M.; Bastianel, M.; Locali-Fabris, E.C.; Antonioli-Luizon, R.; Machado, M.A.; Freitas-Astúa, J. Detection of Brevipalpus-Transmitted Viruses in Their Mite Vectors by RT–PCR. Exp. Appl. Acarol. 2011, 54, 33–39. [Google Scholar] [CrossRef]
- Ramos-González, P.L.; Chabi-Jesus, C.; Guerra-Peraza, O.; Tassi, A.D.; Kitajima, E.W.; Harakava, R.; Salaroli, R.B.; Freitas-Astúa, J. Citrus Leprosis Virus N: A New Dichorhavirus Causing Citrus Leprosis Disease. Phytopathology 2017, 107, 963–976. [Google Scholar] [CrossRef]
- Stobbe, A.H.; Daniels, J.; Espindola, A.S.; Verma, R.; Melcher, U.; Ochoa-Corona, F.; Garzon, C.; Fletcher, J.; Schneider, W. E-probe Diagnostic Nucleic acid Analysis (EDNA): A theoretical approach for handling of next generation sequencing data for diagnostics. J. Microbiol. Methods 2013, 94, 356–366. [Google Scholar] [CrossRef]
- Espindola, A.S.; Cardwell, K.F. Microbe Finder (MiFi®): Implementation of an Interactive Pathogen Detection Tool in Metagenomic Sequence Data. Plants 2021, 10, 250. [Google Scholar] [CrossRef]
- Stobbe, A.H.; Schneider, W.L.; Hoyt, P.R.; Melcher, U. Screening metagenomic data for viruses using the e-probe diagnostic nucleic acid assay. Phytopathology 2014, 104, 1125–1129. [Google Scholar] [CrossRef]
- Dang, T.; Wang, H.; Espíndola, A.S.; Habiger, J.; Vidalakis, G.; Cardwell, K. Development and Statistical Validation of e-Probe Diagnostic Nucleic Acid Analysis (EDNA) Detection Assays for the Detection of Citrus Pathogens from Raw High Throughput Sequencing Data. PhytoFrontiers 2023, 3, 113–123. [Google Scholar] [CrossRef]
- Pasha, A.; Espindola, A.S.; Ziebell, H.; Ochoa-Corona, F.M. Highly Curated and Reliable E-Probes for Detection of Viral Pathogens in Unassembled HTS Datasets from Hops. PhytoFrontiers 2025. [CrossRef]
- Proaño-Cuenca, F.; Espindola, A.S.; Garzon, C.D. Detection of Phytophthora, Pythium, Globisporangium, Hyaloperonospora and Plasmopara species in High-Throughput Sequencing data by in silico and in vitro analysis using Microbe Finder (MiFi®). PhytoFrontiers 2023, 3, 124–136. [Google Scholar] [CrossRef]
- Espindola, A.; Schneider, W.; Hoyt, P.R.; Marek, S.M.; Garzon, C. A new approach for detecting fungal and oomycete plant pathogens in next generation sequencing metagenome data utilizing electronic probes. Int. J. Data Min. Bioinform 2015, 12, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Bocsanczy, A.M.; Espindola, A.S.; Cardwell, K.; Norman, D.J. Development and Validation of E-Probes with the MiFi System for Detection of Ralstonia solanacearum Species Complex in Blueberries. PhytoFrontiersTM 2023, 3, 137–147. [Google Scholar] [CrossRef]
- Narayanan, S.; Espindola, A.S.; Malayer, J.; Cardwell, K.; Ramachandran, A. Development and evaluation of Microbe Finder (MiFi)®: A novel in silico diagnostic platform for pathogen detection from metagenomic data. J. Med. Microbiol. 2023, 72, 001720. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Espindola, A.S.; Cardwell, K.; Martin, F.N.; Hoyt, P.R.; Marek, S.M.; Schneider, W.; Garzon, C.D. A step towards validation of high-throughput sequencing for the identification of plant pathogenic oomycetes. Phytopathology 2022, 112, 1859–1866. [Google Scholar] [CrossRef]
- Marçais, G.; Delcher, A.L.; Phillippy, A.M.; Coston, R.; Salzberg, S.L.; Zimin, A. MUMmer4: A fast and versatile genome alignment system. PLoS Comput Biol. 2018, 14, e1005944. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, C.; Rivera, Y.; Mavrodieva, V.A.; Roy, A. Development and Validation of TWO quadruplex TaqMan Real Time RT-PCR Assays for the Detection of the Citrus Leprosis Virus Disease Complex. Plant Health 2024, Organized by APS on July 27–30 at Memphis, Tennessee. 2024. Available online: https://www.apsnet.org/meetings/annual/meetingarchives/PH2024/Program/Pages/MemberPlayer.aspx?vid=1011140596 (accessed on 18 February 2025).
- Roy, A.; Padmanabhan, C.; Otero-Colina, G.; Rivera, Y.; Mavrodieva, V.A.; Nakhla, M.K. A multiplex real-time PCR assay for the universal detection of orchid fleck virus and differentiation among its four strains infecting multiple hosts (Abs). In Proceedings of the 12th International Conference of Plant Pathology (ICPP), Lyon, France, 2–25 August 2023; Abstract book; p. 230. [Google Scholar]
- Roy, A.; Grinstead, S.; Leon Martínez, G.; Pinzón, J.C.C.; Nunziata, S.O.; Padmanabhan, C.; Hammond, J. Meta-Transcriptomic Analysis Uncovers the Presence of Four Novel Viruses and Multiple Known Virus Genera in a Single Hibiscus rosa-sinensis Plant in Colombia. Viruses 2024, 16, 267. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, C.; Nunziata, S.; Leon, M.G.; Rivera, Y.; Mavrodieva, V.A.; Nakhla, M.K.; Roy, A. High-throughput sequencing application in the detection and discovery of viruses associated with the regulated citrus leprosis disease complex. Front. Plant Sci. 2023, 13, 1058847. [Google Scholar] [CrossRef]
- Fawcett, H.S. Scaly bark or nail-head rust of citrus. Fla. Agric. Exp. Stn. Bull. 1911, 106, 1–41. [Google Scholar]
