
Identification of Mycoviruses in Cytospora chrysosperma: Potential Biocontrol Agents for Walnut Canker

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains of C. chrysosperma
2.2. Metatranscriptomic Sequencing and Bioinformatics Analysis
2.3. Putative Mycovirus Confirmation
2.4. Growth Rate and Pathogenicity Test
2.5. Horizontal Transmission of Hypovirulence
3. Results
3.1. Identification of Walnut Canker Pathogens
3.2. Metatranscriptomic Identification of Mycoviruses Infecting the Tested Strains
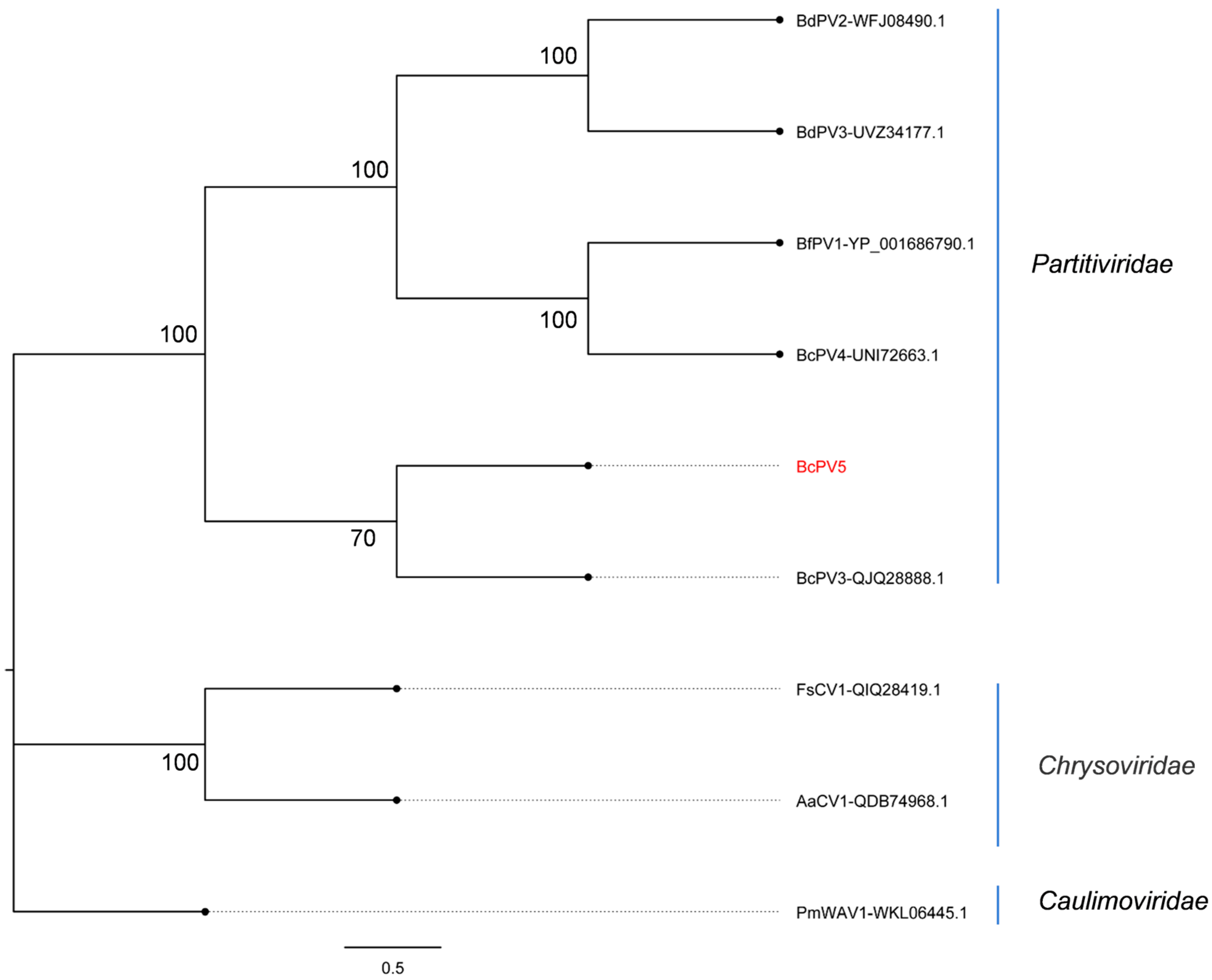
3.3. Phylogenetic Analysis
3.4. Detection of Mycoviruses and Their Impact on the Growth of C. chrysosperma
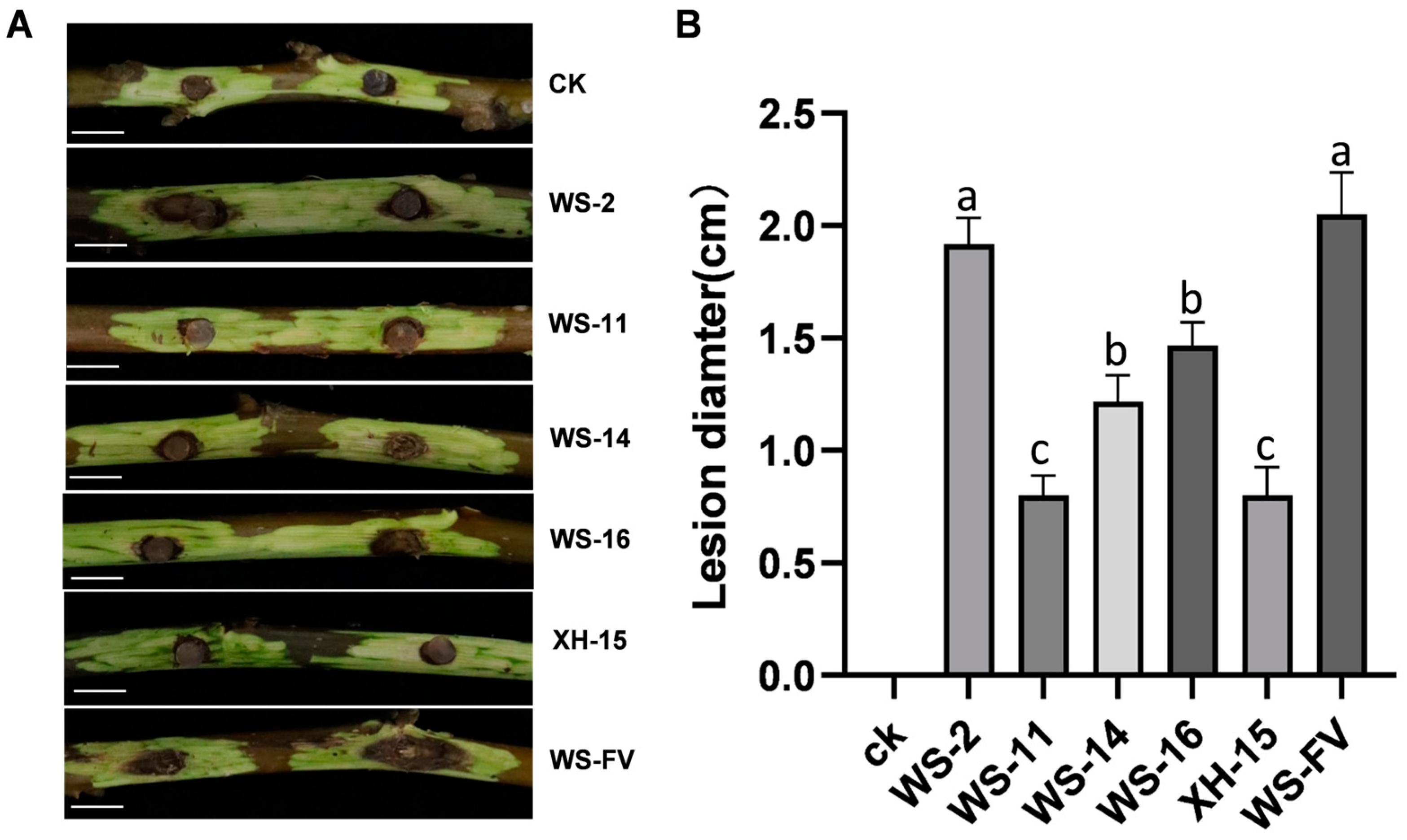
3.5. Virulence Determination of Strains Carrying Mycoviruses
3.6. Horizontal Transmission of Mycoviruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pollegioni, P.; Woeste, K.; Chiocchini, F.; Del Lungo, S.; Ciolfi, M.; Olimpieri, I.; Tortolano, V.; Clark, J.; Hemery, G.E.; Mapelli, S.; et al. Rethinking the history of common walnut (Juglans regia L.) in Europe: Its origins and human interactions. PLoS ONE 2017, 12, e0172541. [Google Scholar] [CrossRef]
- Bernard, A.; Barreneche, T.; Lheureux, F.; Dirlewanger, E. Analysis of genetic diversity and structure in a worldwide walnut (Juglans regia L.) germplasm using SSR markers. PLoS ONE 2018, 13, e0208021. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.; Sha, S.; Wang, L.; Feng, H.; Wang, Z.; Yan, C.; Zhang, Y. First report of Diplodia mutila causing canker and branch dieback of walnut trees in Xinjiang, China. Plant Dis. 2023, 107, 2545. [Google Scholar] [CrossRef]
- Fan, X.; Hyde, K.D.; Liu, M.; Liang, Y.; Tian, C. Cytospora species associated with walnut canker disease in China, with description of a new species C. gigalocus. Fungal Biol. 2015, 119, 310–319. [Google Scholar] [CrossRef]
- Sambaraju, K.R.; DesRochers, P.; Rioux, D. Factors influencing the regional dynamics of butternut canker. Plant Dis. 2018, 102, 743–752. [Google Scholar] [CrossRef]
- Bracalini, M.; Benigno, A.; Aglietti, C.; Panzavolta, T.; Moricca, S. Thousand cankers disease in walnut trees in Europe: Current status and management. Pathogens 2023, 12, 164. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, X.; Kotta-Loizou, I.; Dong, K.; Li, S.; Ni, D.; Hong, N.; Wang, G.; Xu, W. A mycovirus modulates the endophytic and pathogenic traits of a plant associated fungus. ISME J. 2021, 15, 1893–1906. [Google Scholar] [CrossRef]
- Myers, J.M.; James, T.Y. Mycoviruses. Curr. Biol. 2022, 32, R150–R155. [Google Scholar] [CrossRef]
- Wang, L.; He, H.; Wang, S.; Chen, X.; Qiu, D.; Kondo, H.; Guo, L. Evidence for a novel negative-stranded RNA mycovirus isolated from the plant pathogenic fungus Fusarium graminearum. Virology 2018, 518, 232–240. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, J.; Yang, B.; Wang, Q.; Zhou, J.; Yu, W. Molecular characterization of a debilitation-associated partitivirus infecting the pathogenic fungus Aspergillus flavus. Front. Microbiol. 2019, 10, 626. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Osaki, H.; Iwanami, T.; Matsumoto, N.; Ohtsu, Y. Molecular characterization of dsRNA segments 2 and 5 and electron microscopy of a novel reovirus from a hypovirulent isolate, W370, of the plant pathogen Rosellinia necatrix. J. Gen. Virol. 2003, 84, 2431–2437. [Google Scholar] [CrossRef]
- García-Pedrajas, M.D.; Cañizares, M.C.; Sarmiento-Villamil, J.L.; Jacquat, A.G.; Dambolena, J.S. Mycoviruses in biological control: Grom basic research to field implementation. Phytopathology 2019, 109, 1828–1839. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Kanematsu, S.; Suzuki, N. Viruses of the white root rot fungus, Rosellinia necatrix. Adv. Virus Res. 2013, 86, 177–214. [Google Scholar] [CrossRef]
- Kotta-Loizou, I. Mycoviruses and their role in fungal pathogenesis. Curr. Opin. Microbiol. 2021, 63, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, H.; Liao, X.; Gao, B.; Lu, X.; Sun, D.; Gong, W.; Zhong, J.; Zhu, H.; Pan, X.; et al. Mycoviral gene integration converts a plant pathogenic fungus into a biocontrol agent. Proc. Natl. Acad. Sci. USA 2022, 119, e2214096119. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Wu, J.; Jiang, D.; Xie, J.; Cheng, J.; Lin, Y. ORF Ι of mycovirus SsNSRV-1 is associated with debilitating symptoms of Sclerotinia sclerotiorum. Viruses 2020, 12, 456. [Google Scholar] [CrossRef]
- Zhang, H.; Xie, J.; Fu, Y.; Cheng, J.; Qu, Z.; Zhao, Z.; Cheng, S.; Chen, T.; Li, B.; Wang, Q.; et al. A 2-kb mycovirus converts a pathogenic fungus into a beneficial endophyte for Brassica protection and yield enhancement. Mol. Plant. 2020, 13, 1420–1433. [Google Scholar] [CrossRef]
- Hamid, M.R.; Xie, J.; Wu, S.; Maria, S.K.; Zheng, D.; Assane Hamidou, A.; Wang, Q.; Cheng, J.; Fu, Y.; Jiang, D. A novel deltaflexivirus that infects the plant fungal pathogen, Sclerotinia sclerotiorum, can be transmitted among host vegetative incompatible strains. Viruses 2018, 10, 295. [Google Scholar] [CrossRef]
- Melzer, M.S.; Ikeda, S.S.; Boland, G.J. Interspecific transmission of double-stranded RNA and hypovirulence from Sclerotinia sclerotiorum to S. minor. Phytopathology 2002, 92, 780–784. [Google Scholar] [CrossRef]
- Bian, R.; Andika, I.B.; Pang, T.; Lian, Z.; