
Temporal and Spatial Analysis of Rabies Virus Lineages in South Africa


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Analysis of Animal Rabies Case Data
2.2. Analysis of Rabies Sequence Data
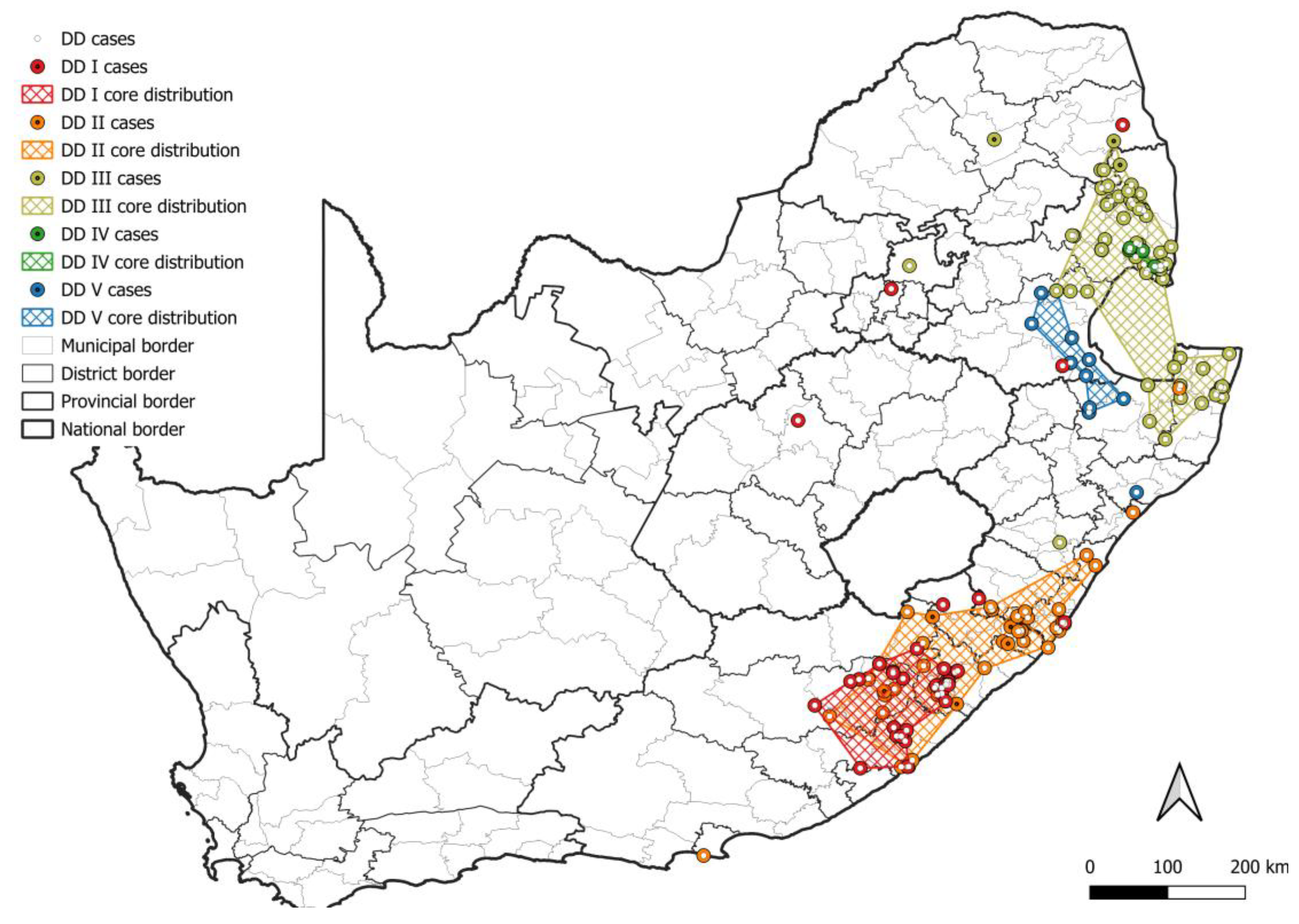
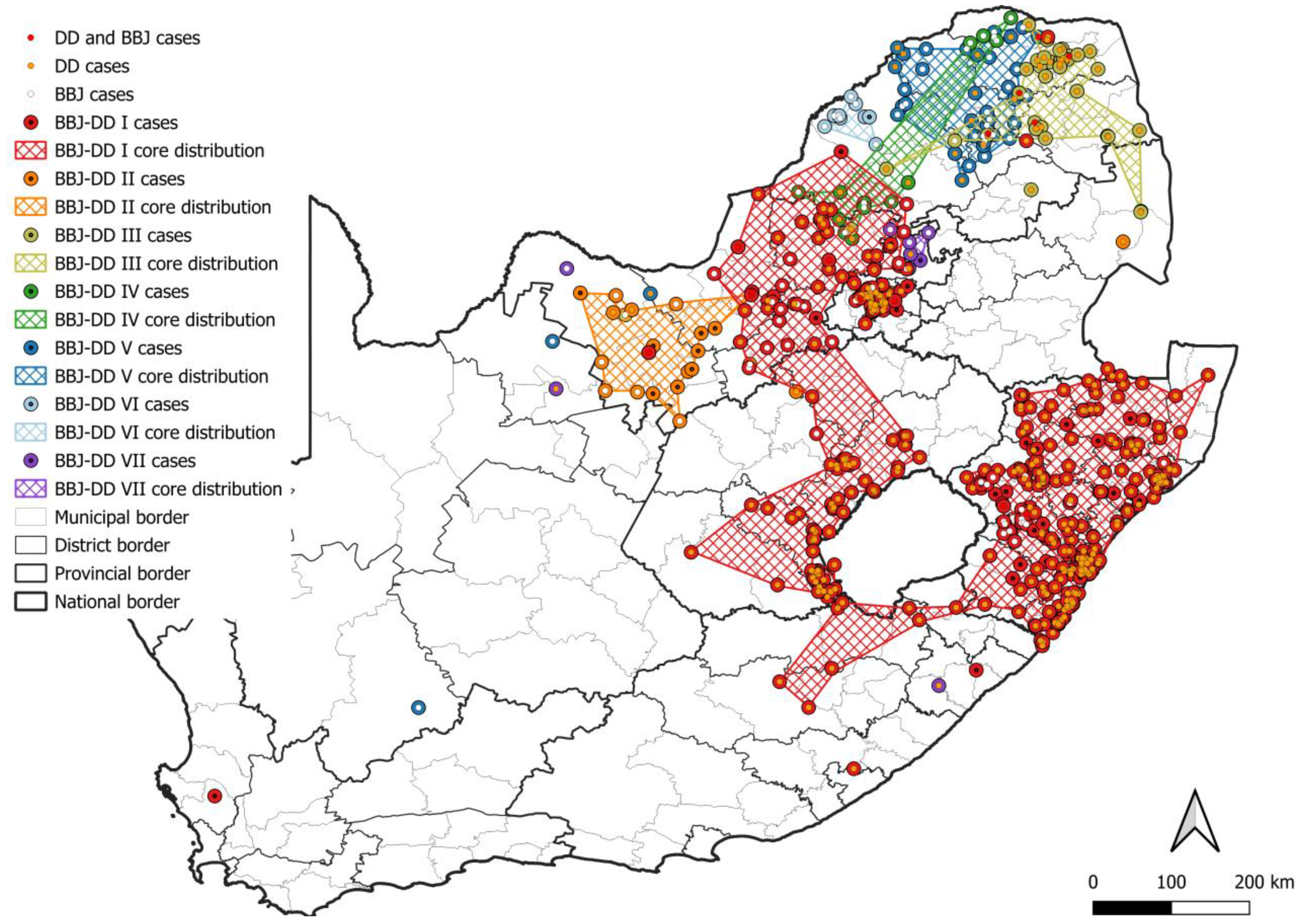
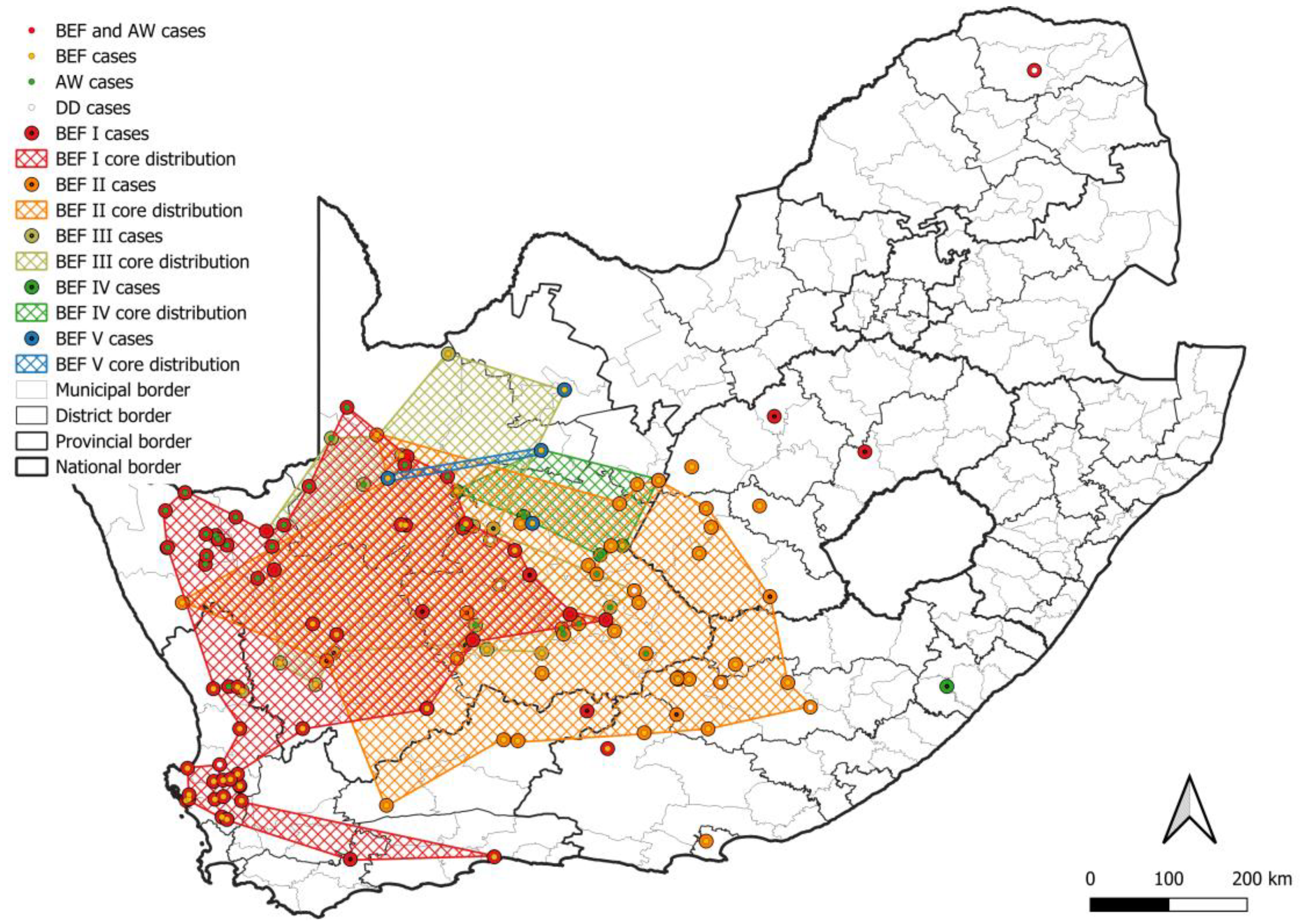
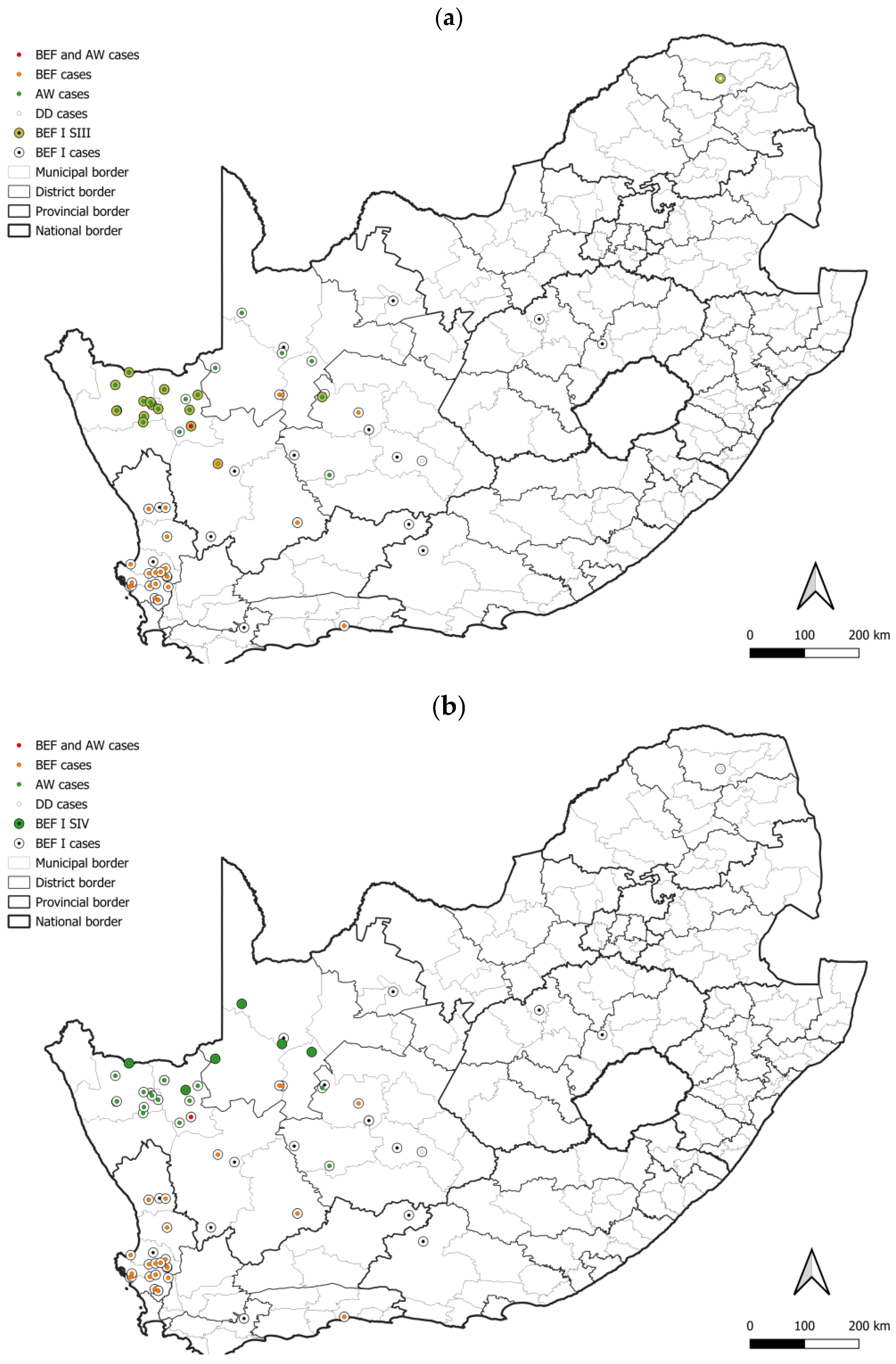
3. Results
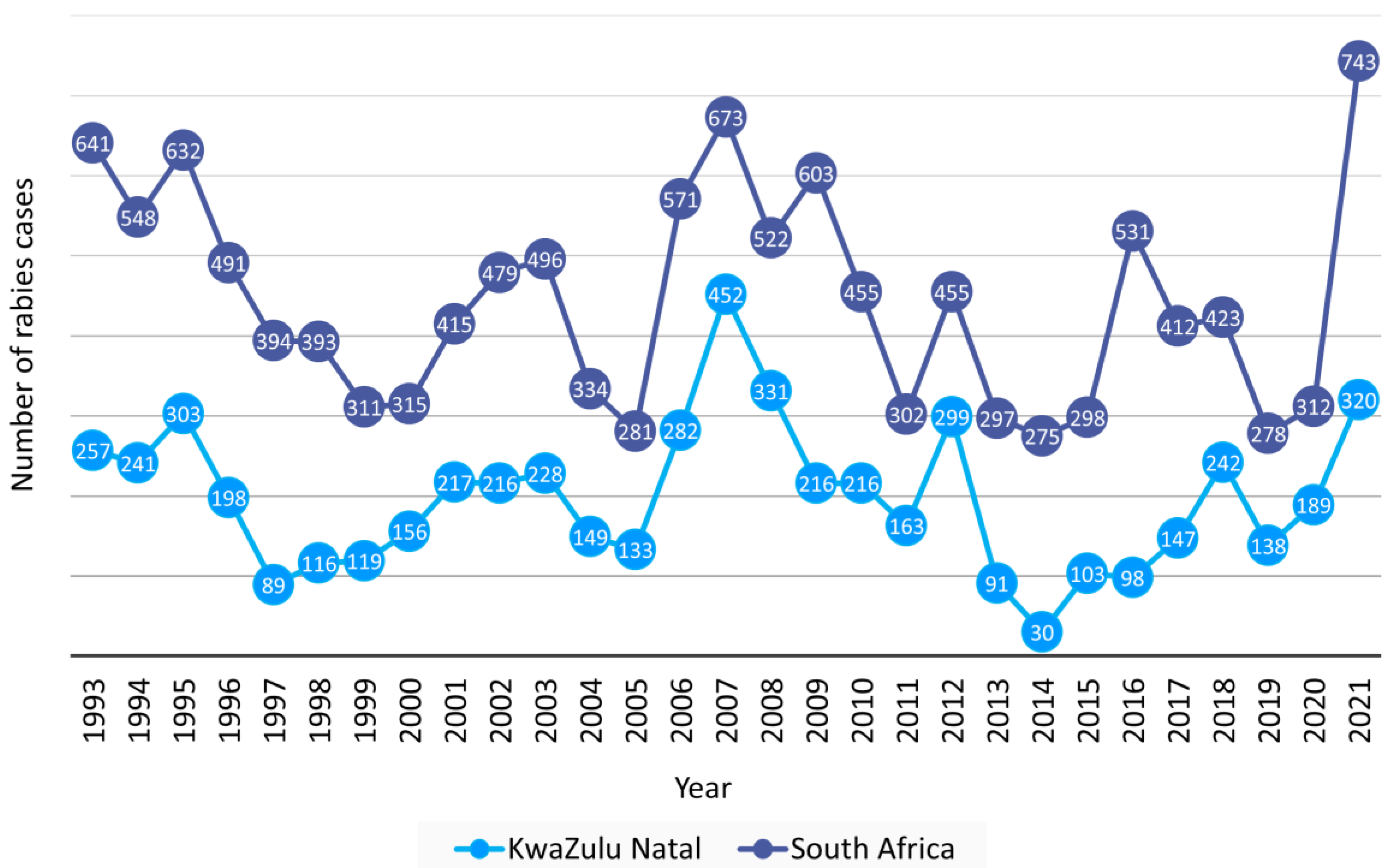
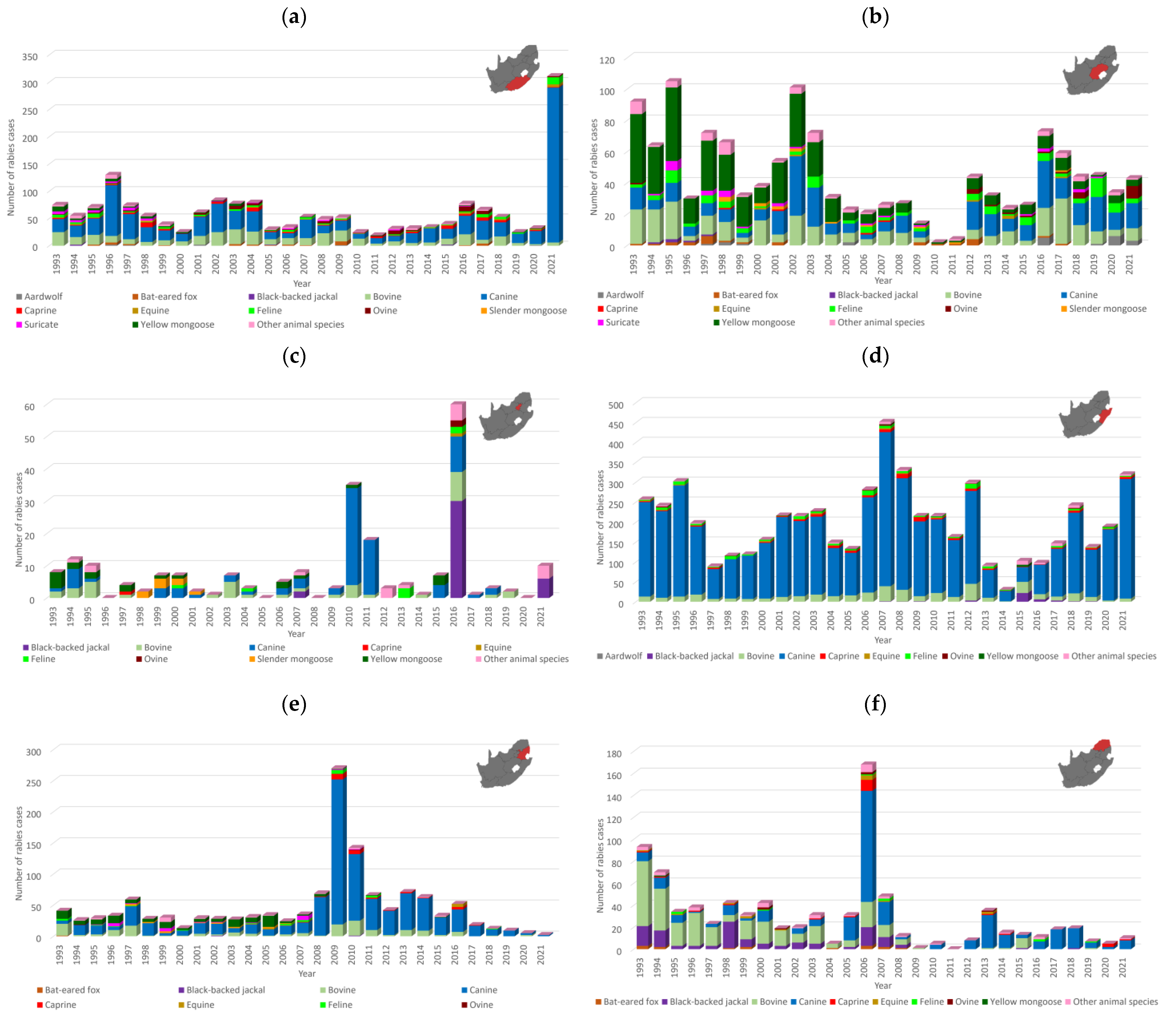
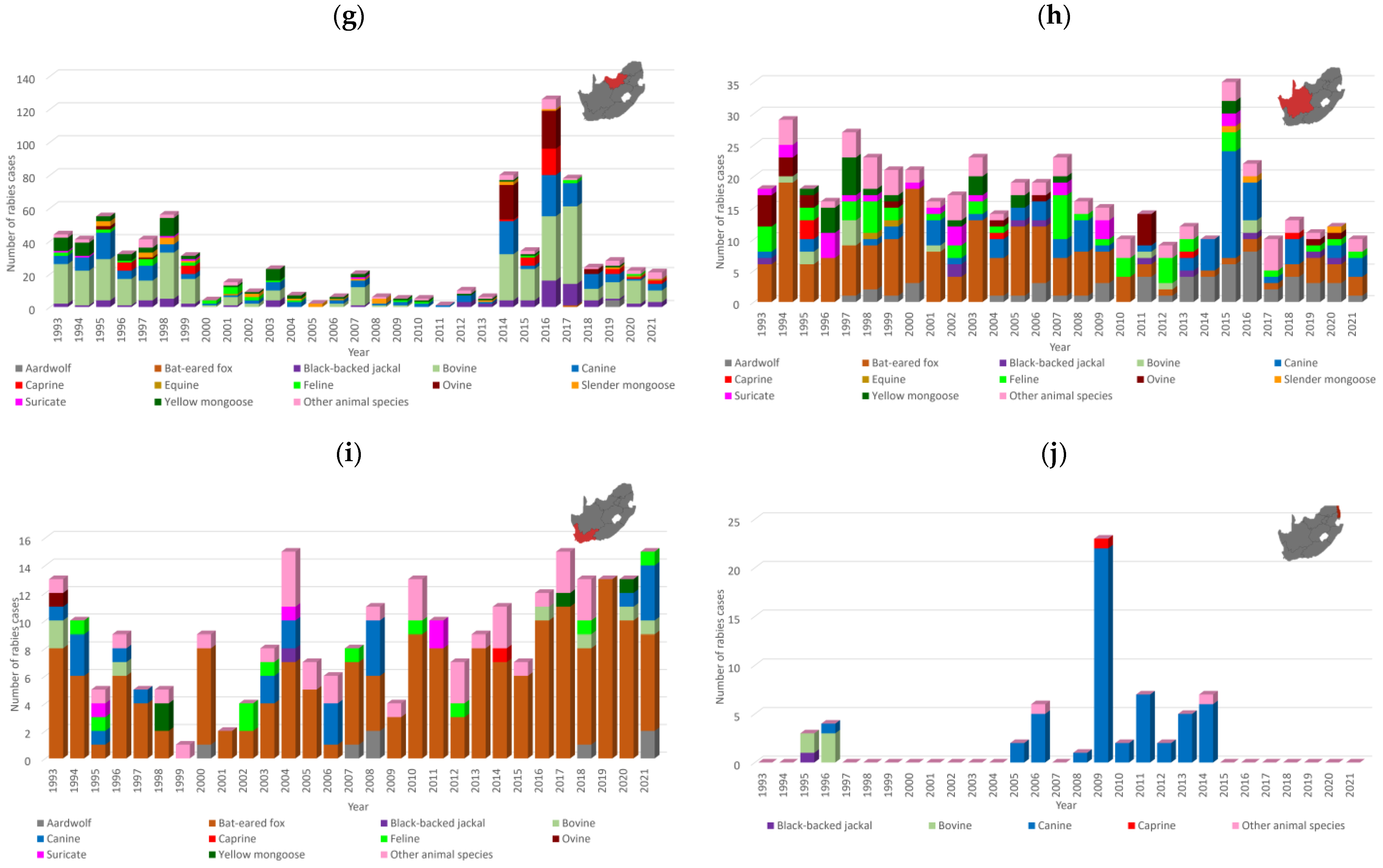
3.1. Number and Distribution of Laboratory-Confirmed Animal Rabies Cases Based on Surveillance Data
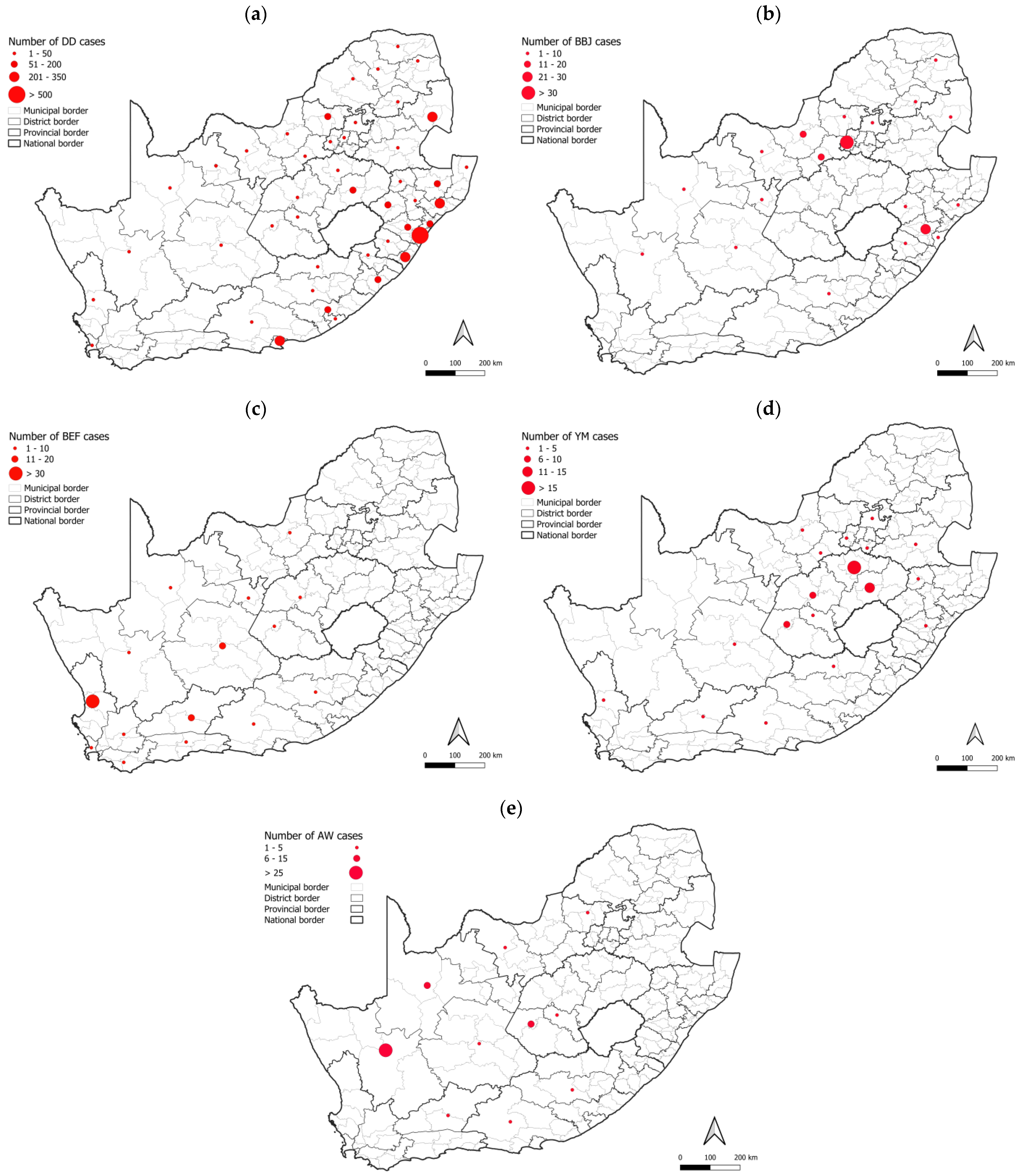
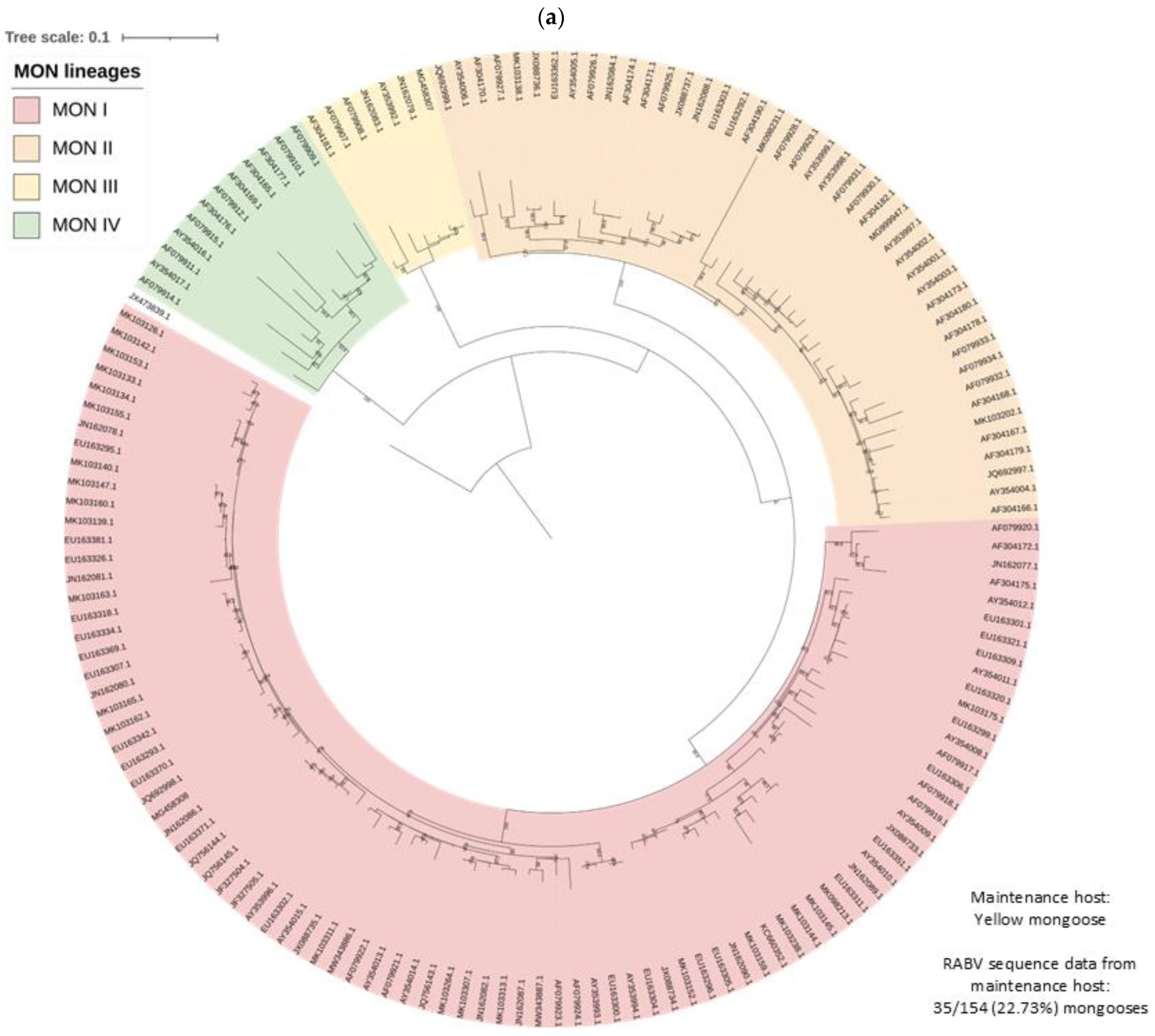
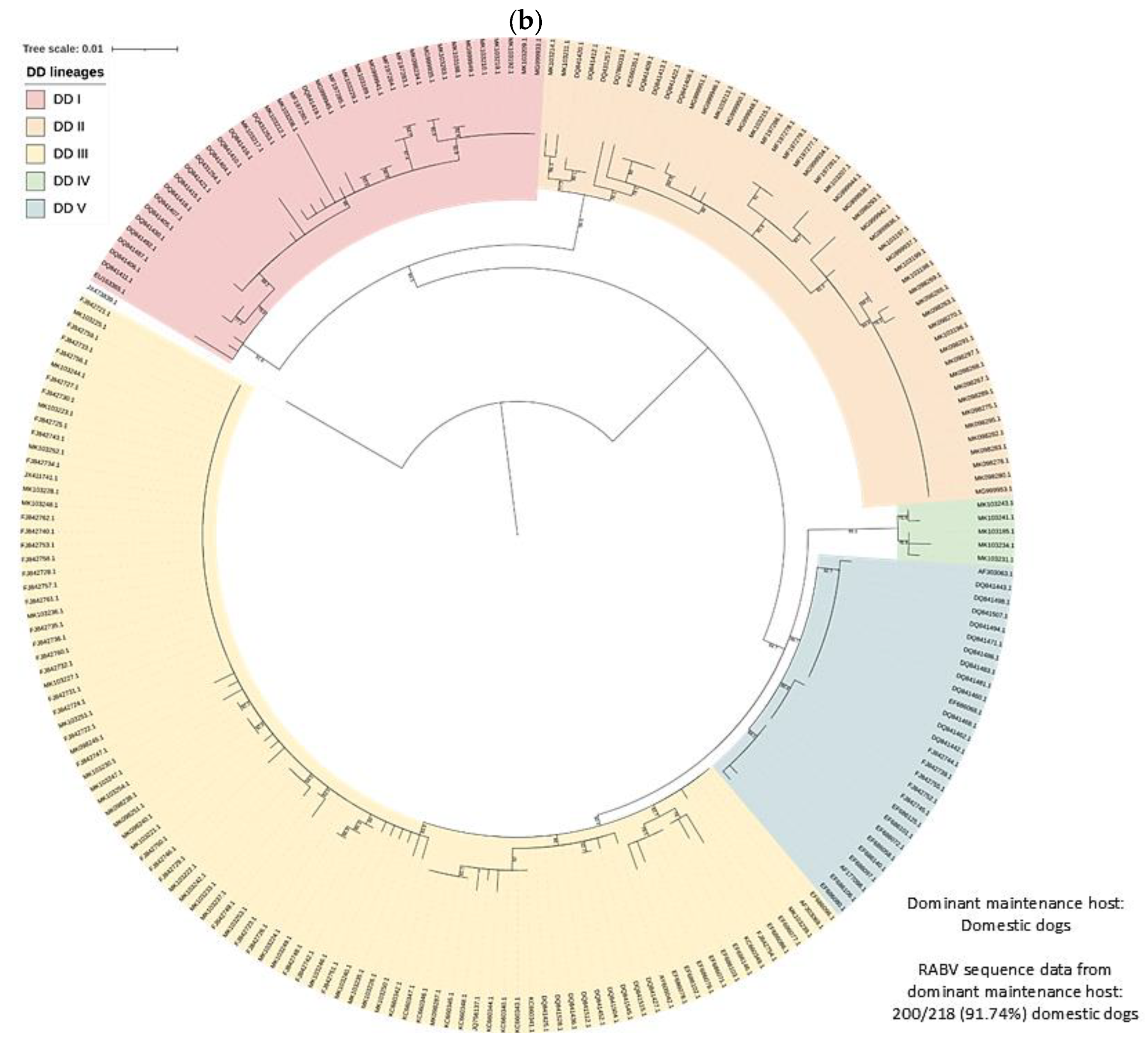
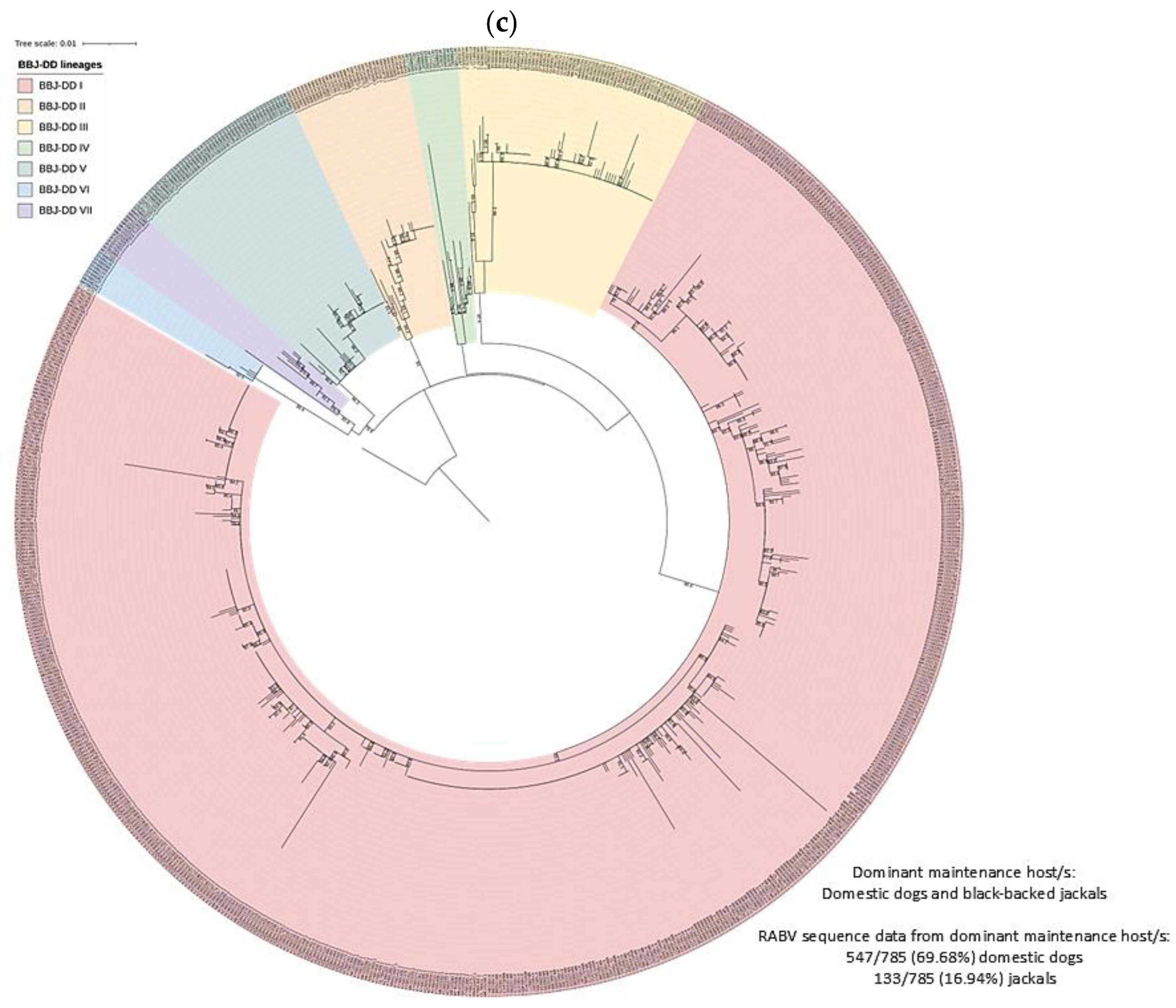
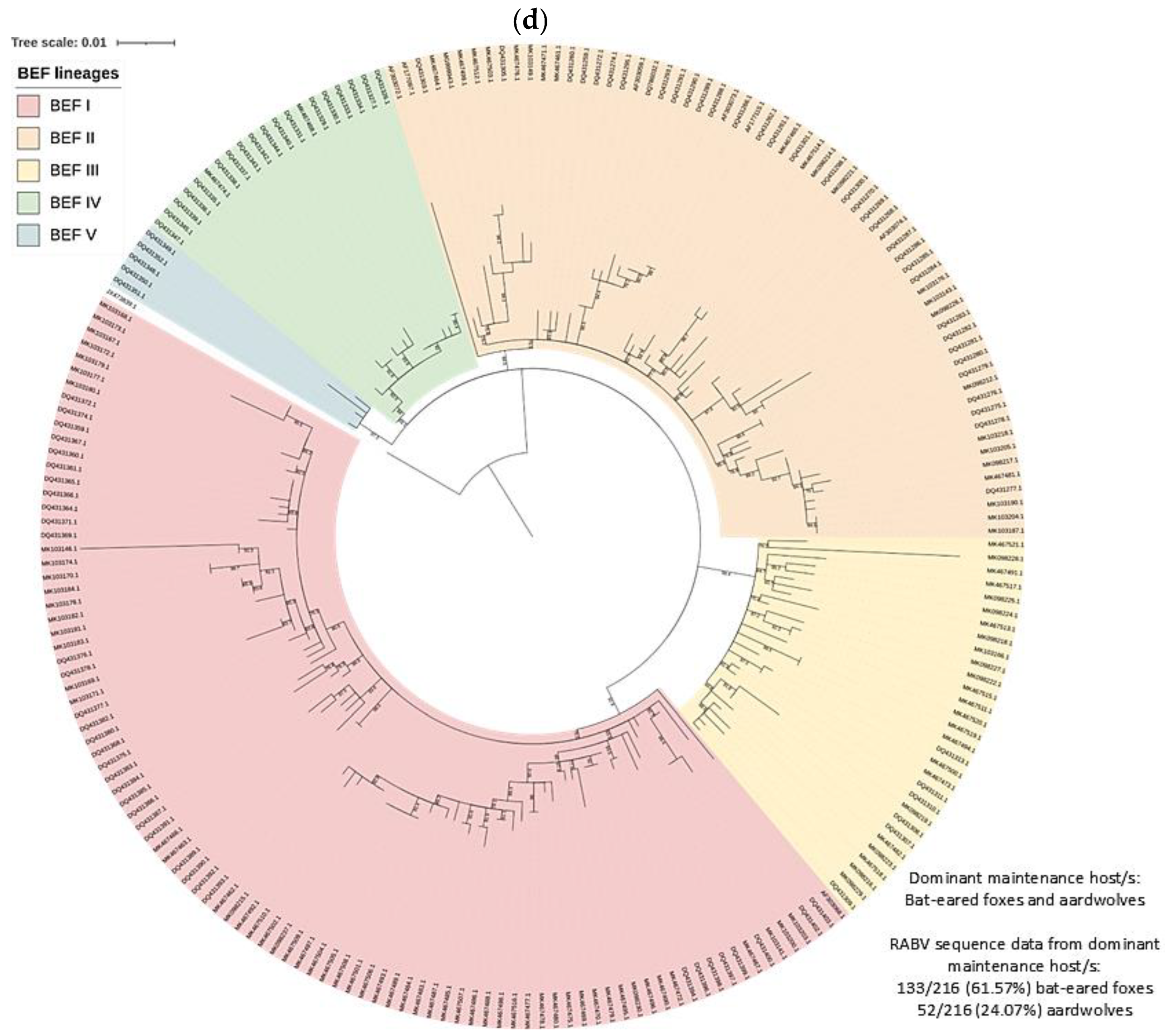
3.2. RABV Lineages and Distribution Based on Geospatial and Molecular Epidemiological Data
3.3. Rabies Control
4. Discussion
5. Limitations of This Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Minghui, R.; Stone, M.; Semedo, M.H.; Nel, L. New global strategic plan to eliminate dog-mediated rabies by 2030. Lancet Glob. Health 2018, 6, e828–e829. [Google Scholar] [CrossRef] [PubMed]
- Coleman, P.G.; Dye, C. Immunization coverage required to prevent outbreaks of dog rabies. Vaccine 1996, 14, 185–186. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, R. Rabies. In Infectious Diseases of Livestock with Special Reference to Southern Africa, 2nd ed.; Coetzer, J., Tustin, R., Eds.; Oxford University Press: Cape Town, South Africa, 2004; pp. 1123–1182. [Google Scholar]
- King, A.; Meredith, C.; Thomson, G. The biology of southern African lyssavirus variants. Curr. Top. Microbiol. Immunol. 1994, 187, 267–295. [Google Scholar] [PubMed]
- Von Teichman, B.; Thomson, G.; Meredith, C.; Nel, L. Molecular epidemiology of rabies virus in South Africa: Evidence for two distinct virus groups. J. Gen. Virol. 1995, 76, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Sabeta, C.T.; Bingham, J.; Nel, L.H. Molecular epidemiology of canid rabies in Zimbabwe and South Africa. Virus Res. 2003, 91, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Ngoepe, C.E.; Shumba, W.; Sabeta, C. Evidence for a host switching in the maintenance of canid rabies variant in two wild carnivore species in the Northern Cape Province, South Africa. J. S. Afr. Vet. Assoc. 2023, 94, 123–129. [Google Scholar] [CrossRef]