- Hartung, J.S.; Roy, A.; Fu, S.; Shao, J.; Schneider, W.L.; Brlansky, R.H. History and diversity of citrus leprosis virus recorded in herbarium specimens. Phytopathology 2015, 105, 1277–1284. [Google Scholar] [CrossRef]
- Amarasinghe, G.K.; Bao, Y.; Basler, C.F.; Bavari, S.; Beer, M.; Bejerman, N.; Blasdell, K.R. Taxonomy of the order Mononegavirales: Update 2017. Arch. Virol. 2017, 162, 2493–2504. [Google Scholar] [CrossRef]
- Adams, I.P.; Glover, R.H.; Monger, W.A.; Mumford, R.; Jackeviciene, E.; Navalinskiene, M.; Samuitiene, M.; Boonham, N. Nextgeneration sequencing and metagenomic analysis: A universal diagnostic tool in plant virology. Mol. Plant Pathol. 2009, 10, 537–545. [Google Scholar] [CrossRef]
- Shendure, J.; Ji, H. Next-generation DNA sequencing. Nat. Biotechnol. 2008, 26, 1135–1145. [Google Scholar] [CrossRef]
- Espindola, A.S. Eukaryotic Plant Pathogen Detection Through High Throughput DNA/RNA Sequencing Data Analysis. Master’s Thesis, Oklahoma State University, Stillwater, OK, USA, 2016. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Group Based on Phylogenetic Relationship | Virus Genus/ Species/Strain /Variant | Number of Available Sequence in GenBank | e-Probes | |||||
|---|---|---|---|---|---|---|---|---|
| Length (nt) | Genome Segment | Sub-Total | Total | Grand Total | ||||
| RNA1 | RNA2 | |||||||
| Dichorhavirus | 28 | 40 | 35 | 32 | 67 | 180 | 684 | |
| 60 | 34 | 24 | 58 | |||||
| 80 | 34 | 22 | 56 | |||||
| Subgroup 1 | Orchid fleck virus (OFV) Generic | 13 | 40 | 6 | 22 | 28 | 99 | |
| 60 | 14 | 20 | 34 | |||||
| 80 | 16 | 21 | 37 | |||||
| Orchid fleck virus citrus strain (OFV-Cit) | 3 | 40 | 1 | 6 | 7 | 7 | ||
| 60 | 0 | 0 | 0 | |||||
| 80 | 0 | 0 | 0 | |||||
| Orchid fleck virus citrus strain, variant 1 (OFV-Cit1) | 2 | 40 | 4 | 0 | 4 | 14 | ||
| 60 | 5 | 0 | 5 | |||||
| 80 | 5 | 0 | 5 | |||||
| Orchid fleck virus citrus strain, variant 2 (OFV-Cit2) | 1 | 40 | 8 | 0 | 8 | 24 | ||
| 60 | 10 | 0 | 10 | |||||
| 80 | 6 | 0 | 6 | |||||
| Orchid fleck virus orchid strain (OFV-Orc) | 10 | 40 | 1 | 2 | 3 | 3 | ||
| 60 | 0 | 0 | 0 | |||||
| 80 | 0 | 0 | 0 | |||||
| Orchid fleck virus orchid strain, variant 1 (OFV-Orc1) | 6 | 40 | 22 | 0 | 22 | 56 | ||
| 60 | 19 | 0 | 19 | |||||
| 80 | 15 | 0 | 15 | |||||
| Orchid fleck virus orchid strain, variant 2 (OFV-Orc2) | 4 | 40 | 5 | 0 | 5 | 6 | ||
| 60 | 1 | 0 | 1 | |||||
| 80 | 0 | 0 | 0 | |||||
| Subgroup 2 | Citrus bright spot virus (CiBSV) | 3 | 40 | 13 | 8 | 21 | 36 | |
| 60 | 7 | 2 | 9 | |||||
| 80 | 5 | 1 | 6 | |||||
| Citrus leprosis virus N (CiLV-N) | 4 | 40 | 15 | 16 | 31 | 78 | ||
| 60 | 16 | 9 | 25 | |||||
| 80 | 13 | 9 | 22 | |||||
| Subgroup 3 | Citrus chlorotic spot virus (CiCSV) | 4 | 40 | 16 | 13 | 29 | 73 | |
| 60 | 11 | 15 | 26 | |||||
| 80 | 8 | 10 | 18 | |||||
| Clerodendrum chlorotic spot virus (ClCSV) | 3 | 40 | 10 | 13 | 23 | 65 | ||
| 60 | 9 | 13 | 22 | |||||
| 80 | 8 | 12 | 20 | |||||
| Coffee ring spot virus (CoRSV) | 1 | 40 | 10 | 8 | 18 | 43 | ||
| 60 | 7 | 8 | 15 | |||||
| 80 | 4 | 6 | 10 | |||||
| Name | RT-qPCR Diagnostic Based on CT Value | Comments Based on CT Value Obtained by RT-qPCR | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genus Specific | Species Specific | OFV Strain Specific | ||||||||||||||||
| Sl. No. | Host | Isolate | City, State | Country | Dichorhavirus * | CiBSV * | CiCSV | CiLV-N | ClCSV | CoRSV * | OFV-Gen | OFV-Cit * | OFV-Cit1 | OFV-Cit2 | OFV-Orc * | OFV-Orc1 | OFV-Orc2 | |
| 1 | Smilax auriculata | 96_S22 | Gainesville, Florida | USA | NT | NT | UD | UD | UD | NT | 13.92 | NT | UD | UD | NT | 27.74 | 15.66 | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 |
| 2 | Liriope sp. | S2-GA | NT | NT | UD | UD | UD | NT | 14.35 | NT | UD | UD | NT | UD | 18.52 | Positive for OFV-Gen, & OFV-Orc2 | ||
| 3 | S1-TH | Tallahassee, Florida | NT | NT | UD | UD | UD | NT | 13.87 | NT | UD | UD | NT | UD | UD | Positive for OFV-Gen | ||
| 4 | Aspidistra sp. | ASP-FL | NT | NT | UD | UD | UD | NT | 23.47 | NT | UD | UD | NT | UD | 33.26 | Positive for OFV-Gen, & OFV-Orc2 | ||
| 5 | FL6ASP | NT | NT | UD | UD | UD | NT | 17.42 | NT | UD | UD | NT | 31.56 | 27.71 | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 | |||
| 6 | Ophiopogan sp. | FL7OHP | NT | NT | UD | UD | UD | NT | 26.39 | NT | UD | UD | NT | 31.37 | 29.12 | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 | ||
| 7 | Cymbidium sp. | CA1 | San Diego, California | NT | NT | UD | UD | UD | NT | 18.05 | NT | UD | UD | NT | UD | 17.08 | Positive for OFV-Gen, & OFV-Orc2 | |
| 8 | CA4 | NT | NT | UD | UD | UD | NT | 18.38 | NT | UD | UD | NT | UD | 18.97 | Positive for OFV-Gen, & OFV-Orc2 | |||
| 9 | Dendrobium sp. | CA2 | NT | NT | UD | UD | UD | NT | 14.03 | NT | UD | UD | NT | 15.91 | 18.66 | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 | ||
| 10 | CA3 | NT | NT | UD | UD | UD | NT | 22.26 | NT | UD | UD | NT | 26.05 | 22.99 | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 | |||
| 11 | Dendrochilum magnum | CA5 | NT | NT | UD | UD | UD | NT | 14.48 | NT | UD | UD | NT | 30.44 | 14.56 | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 | ||
