
Bunyaviral Cap-Snatching Endonuclease Activity and Inhibition with Baloxavir-like Inhibitors in the Context of Full-Length L Proteins

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Protein Expression and Purification
2.3. Biochemical Assays
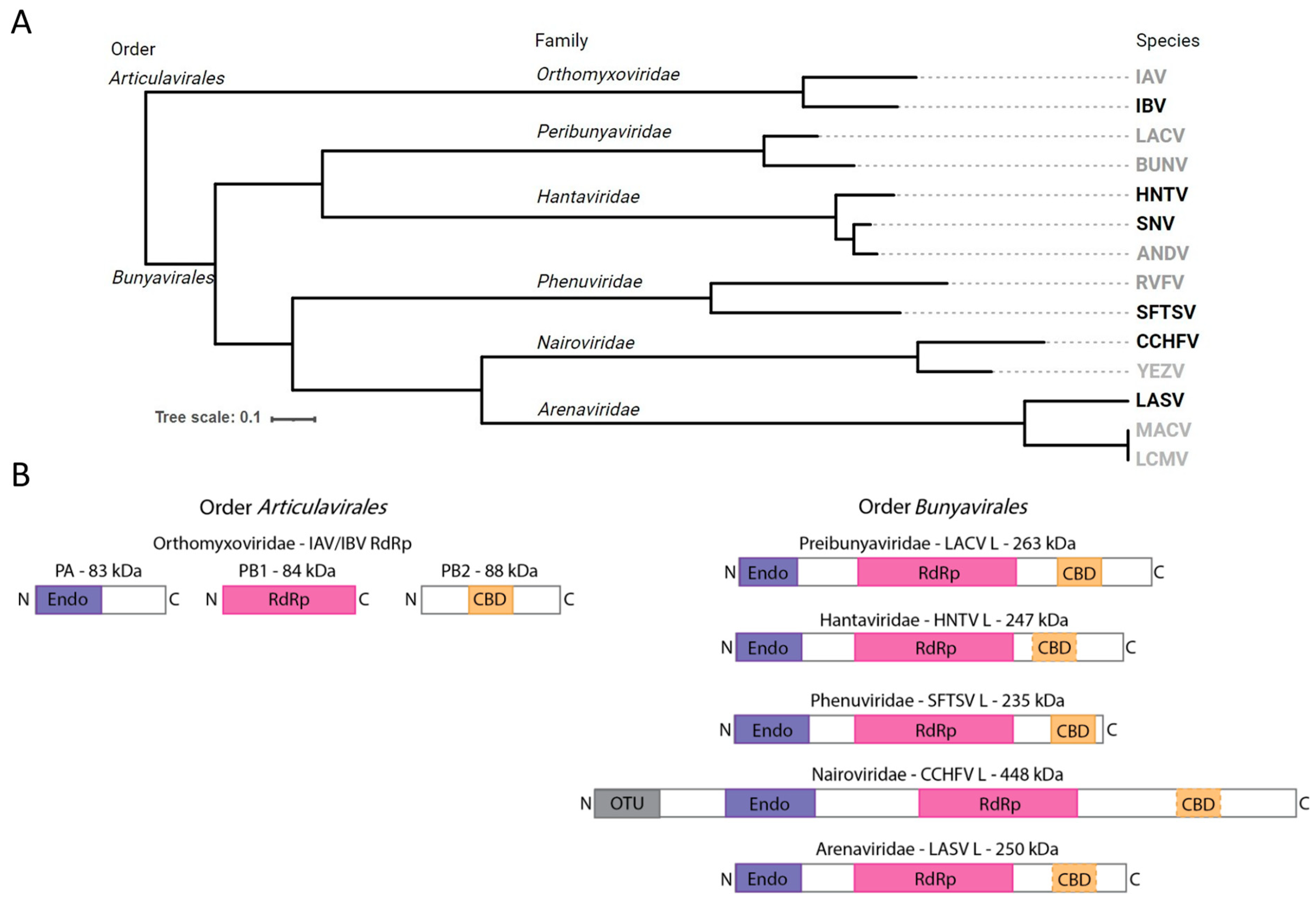
2.4. Phylogenetic Analysis and Design of RdRp Functional Domain Diagram
3. Results
3.1. Expression and Purification of Bunyaviral L Proteins
3.2. Purified Proteins Demonstrate RNA Synthesis Activity
3.3. Bunyaviral L Protein Cleavage Patterns Differ from the Specific IBV Endonuclease Cut
3.3.1. Cleavage of Capped Substrates Lacks Strict Sequence Specificity
3.3.2. Specific Cap-Dependent Cleavage Is Diminished with Bunyaviral Enzymes
3.3.3. BXA and Derivatives Can Inhibit Bunyaviral Endonuclease Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abudurexiti, A.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Avšič-Županc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, É.; Blair, C.D.; et al. Taxonomy of the order Bunyavirales: Update 2019. Arch. Virol. 2019, 164, 1949–1965. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Morens, D.M. Influenza: The once and future pandemic. Public Health Rep. 2010, 125 (Suppl. S3), 16–26. [Google Scholar] [CrossRef]
- Ziegler, T.; Mamahit, A.; Cox, N.J. 65 years of influenza surveillance by a World Health Organization-coordinated global network. Influenza Other Respir. Viruses 2018, 12, 558–565. [Google Scholar] [CrossRef]
- Dumm, R.E.; Fiege, J.K.; Waring, B.M.; Kuo, C.T.; Langlois, R.A.; Heaton, N.S. Non-lytic clearance of influenza B virus from infected cells preserves epithelial barrier function. Nat. Commun. 2019, 10, 779. [Google Scholar] [CrossRef] [PubMed]
- Tompa, D.R.; Immanuel, A.; Srikanth, S.; Kadhirvel, S. Trends and strategies to combat viral infections: A review on FDA approved antiviral drugs. Int. J. Biol. Macromol. 2021, 172, 524–541. [Google Scholar] [CrossRef] [PubMed]
- Olschewski, S.; Cusack, S.; Rosenthal, M. The Cap-Snatching Mechanism of Bunyaviruses. Trends Microbiol. 2020, 28, 293–303. [Google Scholar] [CrossRef]
- Wandzik, J.M.; Kouba, T.; Karuppasamy, M.; Pflug, A.; Drncova, P.; Provaznik, J.; Azevedo, N.; Cusack, S. A Structure-Based Model for the Complete Transcription Cycle of Influenza Polymerase. Cell 2020, 181, 877–893.e21. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.; Bouvier, D.; Crépin, T.; McCarthy, A.A.; Hart, D.J.; Baudin, F.; Cusack, S.; Ruigrok, R.W.H. The cap-snatching endonuclease of influenza virus polymerase resides in the PA subunit. Nature 2009, 458, 914–918. [Google Scholar] [CrossRef]
- Reich, S.; Guilligay, D.; Pflug, A.; Malet, H.; Berger, I.; Crépin, T.; Hart, D.; Lunardi, T.; Nanao, M.; Ruigrok, R.W.H.; et al. Structural insight into cap-snatching and RNA synthesis by influenza polymerase. Nature 2014, 516, 361–366. [Google Scholar] [CrossRef]
- Ferron, F.; Weber, F.; De La Torre, J.C.; Reguera, J. Transcription and replication mechanisms of Bunyaviridae and Arenaviridae L proteins. Virus Res. 2017, 234, 118–134. [Google Scholar] [CrossRef]
- Lelke, M.; Brunotte, L.; Busch, C.; Günther, S. An N-terminal region of Lassa virus L protein plays a critical role in transcription but not replication of the virus genome. J. Virol. 2010, 84, 1934–1944. [Google Scholar] [CrossRef] [PubMed]
- Reguera, J.; Gerlach, P.; Rosenthal, M.; Gaudon, S.; Coscia, F.; Günther, S.; Cusack, S. Comparative Structural and Functional Analysis of Bunyavirus and Arenavirus Cap-Snatching Endonucleases. PLoS Pathog. 2016, 12, e1005636. [Google Scholar] [CrossRef]
- Gogrefe, N.; Reindl, S.; Günther, S.; Rosenthal, M. Structure of a functional cap-binding domain in Rift Valley fever virus L protein. PLoS Pathog. 2019, 15, e1007829. [Google Scholar] [CrossRef] [PubMed]
- Meier, K.; Thorkelsson, S.R.; Durieux Trouilleton, Q.; Vogel, D.; Yu, D.; Kosinski, J.; Cusack, S.; Malet, H.; Grünewald, K.; Quemin, E.R.J.; et al. Structural and functional characterization of the Sin Nombre virus L protein. PLoS Pathog. 2023, 19, e1011533. [Google Scholar] [CrossRef] [PubMed]
- Vogel, D.; Thorkelsson, S.R.; Quemin, E.R.J.; Meier, K.; Kouba, T.; Gogrefe, N.; Busch, C.; Reindl, S.; Günther, S.; Cusack, S.; et al. Structural and functional characterization of the severe fever with thrombocytopenia syndrome virus L protein. Nucleic Acids Res. 2020, 48, 5749–5765. [Google Scholar] [CrossRef]
- Reguera, J.; Weber, F.; Cusack, S. Bunyaviridae RNA polymerases (L-protein) have an N-terminal, influenza-like endonuclease domain, essential for viral cap-dependent transcription. PLoS Pathog. 2010, 6, e1001101. [Google Scholar] [CrossRef]
- Yang, W. Nucleases: Diversity of structure, function and mechanism. Q. Rev. Biophys. 2011, 44, 1–93. [Google Scholar] [CrossRef]
- Kuang, W.; Zhang, H.; Cai, Y.; Zhang, G.; Deng, F.; Li, H.; Hu, Z.; Guo, Y.; Wang, M.; Zhou, Y.; et al. Insights into Two-Metal-Ion Catalytic Mechanism of Cap-Snatching Endonuclease of Ebinur Lake Virus in Bunyavirales. J. Virol. 2022, 96, e0208521. [Google Scholar] [CrossRef]