- Coetzee, P.; Nel, L.H. Emerging epidemic dog rabies in coastal South Africa: A molecular epidemiological analysis. Virus Res. 2007, 126, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Sabeta, C.T.; Mansfield, K.L.; McElhinney, L.M.; Fooks, A.R.; Nel, L.H. Molecular epidemiology of rabies in bat-eared foxes (Otocyon megalotis) in South Africa. Virus Res. 2007, 129, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zulu, G.C.; Sabeta, C.T.; Nel, L.H. Molecular epidemiology of rabies: Focus on domestic dogs (Canis familiaris) and black-backed jackals (Canis mesomelas) from northern South Africa. Virus Res. 2009, 140, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Sabeta, C.T.; Mkhize, G.C.; Ngoepe, E.C. An evaluation of dog rabies control in Limpopo province (South Africa). Epidemiol. Infect. 2011, 139, 1470–1475. [Google Scholar] [CrossRef]
- Nel, L.H.; Sabeta, C.T.; Von Teichman, B.; Jaftha, J.B.; Rupprecht, C.E.; Bingham, J. Mongoose rabies in southern Africa: A re-evaluation based on molecular epidemiology. Virus Res. 2005, 109, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Ngoepe, C.E.; Sabeta, C.; Nel, L. The spread of canine rabies into Free State province of South Africa: A molecular epidemiological characterization. Virus Res. 2009, 142, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Nel, L.; Jacobs, J.; Jaftha, J.; Meredith, C. Natural spillover of a distinctly canidae-associated biotype of rabies virus into an expanded wildlife host range in southern Africa. Virus Genes 1997, 15, 79–82. [Google Scholar] [CrossRef]
- Van Zyl, N.; Markotter, W.; Nel, L.H. Evolutionary history of African mongoose rabies. Virus Res. 2010, 150, 93–102. [Google Scholar] [CrossRef]
- Chaparro, F.; Esterhuysen, J. The role of the yellow mongoose (Cynictis penicillata) in the epidemiology of rabies in South Africa-preliminary results. Onderstepoort J. Vet. Res. 1993, 60, 373–377. [Google Scholar] [PubMed]
- Bingham, J. Canine rabies ecology in Southern Africa. Emerg. Infect. Dis. 2005, 11, 1337–1342. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, R.; Barnard, B.; Meredith, C.; Bishop, G.; Bruckner, G.; Foggin, C.; Hubschle, O. Rabies in southern Africa. Onderstepoort J. Vet. Res. 1993, 60, 325–346. [Google Scholar] [PubMed]
- Koeppel, K.N.; van Schalkwyk, O.L.; Thompson, P.N. Patterns of rabies cases in South Africa between 1993 and 2019, including the role of wildlife. Transbound. Emerg. Dis. 2022, 69, 836–848. [Google Scholar] [CrossRef] [PubMed]
- Malan, A.J.; Coetzer, A.; Bosch, C.; Wright, N.; Nel, L.H. A Perspective of the Epidemiology of Rabies in South Africa, 1998–2019. Trop. Med. Infect. Dis. 2024, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S.; Battistuzzi, F.U. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Towns, J.; Cockerill, T.; Dahan, M.; Foster, I.; Gaither, K.; Grimshaw, A.; Hazlewood, V.; Lathrop, S.; Lifka, D.; Peterson, G.D. XSEDE: Accelerating scientific discovery. Comput. Sci. Eng. 2014, 16, 62–74. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchene, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kuhnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- QGIS Development Team. QGIS Geographic Information System; Open Source Geospatial Foundation Project 2023. Available online: http://qgis.org (accessed on 13 April 2023).
- Centre for Emerging Zoonotic and Parasitic Diseases. Rabies. NICD Commun. Dis. Commun. 2022, 21, 3. [Google Scholar]
- Ravensberg, M.F.; Fanoy, E.B.; Whelan, J.; Embregts, C.W.; GeurtsvanKessel, C.H.; Strydom, J.B. Ongoing rabies outbreak in dogs of unprecedented scale and human cases in Nelson Mandela Bay Municipality, South Africa, up to 13 February 2022. Euro Surveill 2022, 27, 2200252. [Google Scholar] [CrossRef]
- Ngoepe, E.; Chirima, J.G.; Mohale, D.; Mogano, K.; Suzuki, T.; Makita, K.; Sabeta, C.T. Rabies outbreak in black-backed jackals (Canis mesomelas), South Africa, 2016. Epidemiol. Infect. 2022, 150, e137. [Google Scholar] [CrossRef] [PubMed]
- Mkhize, G.C.; Ngoepe, E.C.; Du Plessis, B.J.; Reininghaus, B.; Sabeta, C.T. Re-emergence of dog rabies in Mpumalanga province, South Africa. Vector Borne Zoonotic Dis. 2010, 10, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Malan, A.J.; Coetzer, A.; Sabeta, C.T.; Nel, L.H. Epidemiological Interface of Sylvatic and Dog Rabies in the North West Province of South Africa. Trop. Med. Infect. Dis. 2022, 7, 90. [Google Scholar] [CrossRef]
- Sabeta, C.T.; Weyer, J.; Geertsma, P.; Mohale, D.; Miyen, J.; Blumberg, L.H.; Leman, P.A.; Phahladira, B.; Shumba, W.; Walters, J.; et al. Emergence of rabies in the gauteng province, South Africa: 2010–2011. J. S. Afr. Vet. Assoc. 2013, 84, 2010–2011. [Google Scholar] [CrossRef]
- Coetzer, A. Chapter II: Epidemiology of an outbreak of rabies in wildlife species in the KwaZulu-Natal province of South Africa. In The Epidemiology of Rabies in Selected Sub-Saharan Countries; University of Pretoria: Pretoria, South Africa, 2018. [Google Scholar]
- Dodds, W.J.; Larson, L.J.; Christine, K.L.; Schultz, R.D. Duration of immunity after rabies vaccination in dogs: The Rabies Challenge Fund research study. Can. J. Vet. Res. 2020, 84, 153–158. [Google Scholar] [PubMed]
- Weyer, J.; Szmyd-Potapczuk, A.V.; Blumberg, L.H.; Leman, P.A.; Markotter, W.; Swanepoel, R.; Paweska, J.T.; Nel, L.H. Epidemiology of human rabies in South Africa, 1983–2007. Virus Res. 2011, 155, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Weyer, J.; Dermaux-Msimang, V.; Grobbelaar, A.; Le Roux, C.; Moolla, N.; Paweska, J.; Blumberg, L. Epidemiology of human rabies in South Africa, 2008–2018. S. Afr. Med. J. 2020, 110, 877–881. [Google Scholar] [CrossRef]
- LeRoux, K.; Stewart, D.; Perrett, K.D.; Nel, L.H.; Kessels, J.A.; Abela-Ridder, B. Rabies control in Kwazulu-Natal, South Africa. Bull. World Health Organ. 2018, 96, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Van Sittert, S.J.; Raath, J.; Akol, G.W.; Miyen, J.M.; Mlahlwa, B.; Sabeta, C.T. Rabies in the Eastern Cape Province of South Africa—Where are we going wrong? J. S. Afr. Vet. Assoc. 2010, 81, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Hergert, M.; Le Roux, K.; Nel, L.H. Characteristics of owned dogs in rabies endemic KwaZulu-Natal province, South Africa. BMC Vet. Res. 2018, 14, 278. [Google Scholar] [CrossRef]
- Hergert, M.; LeRoux, K.; Nel, L. Risk factors associated with nonvaccination rabies status of dogs in KwaZulu-Natal, South Africa. Vet. Med. Res. Rep. 2016, 7, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Koeppel, K.N.; Geertsma, P.; Kuhn, B.F.; van Schalkwyk, O.L.; Thompson, P.N. Antibody response to Raboral VR-G® oral rabies vaccine in captive and free-ranging black-backed jackals (Canis mesomelas). Onderstepoort J. Vet. Res. 2022, 89, a1975. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viljoen, N.; Sabeta, C.; Markotter, W.; Weyer, J. Temporal and Spatial Analysis of Rabies Virus Lineages in South Africa. Viruses 2025, 17, 340. https://doi.org/10.3390/v17030340
Viljoen N, Sabeta C, Markotter W, Weyer J. Temporal and Spatial Analysis of Rabies Virus Lineages in South Africa. Viruses. 2025; 17(3):340. https://doi.org/10.3390/v17030340
Chicago/Turabian StyleViljoen, Natalie, Claude Sabeta, Wanda Markotter, and Jacqueline Weyer. 2025. "Temporal and Spatial Analysis of Rabies Virus Lineages in South Africa" Viruses 17, no. 3: 340. https://doi.org/10.3390/v17030340
APA StyleViljoen, N., Sabeta, C., Markotter, W., & Weyer, J. (2025). Temporal and Spatial Analysis of Rabies Virus Lineages in South Africa. Viruses, 17(3), 340. https://doi.org/10.3390/v17030340