| 12 | Citrus reticulata | Orc2-HI | Hilo, Hawaii | NT | NT | UD | UD | UD | NT | 23.67 | NT | UD | UD | NT | UD | 25.94 | Positive for OFV-Gen, & OFV-Orc2 | |
| 13 | Farfugium japonicum | QR035 | El Pueblito, Querétaro | Mexico | NT | NT | UD | UD | UD | NT | 24.32 | NT | 27.92 | UD | NT | UD | UD | Positive for OFV-Gen, & OFV-Cit1 |
| 14 | Aralia sp. | CO002 | Manzanillo, Colima, | NT | NT | UD | UD | UD | NT | 32.19 | NT | UD | UD | NT | UD | UD | Positive for OFV-Gen | |
| 15 | Hibiscus rosa-sinensis | CO003 | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | ||
| 16 | CO004 | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | |||
| 17 | Dendrobium sp. | CO005ac | NT | NT | UD | UD | UD | NT | 30.52 | NT | UD | UD | NT | UD | UD | Positive for OFV-Gen | ||
| 18 | CO005bd | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | |||
| 19 | CO005e | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | |||
| 20 | Citrus aurantium | JA021 | Sayula, Jalisco | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | |
| 21 | JA022 | NT | NT | UD | UD | UD | NT | 13.44 | NT | UD | 17.99 | NT | UD | UD | Positive for OFV-Gen, & OFV-Cit2 | |||
| 22 | JA023 | Zapotlanejo, Jalisco | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | ||
| 23 | Hibiscus rosa-sinensis | TA018 | Villahermosa, Tabasco | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | |
| 24 | Pedilanthus tithymaloides | TA019 | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | ||
| 25 | Orchids | S78_VDPCU | Pradera, Valle del Cauca | Colombia | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species |
| 26 | S79_ VOrBTP | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | |||
| 27 | S80_VOrP | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | |||
| 28 | S84_VBAP | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | |||
| 29 | Liriope sp. | CR1 | San José | Costa Rica | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species |
| 30 | Sapium sp. | CR2 | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | ||
| 31 | CR3 | NT | NT | UD | UD | UD | NT | UD | NT | UD | UD | NT | UD | UD | Negative for tested dichorhavirus species | |||
| RNASeq Data Generated from Different Host (Isolate) | Genus, Species Strain and Variant Specific e-Probes | Pairwise t Test Diagnostic Based on Type of Libraries and Legth of e-Probes | EDNA Diagnotic Results Based on Combined Analysis of 40, 60 and 80 NTs e-Probes | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Single Library | Concatenated Libraries | |||||||||
| p Value | Diagnostic | p Value | Diagnostic | |||||||
| 40 nt | 60 nt | 80 nt | 40 nt | 60 nt | 80 nt | |||||
| Smilax auriculata (96_S22) | Dichorhavirus | 1.95 × 10−5 | 0.0002 | 0.0025 | Positive | 1.58 × 10−5 | 0.0001 | 0.0005 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc, OFV-Orc1 & OFV-Orc2, suspicious for OFV-Cit |
| OFV-Gen | 6.93 × 10−12 | 6.93 × 10−10 | 3.52 × 10−6 | Positive | 5.32 × 10−12 | 7.11 × 10−10 | 2.53 × 10−8 | Positive | ||
| OFV-Cit | 0.1029 | NP | NP | Negative | 0.0735 | NP | NP | Suspicious | ||
| OFV-Cit1 | 0.2119 | NC | NC | Negative | 0.2119 | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | 2.32 × 10−5 | NP | NP | Positive | 3.46 × 10−5 | NP | NP | Positive | ||
| OFV-Orc1 | 0.0411 | 0.0209 | NC | Positive | 0.0023 | 0.0004 | 0.1753 | Positive | ||
| OFV-Orc2 | 1.71 × 10−7 | 0.5 | NP | Positive | 1.95 × 10−7 | 0.5 | NP | Positive | ||
| Aspidistra sp. (Asp-FL) | Dichorhavirus | 0.0209 | 0.0596 | 0.1135 | Positive | 0.0006 | 0.0099 | 0.0591 | Positive | True Positive for Dichorhavirus, OFV-Gen, & OFV-Orc suspicious for OFV-Orc1 |
| OFV-Gen | 0.0002 | 0.0007 | 0.0031 | Positive | 4.37 × 10−5 | 0.0004 | 0.0003 | Positive | ||
| OFV-Cit | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | 0.0921 | NP | NP | Negative | 0.0380 | NP | NP | Positive | ||
| OFV-Orc1 | 0.0811 | NC | NC | Suspicious | 0.0946 | NC | NC | Suspicious | ||
| OFV-Orc2 | NC | NC | NP | Negative | NC | NC | NP | Negative | ||
| Aspidistra sp. (FL6ASP) | Dichorhavirus | 0.0006 | 0.0103 | 0.0287 | Positive | 6.64 × 10−5 | 0.0022 | 0.0131 | Positive | True Positive for Dichorhavirus, OFV-Gen, & OFV-Orc, suspicious for OFV-Cit, OFV-Orc1 and OFV-Cit2 |
| OFV-Gen | 6.48 × 10−8 | 5.71 × 10−6 | 5.94 × 10−5 | Positive | 3.43 × 10−9 | 2.36 × 10−6 | 4.55 × 10−5 | Positive | ||
| OFV-Cit | 0.1957 | NP | NP | Negative | 0.0485 | NP | NP | Suspicious | ||
| OFV-Cit1 | 0.2119 | NC | NC | Negative | 0.2119 | NC | NC | Negative | ||
| OFV-Cit2 | NC | 0.1231 | 0.2023 | Negative | 0.0853 | 0.1158 | NC | Suspicious | ||
| OFV-Orc | 0.0921 | NP | NP | Suspicious | 0.0380 | NP | NP | Positive | ||
| OFV-Orc1 | 0.0811 | NC | NC | Suspicious | 0.0946 | NC | NC | Negative | ||
| OFV-Orc2 | NC | NC | NP | Negative | NC | NC | NP | Negative | ||
| Ophiopogan sp. (FL7OHP) | Dichorhavirus | 0.0049 | 0.0902 | 0.1094 | Positive | 0.0011 | 0.0589 | 0.0873 | Positive | True Positive for Dichorhavirus, OFV-Gen, & OFV-Orc1, suspicious for OFV-Orc |
| OFV-Gen | 7.85 × 10−5 | 0.0008 | 0.0115 | Positive | 6.60 × 10−7 | 6.88 × 10−5 | 0.0037 | Positive | ||
| OFV-Cit | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | 0.0938 | NP | NP | Suspicious | 0.0938 | NP | NP | Suspicious | ||
| OFV-Orc1 | 0.0415 | 0.1717 | NC | Positive | 0.0076 | 0.1753 | 0.0814 | Positive | ||
| OFV-Orc2 | 0.2119 | NC | NP | Negative | 0.2119 | NC | NP | Negative | ||
| Liriope sp. (S1-TH) | Dichorhavirus | 0.0010 | 0.0063 | 0.0288 | Positive | 5.30 × 10−7 | 5.04 × 10−6 | 3.88 × 10−6 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc, OFV-Orc1, & OFV-Orc2, suspicious for OFV-Cit |
| OFV-Gen | 3.33 × 10−7 | 4.54 × 10−5 | 0.0001 | Positive | 1.76 × 10−12 | 2.45× 10−8 | 9.89 × 10−9 | Positive | ||
| OFV-Cit | NC | NP | NP | Negative | 0.0593 | NP | NP | Suspicious | ||
| OFV-Cit1 | NC | NC | NC | Negative | 0.2119 | NC | NC | Negative | ||
| OFV-Cit2 | NC | 0.1299 | NC | Negative | 0.1305 | 0.2023 | 0.2023 | Negative | ||
| OFV-Orc | 0.0606 | NP | NP | Suspicious | 0.0001 | NP | NP | Positive | ||
| OFV-Orc1 | 0.0058 | 0.0266 | 0.0663 | Positive | 2.58 × 10−5 | 0.0104 | 0.0412 | Positive | ||
| OFV-Orc2 | 0.0319 | NC | NP | Positive | 3.53 × 10−8 | NC | NP | Positive | ||
| Liriope sp. (S2-GA) | Dichorhavirus | 0.0010 | 0.0032 | 0.0003 | Positive | 4.15 × 10−6 | 6.60 × 10−5 | 0.0003 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Cit, OFV-Orc & OFV-Orc2 |
| OFV-Gen | 3.09 × 10−6 | 1.95 × 10−5 | 0.0003 | Positive | 2.21 × 10−13 | 3.58 × 10−9 | 2.41 × 10−8 | Positive | ||
| OFV-Cit | 0.1957 | NP | NP | Negative | 0.0422 | NP | NP | Positive | ||
| OFV-Cit1 | NC | NC | NC | Negative | 0.2119 | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | 0.0654 | NP | NP | Suspicious | 3.49 × 10−5 | NP | NP | Positive | ||
| OFV-Orc1 | NC | NC | NC | Negative | NC | 0.1717 | NC | Negative | ||
| OFV-Orc2 | 0.0192 | NC | NP | Positive | 1.14 × 10−7 | 0.5 | NP | Positive | ||
| Cymbidium sp. (CA1) | Dichorhavirus | 3.17 × 10−5 | 0.0004 | 0.0044 | Positive | 1.98 × 10−5 | 0.0002 | 0.0028 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc, & OFV-Orc2 |
| OFV-Gen | 3.79 × 10−12 | 6.47 × 10−10 | 5.37 × 10−8 | Positive | 6.75 × 10−13 | 5.46 × 10−10 | 1.09 × 10−8 | Positive | ||
| OFV-Cit | 0.1957 | NP | NP | Negative | 0.1957 | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | 0.0001 | NP | NP | Positive | 0.0001 | NP | NP | Positive | ||
| OFV-Orc1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc2 | 4.50 × 10−8 | NC | NP | Positive | 3.40 × 10−8 | 0.5 | NP | Positive | ||
| Dendrobium sp. (CA2) | Dichorhavirus | 4.40 × 10−6 | 4.89 × 10−5 | 0.0004 | Positive | 9.79 × 10−7 | 1.17 × 10−5 | 1.04 × 10−5 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Cit, OFV-Orc, OFV-Orc1 & OFV-Orc2 and suspicious for OFV-Cit2 |
| OFV-Gen | 1.26 × 10−14 | 2.24 × 10−9 | 4.93 × 10−7 | Positive | 9.49 × 10−17 | 6.51 × 10−12 | 2.28 × 10−8 | Positive | ||
| OFV-Cit | 0.0432 | NP | NP | Positive | 0.0586 | NP | NP | Suspicious | ||
| OFV-Cit1 | NC | NC | NC | Negative | 0.2119 | NC | NC | Negative | ||
| OFV-Cit2 | NC | 0.1841 | NC | Negative | 0.1908 | 0.0854 | 0.2023 | Suspicious | ||
| OFV-Orc | 0.0317 | NP | NP | Positive | 3.55 × 10−5 | NP | NP | Positive | ||
| OFV-Orc1 | 1.51 × 10−20 | 3.06 × 10−11 | 5.41 × 10−5 | Positive | 5.52 × 10−40 | 7.85 × 10−32 | 1.58 × 10−6 | Positive | ||
| OFV-Orc2 | 2.53 × 10−3 | NC | NP | Positive | 0.0029 | NC | NP | Positive | ||
| Dendrobium sp. (CA3) | Dichorhavirus | 9.66 × 10−5 | 2.17 × 10−8 | 0.0236 | Positive | 0.0001 | 0.0012 | 0.0131 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc1 & OFV-Orc2 and suspicious for OFV-Orc |
| OFV-Gen | 5.38 × 10−8 | 2.26 × 10−7 | 6.19 × 10−5 | Positive | 7.01 × 10−8 | 8.84 × 10−8 | 1.62 × 10−5 | Positive | ||
| OFV-Cit | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | 0.0938 | NP | NP | Suspicious | 0.0820 | NP | NP | Suspicious | ||
| OFV-Orc1 | 8.11 × 10−2 | 1.72 × 10−1 | NC | Positive | 5.73 × 10−5 | 1.55 × 10−6 | 0.0096 | Positive | ||
| OFV-Orc2 | 9.73 × 10−3 | NC | NP | Positive | 0.0081 | NC | NP | Positive | ||
| Cymbidium sp. (CA4) | Dichorhavirus | 3.51 × 10−5 | 0.0005 | 0.0067 | Positive | 2.77 × 10−5 | 0.0003 | 0.0033 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc & OFV-Orc2 |
| OFV-Gen | 5.84 × 10−12 | 8.31 × 10−10 | 1.05 × 10−7 | Positive | 2.61 × 10−12 | 5.52 × 10−10 | 1.74 × 10−8 | Positive | ||
| OFV-Cit | 0.1957 | NP | NP | Negative | 0.1957 | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | 0.0008 | NP | NP | Positive | 0.0002 | NP | NP | Positive | ||
| OFV-Orc1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc2 | 3.53 × 10−8 | NC | NP | Positive | 3.44 × 10−8 | NC | NP | Positive | ||
| Dendrochilum magnum (CA5) | Dichorhavirus | 3.35 × 10−5 | 0.0004 | 0.0060 | Positive | 2.58 × 10−5 | 0.0003 | 0.0030 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc, OFV-Orc1 & OFV-Orc2 |
| OFV-Gen | 6.85 × 10−12 | 7.11 × 10−10 | 1.07 × 10−7 | Positive | 3.38 × 10−12 | 5.39 × 10−10 | 2.29 × 10−8 | Positive | ||
| OFV-Cit | 0.1957 | NP | NP | Negative | 0.1957 | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | 0.0004 | NP | NP | Positive | 0.0001 | NP | NP | Positive | ||
| OFV-Orc1 | 0.1699 | NC | NC | Negative | 0.0107 | NC | NC | Positive | ||
| OFV-Orc2 | 3.48 × 10−8 | 0.5 | NP | Positive | 3.44 × 10−8 | 0.5 | NP | Positive | ||
| Citrus reticulata (Orc2-HI) | Dichorhavirus | 0.0416 | 0.1629 | 0.1630 | Positive | 1.67 × 10−5 | 0.0001 | 0.0008 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc, & OFV-Orc2 |
| OFV-Gen | 0.0217 | NC | NC | Positive | 1.96 × 10−11 | 7.14 × 10−9 | 6.37 × 10−8 | Positive | ||
| OFV-Cit | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | NC | NP | NP | Negative | 0.0001 | NP | NP | Positive | ||
| OFV-Orc1 | NC | NC | NC | Negative | NC | 0.1717 | NC | Negative | ||
| OFV-Orc2 | NC | NC | NP | Negative | 8.88 × 10−8 | 0.5 | NP | Positive | ||
| Aralia sp. (CO002) | Dichorhavirus | 0.0794 | 0.0416 | NC | Positive | 0.0794 | 0.0416 | NC | Positive | True Positive for Dichorhavirus and suspicious for OFV-Gen |
| OFV-Gen | NC | NC | NC | Negative | 0.0805 | 0.0802 | NC | Suspicious | ||
| OFV-Cit | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Orc1 | NC | NC | NC | Negative | 0.0415 | 0.0209 | NC | Positive | ||
| OFV-Orc2 | NC | NC | NP | Negative | NC | NC | NP | Negative | ||
| Dendrobium sp. (CO005ac) | Dichorhavirus | 0.0794 | NC | NC | Suspicious | 0.0794 | NC | NC | Suspicious | Suspicious for dichorhavirus as infected with Novel dichorhavirus |
| OFV-Gen | 0.1153 | NC | NC | Negative | 0.1195 | NC | NC | Negative | ||
| OFV-Cit | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Orc1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc2 | NC | NC | NP | Negative | NC | NC | NP | Negative | ||
| Farfugium japonicum (QR035) | Dichorhavirus | 0.0137 | 0.0536 | NC | Positive | 0.0013 | 0.0139 | 0.0488 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Cit, & OFV-Cit1 |
| OFV-Gen | 0.0013 | 0.0005 | 0.0173 | Positive | 3.93 × 10−6 | 1.85 × 10−5 | 7.61 × 10−5 | Positive | ||
| OFV-Cit | 0.0239 | NP | NP | Positive | 0.0046 | NP | NP | Positive | ||
| OFV-Cit1 | 0.1021 | 0.0892 | NC | Suspicious | 0.0256 | 0.0434 | 0.2119 | Positive | ||
| OFV-Cit2 | NC | NC | NC | Negative | 0.1908 | NC | NC | Negative | ||
| OFV-Orc | NC | NP | NP | Negative | 0.2525 | NP | NP | Negative | ||
| OFV-Orc1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc2 | NC | NC | NP | Negative | NC | NC | NP | Negative | ||
| Citrus aurantium (JA022) | Dichorhavirus | 0.0002 | 0.0011 | 1.89 × 10−2 | Positive | 2.23 × 10−5 | 0.0001 | 0.0038 | Positive | True Positive for Dichorhavirus, OFV-Gen, OFV-Cit, & OFV-Cit2 and Suspicious for OFV-Orc2 |
| OFV-Gen | 7.48 × 10−8 | 1.39 × 10−6 | 4.65 × 10−5 | Positive | 8.22 × 10−9 | 2.66 × 10−7 | 2.20 × 10−6 | Positive | ||
| OFV-Cit | 6.85 × 10−8 | NP | NP | Positive | 1.23 × 10−9 | NP | NP | Positive | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | 0.0001 | 7.76 × 10−6 | 0.0042 | Positive | 2.26 × 10−6 | 3.89 × 10−14 | 0.0014 | Positive | ||
| OFV-Orc | 0.2525 | NP | NP | Negative | 0.1962 | NP | NP | Negative | ||
| OFV-Orc1 | 0.1584 | 0.1717 | NC | Negative | 0.1548 | 0.1717 | NC | Negative | ||
| OFV-Orc2 | NC | NC | NP | Negative | 0.0892 | NC | NP | Suspicious | ||
| Citrus aurantium (JA023) | Dichorhavirus | NC | NC | NC | Negative | NC | NC | NC | Negative | True Negative for all the e-probes |
| OFV-Gen | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Orc1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc2 | NC | NC | NP | Negative | NC | NC | NP | Negative | ||
| Sapium sp. (CR2) | Dichorhavirus | NC | NC | NC | Negative | NC | NC | NC | Negative | True Negative for all the e-probes as infected with Emravirus |
| OFV-Gen | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Cit1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Cit2 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc | NC | NP | NP | Negative | NC | NP | NP | Negative | ||
| OFV-Orc1 | NC | NC | NC | Negative | NC | NC | NC | Negative | ||
| OFV-Orc2 | NC | NC | NP | Negative | NC | NC | NP | Negative | ||
| True Positive (TP) Detection by EDNA Diagnosis Using e-Probes of Different Length | False Negative (FN) Detection by EDNA Diagnosis in Comparison with RT-qPCR and Bioinformatic Analysis | Sensitivity (TP/TP+FN) of EDNA Diagnostics in Comparison with RT-qPCR and Bioinformatic Analysis | Validation of False Negative (FN) Detected in EDNA Diagnosis by Comparing the Results of | Sensitivity (TP/TP+FN) of EDNA Diagnosis in Comparison with | |||||||||||||||||
| 40 nt | 60 nt | 80 nt | All Sizes Together | 40 nt | 60 nt | 80 nt | 40 nt | 60 nt | 80 nt | RT-qPCR | RT-qPCR and/or Bioinformatic Analysis | RT-qPCR and Bioinformatic Analysis Plus RT-qPCR | |||||||||
| Single | Concate | Single | Concate | Single | Concate | Combined | Single | Concate | Single | Concate | Single | Concate | Single | Concate | Single | Concate | Single | Concate | |||
| 14 | 14 | 11 | 14 | 10 | 12 | 15 | 0 | 0 | 4 | 1 | 4 | 2 | 1 | 1 | 0.7895 | 0.9375 | 0.7895 | 0.8824 | ND | 1 | ND/0.9375 |
| 14 | 14 | 13 | 14 | 13 | 14 | 14 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 0.9333 | 1 | 0.9333 | 1 | 1 | 0 | 0.9333/1 |
| 3 | 3 | ND | ND | ND | ND | 3 | 0 | 0 | ND | ND | ND | ND | 1 | 1 | NT | NT | NT | NT | ND | 0 | ND/1 |
| 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0.5/1 |
| 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 0.5 | 1 | 0.5 | 1 | 0.5 | 1 | 1 | 0 | 0.5/1 |
| 5 | 10 | ND | ND | ND | ND | 10 | 5 | 2 | ND | ND | ND | ND | 0.6667 | 0.8333 | NT | NT | NT | NT | ND | 1 | ND/0.9091 |
| 5 | 6 | 4 | 5 | 1 | 4 | 6 | 1 | 0 | 1 | 0 | 1 | 0 | 0.8571 | 1 | 0.8571 | 1 | 0.8571 | 1 | 1 | 0 | 0.8571/1 |
| 8 | 9 | 0 | 0 | ND | ND | 9 | 2 | 1 | 9 | 9 | ND | ND | 0.8182 | 0.9 | 0.5 | 0.5 | NT | NT | 4 | 1 | 0.6923/0.9 |
| True Negative (TN) Detection by EDNA Diagnosis Using e-Probes of Different Length | False Positive (FP) Detection by EDNA Diagnosis in Comparison with RT-qPCR and Bioinformatic Analysis | Specificity (TN/TN+FP) of EDNA Diagnostics in Comparison with RT-qPCR and Bioinformatic Analysis | Validation of False Positive (FP) Detected in EDNA Diagnosis by Comparing the Results of | Specificity (TN/TN+FP) of EDNA Diagnosis in Comparison with | |||||||||||||||||
| 40 nt | 60 nt | 80 nt | All Sizes Together | 40 nt | 60 nt | 80 nt | 40 nt | 60 nt | 80 nt | RT-qPCR | RT-qPCR and/or Bioinformatic Analysis | RT-qPCR and Bioinformatic Analysis Plus RT-qPCR | |||||||||
| Single | Concate | Single | Concate | Single | Concate | Combined | Single | Concate | Single | Concate | Single | Concate | Single | Concate | Single | Concate | Single | Concate | |||
| 17 | 17 | 20 | 17 | 21 | 19 | 16 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0.941176 | 0.941176 | 1 | 1 | ND | 1 | ND/0.9412 |
| 17 | 17 | 18 | 17 | 18 | 17 | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | Both are 1 |
| 28 | 28 | ND | ND | ND | ND | 28 | 1 | 2 | ND | ND | ND | ND | 0.9655 | 0.9333 | NT | NT | NT | NT | ND | 1 | ND/0.9655 |
| 30 | 30 | 30 | 30 | 30 | 30 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | Both are 1 |
| 31 | 30 | 30 | 30 | 31 | 31 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | Both are 1 |
| 26 | 21 | ND | ND | ND | ND | 21 | 0 | 1 | ND | ND | ND | ND | 1 | 0.9546 | NT | NT | NT | NT | ND | 0 | ND/1 |
| 26 | 25 | 27 | 26 | 30 | 27 | 25 | 4 | 6 | 3 | 4 | 1 | 1 | 0.8621 | 0.8065 | 8929 | 0.862069 | 0.9615 | 0.9615 | 2 | 2 | Both are 0.9259 |
| 23 | 22 | 31 | 31 | ND | ND | 22 | 1 | 1 | 0 | 0 | ND | ND | 0.9565 | 0.9565 | 1 | 1 | NT | NT | 1 | 1 | Both are 0.9565 |
| Sl. No. | Isolate | Country | Meta-Transcriptomic Analysis | Sensitivity | Specificity | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Read | Total Contig Lentgh Matched with Dichorhavirus/ Nucleorhabdovirus | Total Virus Read (%) | Dichorhavirus/ Nucleorhabdovirus Among Total Virus Read (%) | Relative Abundance (RA) of Pathogen in the Sample | Detection of Dichorhavirus/Nucleorhabdovirus Related Sequences Using Bioinformatic Analysis of RNASeq Data | EDNA Diagnosis | Comments Based on CT Value Obtained by RT-qPCR | |||||||
| Raw | Post Trim | Total Virus | Dichorhavirus/Nucleorhabdovirus | Other Virus | ||||||||||
| 1 | 96_S22 | USA | 29,026,369 | 28,668,880 | 893,873 | 893,236 | 637 | 12,510 | 3.11 | 99.93 | 0.03116 | OFV Srtain-Orc2 | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc, OFV-Orc1 & OFV-Orc2, suspicious for OFV-Cit | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 |
| 2 | ASP-FL | 89,070,401 | 87,154,532 | 32,178 | 618 | 31,560 | 2040 | 0.04 | 1.92 | 7.09 × 10−6 | New OFV Srtain OFV-Orc3 | True Positive for Dichorhavirus, OFV-Gen, & OFV-Orc suspicious for OFV-Orc1 | Positive for OFV-Gen, & OFV-Orc2 | |
| 3 | FL6ASP | 19,575,807 | 19,424,805 | 119,926 | 118,513 | 1413 | 12,841 | 0.62 | 98.82 | 0.00610 | OFV-Orc2 & New OFV Srtain OFV-Orc3 | True Positive for Dichorhavirus, OFV-Gen, & OFV-Orc, suspicious for OFV-Cit, OFV-Orc1 and OFV-Cit2 | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 | |
| 4 | FL7OHP | 21,964,465 | 21,801,326 | 1556 | 372 | 1184 | 4707 | 0.01 | 23.91 | 1.71 × 10−5 | New OFV Srtain OFV-Orc3 | True Positive for Dichorhavirus, OFV-Gen, & OFV-Orc1, suspicious for OFV-Orc | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 | |
| 5 | S1-TH | 122,785,064 | 121,388,722 | 7,716,120 | 3,047,882 | 4,668,238 | 18,378 | 6.36 | 39.50 | 0.02511 | New OFV Srtain OFV-Orc3 | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc, OFV-Orc1, & OFV-Orc2, suspicious for OFV-Cit | Positive for OFV-Gen | |
| 6 | S2-GA | 91,745,716 | 90,743,284 | 6,963,254 | 6,370,557 | 592,697 | 12,463 | 7.67 | 91.49 | 0.07020 | OFV Srtain-Orc2 | True Positive for Dichorhavirus, OFV-Gen, OFV-Cit, OFV-Orc & OFV-Orc2 | Positive for OFV-Gen, & OFV-Orc2 | |
| 7 | CA1 | 22,261,865 | 22,116,296 | 3,013,247 | 3,009,450 | 3797 | 12,479 | 13.63 | 99.87 | 0.13607 | OFV Srtain-Orc2 | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc, & OFV-Orc2 | Positive for OFV-Gen, & OFV-Orc2 | |
| 8 | CA2 | 20,763,435 | 20,598,618 | 1,269,615 | 1,268,336 | 1279 | 18,662 | 6.16 | 99.90 | 0.06157 | OFV Srtain-Orc1 & Orc2 | True Positive for Dichorhavirus, OFV-Gen, OFV-Cit, OFV-Orc, OFV-Orc1 & OFV-Orc2 and suspicious for OFV-Cit2 | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 | |
| 9 | CA3 | 23,475,312 | 23,312,790 | 13,451 | 9742 | 3709 | 14,933 | 0.06 | 72.43 | 0.00042 | OFV Srtain-Orc1 & Orc2 | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc1 & OFV-Orc2 and suspicious for OFV-Orc | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 | |
| 10 | CA4 | 19,604,543 | 19,458,011 | 1,476,793 | 1,475,932 | 861 | 12,506 | 7.59 | 99.94 | 0.07585 | OFV Srtain-Orc2 | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc & OFV-Orc2 | Positive for OFV-Gen, & OFV-Orc2 | |
| 11 | CA5 | 24,331,027 | 24,173,029 | 1,832,898 | 1,725,961 | 106,937 | 12,752 | 7.58 | 94.17 | 0.07140 | OFV Srtain-Orc2 | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc, OFV-Orc1 & OFV-Orc2 | Positive for OFV-Gen, OFV-Orc1 & OFV-Orc2 | |
| 12 | Orc2-HI | 50,962,963 | 48,290,861 | 6,721,283 | 2,540,314 | 4,180,969 | 12,467 | 13.92 | 37.80 | 0.05260 | OFV Srtain-Orc2 | True Positive for Dichorhavirus, OFV-Gen, OFV-Orc, & OFV-Orc2 | Positive for OFV-Gen, & OFV-Orc2 | |
| 13 | QR035 | Mexico | 26,721,386 | 26,542,292 | 5,955,476 | 862 | 5,954,614 | 8367 | 3.60 | 0.01 | 3.25 × 10−5 | OFV Srtain-Cit1 | True Positive for Dichorhavirus, OFV-Gen, OFV-Cit, & OFV-Cit1 | Positive for OFV-Gen, & OFV-Cit1 |
| 14 | CO002 | 6,947,727 | 6,879,156 | 1,596,975 | 1,588,639 | 8336 | 25,439 | 23.22 | 99.48 | 0.23094 | Novel nucleo rhabdovirus | True Positive for Dichorhavirus and suspicious for OFV-Gen | Positive for OFV-Gen | |
| 15 | CO003 | 17,110,470 | 16,983,502 | 14,635,242 | 0 | 14,635,242 | 0 | 86.17 | 0.00 | 0 | Negative to any rhabdovirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 16 | CO004 | 17,427,101 | 17,286,898 | 4358 | 0 | 4358 | 0 | 0.03 | 0.00 | 0 | Negative to any rhabdovirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 17 | CO005ac | 22,162,878 | 21,989,748 | 6,843,912 | 2,454,149 | 4,389,763 | 12,766 | 31.12 | 35.86 | 0.11160 | Novel dichorhavirus | Suspicious for dichorhavirus as infected with Novel dichorhavirus | Positive for OFV-Gen | |
| 18 | CO005bd | 23,278,875 | 23,102,829 | 9,897,923 | 0 | 9,897,923 | 0 | 42.84 | 0.00 | 0 | Negative to any rhabdovirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 19 | CO005e | 24,703,710 | 24,520,564 | 7,404,784 | 0 | 7,404,784 | 0 | 30.20 | 0.00 | 0 | Negative to any rhabdovirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 20 | JA021 | 20,346,774 | 20,193,394 | 1338 | 0 | 1338 | 0 | 0.01 | 0.00 | 0 | No virus detected | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 21 | JA022 | 20,685,498 | 20,523,285 | 263,176 | 255,960 | 7216 | 18,481 | 1.28 | 97.26 | 0.01247 | OFV Srtain-Cit2 | True Positive for Dichorhavirus, OFV-Gen, OFV-Cit, & OFV-Cit2 and Suspicious for OFV-Orc2 | Positive for OFV-Gen, & OFV-Cit2 | |
| 22 | JA023 | 17,124,047 | 16,929,790 | 147 | 0 | 147 | 0 | 0.00 | 0.00 | 0 | No virus (Healthy like) | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 23 | TA018 | 15,660,539 | 15,459,953 | 269,342 | 255,846 | 13,496 | 12,551 | 17.38 | 94.99 | 0.01655 | Novel dichorhavirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 24 | TA019 | 22,257,306 | 22,092,419 | 574,354 | 768 | 573,586 | 10,532 | 2.60 | 0.13 | 3.48 × 10−5 | Negative to any rhabdovirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 25 | S78_VDPCU | Colombia | 78,148,248 | 77,029,412 | 62,935,224 | 3,754,190 | 59,181,034 | 12,962 | 81.70 | 5.97 | 0.04874 | Novel dichorhavirus | True Negative for all the e-probes | Negative for tested dichorhavirus species |
| 26 | S79_VOrBTP | 26,126,253 | 25,266,822 | 14,241,854 | 10,766,019 | 3,475,835 | 12,515 | 56.37 | 75.59 | 0.42609 | Novel dichorhavirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 27 | S80_VOrP | 38,962,137 | 38,028,459 | 108,237 | 4724 | 103,513 | 12,460 | 0.28 | 4.37 | 0.00012 | Novel dichorhavirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 28 | S84_VBAP | 80,430,198 | 79,341,663 | 1,760,691 | 1,657,883 | 102,808 | 12,510 | 2.22 | 94.16 | 0.02090 | Novel dichorhavirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 29 | CR1 | Costa Rica | 31,309,832 | 31,105,043 | 14,052 | 0 | 14,052 | 0 | 0.05 | 0.00 | 0 | Negative to any rhabdovirus | True Negative for all the e-probes | Negative for tested dichorhavirus species |
| 30 | CR2 | 26,277,825 | 26,098,518 | 368,003 | 0 | 368,003 | 0 | 1.41 | 0.00 | 0 | Negative to any rhabdovirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| 31 | CR3 | 9,538,604 | 9,374,241 | 72,214 | 0 | 72,214 | 0 | 0.77 | 0.00 | 0 | Negative to any rhabdovirus | True Negative for all the e-probes | Negative for tested dichorhavirus species | |
| Sl. No | Sample | Dichorhavirus | e-Probe Size Used in EDNA Detection (nt) | Method Used for Analysis | EDNA Diagnostic as Compare to RT-qPCR Only | Changed in RT-qPCR Detection after Bioinformatic Analysis | EDNA Diagnostic as Compare to RT-qPCR and Bioinformatic Analysis | EDNA Diagnosis | Overall Comment on e-Probe Specificity Based on Analysis of Atleast Two Methods | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genus/ Species/Strain/ Variant | True Positive (TP) | True Negative (TN) | False Positive (FP) | False Negative (FN) | True Positive (TP) | True Negative (TN) | False Positive (FP) | False Negative (FN) | ||||||||||
| 40 | 60 | 80 | RT-qPCR (CT Value) | Bioinformatic Pipeline | ||||||||||||||
| 1 | ASP-FL (Aspidistra sp.) | OFV_Orc2 | NEG | NEG | No probe | 33.26 | NEG | 1 | TP to TN | 1 | Negative | Correct | ||||||
| 2 | FL6ASP (Aspidistra sp.) | OFV_Orc1 | NEG | NEG | NEG | 31.56 | NEG | 1 | TP to TN | 1 | Negative | Correct | ||||||
| OFV-Orc | POS | No probe | No probe | NT | POS | Inconsistent (IC) | IC to TP | 1 | Positive | Correct | ||||||||
| OFV_Orc2 | NEG | NEG | No probe | 27.71 | POS | 1 | TP to TP (No change) | 1 | False negative | Incorrect | ||||||||
| 3 | FL7OHP (Ophiopogan sp.) | Dichorhavirus | POS | NEG | NEG | NT | POS | Inconsistent | IC to TP | 1 | Positive | Correct | ||||||
| OFV_Orc1 | POS | NEG | NEG | 31.37 | NEG | 1 | TP to TN | 1 | Positive | Correct | ||||||||
| OFV_Orc2 | NEG | NEG | No probe | 29.12 | NEG | 1 | TP to TN | 1 | Negative | Correct | ||||||||
| 4 | S1 (Liriope-TH) | OFV_Orc1 | POS | POS | POS | UD | NEG | 1 | TN to TN (No change) | 1 | False positive | Incorrect | ||||||
| OFV_Orc2 | POS | NEG | No probe | UD | NEG | 1 | TN to TN (No change) | 1 | False positive | Incorrect | ||||||||
| 5 | S2 (Liriope-GA) | OFV_Orc2 | POS | NEG | No probe | 18.52 | POS | 1 | TP to TP (No change) | 1 | Positive | Correct | ||||||
| 6 | OFV-Hawaii (Citrus reticulata) | OFV_Orc2 | POS | NEG | NEG | 25.94 | POS | 1 | TP to TP (No change) | 1 | Positive | Correct | ||||||
| 7 | CA1 (Cymbidium sp.) | OFV_Orc2 | POS | NEG | NEG | 17.08 | POS | 1 | TP to TP (No change) | 1 | Positive | Correct | ||||||
| 8 | CA2 (Dendrobium sp.) | OFV-Cit | POS | No probe | No probe | NT | NEG | Inconsistent | IC to TN | 1 | False positive | Incorrect | ||||||
| OFV_Orc2 | NEG | NEG | No probe | 18.66 | POS | 1 | TP to TP (No change) | 1 | Positive | Correct | ||||||||
| 9 | CA3 (Dendrobium sp.) | OFV-Orc | NEG | No probe | No probe | NT | POS | Inconsistent | IC to TP | 1 | False negative | Incorrect | ||||||
| OFV_Orc2 | POS | NEG | No probe | 22.99 | POS | 1 | TP to TP (No change) | 1 | Positive | Correct | ||||||||
| 10 | CA4 (Cymbidium sp.) | OFV_Orc2 | POS | NEG | No probe | 18.97 | POS | 1 | TP to TP (No change) | 1 | Positive | Correct | ||||||
| 11 | CA5 (Dendrochilum magnum) | OFV_Orc1 | POS | NEG | NEG | 30.44 | NEG | 1 | TP to TP (No change) | 1 | Positive | Correct | ||||||
| OFV_Orc2 | POS | NEG | No probe | 14.56 | POS | 1 | TP to TP (No change) | 1 | Positive | Correct | ||||||||
| 12 | CO002 (Aralia sp.) | Dichorhavirus | NEG | POS | NEG | NT | NEG | Inconsistent | IC to TN | 1 | False Positive, reacted with Novel Nucleo-rhabdovirus sequence | Incorrect | ||||||
| OFV_Orc1 | POS | POS | NEG | UD | NEG | 1 | TN to TN (No change) | 1 | False positive | Incorrect | ||||||||
| 13 | CO005ac (Dendrobium sp.) | Dichorhavirus | NEG | NEG | NEG | NT | POS | Inconsistent | IC to TP | 1 | False Negative, reacted with Novel Dichorhavirus species sequence | Incorrect | ||||||
| OFV-Gen | NEG | NEG | NEG | 30.52 | NEG | 1 | TP to TN | 1 | Negative | Correct | ||||||||
| OFV_Cit2 | NEG | NEG | NEG | 34.74 | NEG | 1 | FP to TN | 1 | Negative | Correct | ||||||||
| OFV_Cit1 | NEG | NEG | NEG | 33.63 | NEG | 1 | TP to TN | 1 | Negative | Correct | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, A.; Shao, J.; Espindola, A.S.; Ramos Lopez, D.; Otero-Colina, G.; Rivera, Y.; Mavrodieva, V.A.; Nakhla, M.K.; Schneider, W.L.; Cardwell, K. Detection and In Vivo Validation of Dichorhavirus e-Probes in Meta-Transcriptomic Data via Microbe Finder (MiFi®) Discovers a Novel Host and a Possible New Strain of Orchid Fleck Virus. Viruses 2025, 17, 441. https://doi.org/10.3390/v17030441
Roy A, Shao J, Espindola AS, Ramos Lopez D, Otero-Colina G, Rivera Y, Mavrodieva VA, Nakhla MK, Schneider WL, Cardwell K. Detection and In Vivo Validation of Dichorhavirus e-Probes in Meta-Transcriptomic Data via Microbe Finder (MiFi®) Discovers a Novel Host and a Possible New Strain of Orchid Fleck Virus. Viruses. 2025; 17(3):441. https://doi.org/10.3390/v17030441
Chicago/Turabian StyleRoy, Avijit, Jonathan Shao, Andres S. Espindola, Daniel Ramos Lopez, Gabriel Otero-Colina, Yazmín Rivera, Vessela A. Mavrodieva, Mark K. Nakhla, William L. Schneider, and Kitty Cardwell. 2025. "Detection and In Vivo Validation of Dichorhavirus e-Probes in Meta-Transcriptomic Data via Microbe Finder (MiFi®) Discovers a Novel Host and a Possible New Strain of Orchid Fleck Virus" Viruses 17, no. 3: 441. https://doi.org/10.3390/v17030441
APA StyleRoy, A., Shao, J., Espindola, A. S., Ramos Lopez, D., Otero-Colina, G., Rivera, Y., Mavrodieva, V. A., Nakhla, M. K., Schneider, W. L., & Cardwell, K. (2025). Detection and In Vivo Validation of Dichorhavirus e-Probes in Meta-Transcriptomic Data via Microbe Finder (MiFi®) Discovers a Novel Host and a Possible New Strain of Orchid Fleck Virus. Viruses, 17(3), 441. https://doi.org/10.3390/v17030441