- Noshi, T.; Kitano, M.; Taniguchi, K.; Yamamoto, A.; Omoto, S.; Baba, K.; Hashimoto, T.; Ishida, K.; Kushima, Y.; Hattori, K.; et al. In vitro characterization of baloxavir acid, a first-in-class cap-dependent endonuclease inhibitor of the influenza virus polymerase PA subunit. Antivir. Res. 2018, 160, 109–117. [Google Scholar] [CrossRef]
- Hayden Frederick, G.; Sugaya, N.; Hirotsu, N.; Lee, N.; de Jong Menno, D.; Hurt Aeron, C.; Ishida, T.; Sekino, H.; Yamada, K.; Portsmouth, S.; et al. Baloxavir Marboxil for Uncomplicated Influenza in Adults and Adolescents. N. Engl. J. Med. 2018, 379, 913–923. [Google Scholar] [CrossRef]
- Ikematsu, H.; Hayden Frederick, G.; Kawaguchi, K.; Kinoshita, M.; de Jong Menno, D.; Lee, N.; Takashima, S.; Noshi, T.; Tsuchiya, K.; Uehara, T. Baloxavir Marboxil for Prophylaxis against Influenza in Household Contacts. N. Engl. J. Med. 2020, 383, 309–320. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. FDA Approves New Drug to Treat Influenza; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2018. [Google Scholar]
- Omoto, S.; Speranzini, V.; Hashimoto, T.; Noshi, T.; Yamaguchi, H.; Kawai, M.; Kawaguchi, K.; Uehara, T.; Shishido, T.; Naito, A.; et al. Characterization of influenza virus variants induced by treatment with the endonuclease inhibitor baloxavir marboxil. Sci. Rep. 2018, 8, 9633. [Google Scholar] [CrossRef] [PubMed]
- Todd, B.; Tchesnokov, E.P.; Götte, M. The active form of the influenza cap-snatching endonuclease inhibitor baloxavir marboxil is a tight binding inhibitor. J. Biol. Chem. 2021, 296, 100486. [Google Scholar] [CrossRef]
- Kumar, G.; Cuypers, M.; Webby, R.R.; Webb, T.R.; White, S.W. Structural insights into the substrate specificity of the endonuclease activity of the influenza virus cap-snatching mechanism. Nucleic Acids Res. 2021, 49, 1609–1618. [Google Scholar] [CrossRef]
- Feracci, M.; Hernandez, S.; Garlatti, L.; Mondielli, C.; Vincentelli, R.; Canard, B.; Reguera, J.; Ferron, F.; Alvarez, K. Biophysical and structural study of La Crosse virus endonuclease inhibition for the development of new antiviral options. IUCrJ 2024, 11 Pt 3, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Lessoued, S.; Meier, K.; Devignot, S.; Barata-García, S.; Mate, M.; Bragagnolo, G.; Weber, F.; Rosenthal, M.; Reguera, J. Structure and function of the Toscana virus cap-snatching endonuclease. Nucleic Acids Res. 2019, 47, 10914–10930. [Google Scholar] [CrossRef] [PubMed]
- Kuang, W.; Zhang, H.; Cai, Y.; Zhang, G.; Deng, F.; Li, H.; Zhou, Y.; Wang, M.; Gong, P.; Guo, Y.; et al. Structural and Biochemical Basis for Development of Diketo Acid Inhibitors Targeting the Cap-Snatching Endonuclease of the Ebinur Lake Virus (Order: Bunyavirales). J. Virol. 2022, 96, e0217321. [Google Scholar] [CrossRef]
- Toba, S.; Sato, A.; Kawai, M.; Taoda, Y.; Unoh, Y.; Kusakabe, S.; Nobori, H.; Uehara, S.; Uemura, K.; Taniguchi, K.; et al. Identification of cap-dependent endonuclease inhibitors with broad-spectrum activity against bunyaviruses. Proc. Natl. Acad. Sci. USA 2022, 119, e2206104119. [Google Scholar] [CrossRef]
- Taoda, Y.; Sato, A.; Toba, S.; Unoh, Y.; Kawai, M.; Sasaki, M.; Orba, Y.; Sawa, H. Structure-activity relationship studies of anti-bunyaviral cap-dependent endonuclease inhibitors. Bioorganic Med. Chem. Lett. 2023, 83, 129175. [Google Scholar] [CrossRef] [PubMed]
- Holm, T.; Kopicki, J.-D.; Busch, C.; Olschewski, S.; Rosenthal, M.; Uetrecht, C.; Günther, S.; Reindl, S. Biochemical and structural studies reveal differences and commonalities among cap-snatching endonucleases from segmented negative-strand RNA viruses. J. Biol. Chem. 2018, 293, 19686–19698. [Google Scholar] [CrossRef]
- Williams, H.M.; Thorkelsson, S.R.; Vogel, D.; Busch, C.; Milewski, M.; Cusack, S.; Grünewald, K.; Quemin, E.R.J.; Rosenthal, M. Structural snapshots of phenuivirus cap-snatching and transcription. Nucleic Acids Res. 2024, 52, 6049–6065. [Google Scholar] [PubMed]
- Keown, J.R.; Carrique, L.; Nilsson-Payant, B.E.; Fodor, E.; Grimes, J.M. Structural characterization of the full-length Hantaan virus polymerase. PLoS Pathog. 2024, 20, e1012781. [Google Scholar] [CrossRef]
- Berger, I.; Fitzgerald, D.J.; Richmond, T.J. Baculovirus expression system for heterologous multiprotein complexes. Nat. Biotechnol. 2004, 22, 1583–1587. [Google Scholar] [CrossRef] [PubMed]
- Bieniossek, C.; Richmond, T.J.; Berger, I. MultiBac: Multigene baculovirus-based eukaryotic protein complex production. Curr. Protoc. Protein Sci. 2008, 51, 5–20. [Google Scholar]
- Tchesnokov, E.P.; Bailey-Elkin, B.A.; Mark, B.L.; Götte, M. Independent inhibition of the polymerase and deubiquitinase activities of the Crimean-Congo Hemorrhagic Fever Virus full-length L-protein. PLoS Neglected Trop. Dis. 2020, 14, e0008283. [Google Scholar] [CrossRef]
- Tchesnokov, E.P.; Raeisimakiani, P.; Ngure, M.; Marchant, D.; Götte, M. Recombinant RNA-Dependent RNA Polymerase Complex of Ebola Virus. Sci. Rep. 2018, 8, 3970. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.Y.; Shoichet, B.K. A detergent-based assay for the detection of promiscuous inhibitors. Nat. Protoc. 2006, 1, 550–553. [Google Scholar] [CrossRef]
- Li, C.X.; Shi, M.; Tian, J.H.; Lin, X.D.; Kang, Y.J.; Chen, L.J.; Qin, X.C.; Xu, J.; Holmes, E.C.; Zhang, Y.Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. eLife 2015, 4, e05378. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef]
- Durieux Trouilleton, Q.; Barata-García, S.; Arragain, B.; Reguera, J.; Malet, H. Structures of active Hantaan virus polymerase uncover the mechanisms of Hantaviridae genome replication. Nat. Commun. 2023, 14, 2954. [Google Scholar] [CrossRef] [PubMed]
- Kouba, T.; Vogel, D.; Thorkelsson, S.R.; Quemin, E.R.J.; Williams, H.M.; Milewski, M.; Busch, C.; Günther, S.; Grünewald, K.; Rosenthal, M.; et al. Conformational changes in Lassa virus L protein associated with promoter binding and RNA synthesis activity. Nat. Commun. 2021, 12, 7018. [Google Scholar] [CrossRef] [PubMed]
- Beljelarskaya, S.N. Baculovirus expression systems for production of recombinant proteins in insect and mammalian cells. Mol. Biol. 2011, 45, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Vogel, D.; Rosenthal, M.; Gogrefe, N.; Reindl, S.; Günther, S. Biochemical characterization of the Lassa virus L protein. J. Biol. Chem. 2019, 294, 8088–8100. [Google Scholar] [CrossRef]
- Pflug, A.; Guilligay, D.; Reich, S.; Cusack, S. Structure of influenza A polymerase bound to the viral RNA promoter. Nature 2014, 516, 355–360. [Google Scholar] [CrossRef]
- Chang, S.; Sun, D.; Liang, H.; Wang, J.; Li, J.; Guo, L.; Wang, X.; Guan, C.; Boruah, B.M.; Yuan, L.; et al. Cryo-EM structure of influenza virus RNA polymerase complex at 4.3 Å resolution. Mol. Cell 2015, 57, 925–935. [Google Scholar] [CrossRef]
- Reich, S.; Guilligay, D.; Cusack, S. An in vitro fluorescence based study of initiation of RNA synthesis by influenza B polymerase. Nucleic Acids Res. 2017, 45, 3353–3368. [Google Scholar]
- Fan, H.; Walker, A.P.; Carrique, L.; Keown, J.R.; Serna Martin, I.; Karia, D.; Sharps, J.; Hengrung, N.; Pardon, E.; Steyaert, J.; et al. Structures of influenza A virus RNA polymerase offer insight into viral genome replication. Nature 2019, 573, 287–290. [Google Scholar] [CrossRef]
- Kouba, T.; Drncová, P.; Cusack, S. Structural snapshots of actively transcribing influenza polymerase. Nat. Struct. Mol. Biol. 2019, 26, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Peng, R.; Peng, Q.; Wang, M.; Xu, Y.; Liu, S.; Tian, X.; Deng, H.; Tong, Y.; Hu, X.; et al. Cryo-EM structures of Lassa and Machupo virus polymerases complexed with cognate regulatory Z proteins identify targets for antivirals. Nat. Microbiol. 2021, 6, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Williams, H.M.; Thorkelsson, S.R.; Vogel, D.; Milewski, M.; Busch, C.; Cusack, S.; Grünewald, K.; Quemin, E.R.J.; Rosenthal, M. Structural insights into viral genome replication by the severe fever with thrombocytopenia syndrome virus L protein. Nucleic Acids Res. 2023, 51, 1424–1442. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Chang, T.; Li, Z.; Wang, C.; Zhao, H.; Li, M.; Tang, P.; Wen, X.; Yu, M.; Wu, J.; et al. Cryo-EM structures of Thogoto virus polymerase reveal unique RNA transcription and replication mechanisms among orthomyxoviruses. Nat. Commun. 2024, 15, 4620. [Google Scholar] [CrossRef]
- Li, H.; Wu, Y.; Li, M.; Guo, L.; Gao, Y.; Wang, Q.; Zhang, J.; Lai, Z.; Zhang, X.; Zhu, L.; et al. An intermediate state allows influenza polymerase to switch smoothly between transcription and replication cycles. Nat. Struct. Mol. Biol. 2023, 30, 1183–1192. [Google Scholar] [CrossRef]
- Datta, K.; Wolkerstorfer, A.; Szolar, O.H.; Cusack, S.; Klumpp, K. Characterization of PA-N terminal domain of Influenza A polymerase reveals sequence specific RNA cleavage. Nucleic Acids Res. 2013, 41, 8289–8299. [Google Scholar] [CrossRef]
- Garcin, D.; Lezzi, M.; Dobbs, M.; Elliott, R.M.; Schmaljohn, C.; Kang, C.Y.; Kolakofsky, D. The 5′ ends of Hantaan virus (Bunyaviridae) RNAs suggest a prime-and-realign mechanism for the initiation of RNA synthesis. J. Virol. 1995, 69, 5754–5762. [Google Scholar] [CrossRef]
- Heinemann, P.; Schmidt-Chanasit, J.; Günther, S. The N Terminus of Andes Virus L Protein Suppresses mRNA and Protein Expression in Mammalian Cells. J. Virol. 2013, 87, 6975–6985. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loutan, A.J.; Yang, B.; Connolly, G.; Montoya, A.; Smiley, R.J.; Chatterjee, A.K.; Götte, M. Bunyaviral Cap-Snatching Endonuclease Activity and Inhibition with Baloxavir-like Inhibitors in the Context of Full-Length L Proteins. Viruses 2025, 17, 420. https://doi.org/10.3390/v17030420
Loutan AJ, Yang B, Connolly G, Montoya A, Smiley RJ, Chatterjee AK, Götte M. Bunyaviral Cap-Snatching Endonuclease Activity and Inhibition with Baloxavir-like Inhibitors in the Context of Full-Length L Proteins. Viruses. 2025; 17(3):420. https://doi.org/10.3390/v17030420
Chicago/Turabian StyleLoutan, Arlo J., Baiuyan Yang, Gabrielle Connolly, Adam Montoya, Robert J. Smiley, Arnab K. Chatterjee, and Matthias Götte. 2025. "Bunyaviral Cap-Snatching Endonuclease Activity and Inhibition with Baloxavir-like Inhibitors in the Context of Full-Length L Proteins" Viruses 17, no. 3: 420. https://doi.org/10.3390/v17030420
APA StyleLoutan, A. J., Yang, B., Connolly, G., Montoya, A., Smiley, R. J., Chatterjee, A. K., & Götte, M. (2025). Bunyaviral Cap-Snatching Endonuclease Activity and Inhibition with Baloxavir-like Inhibitors in the Context of Full-Length L Proteins. Viruses, 17(3), 420. https://doi.org/10.3390/v17030420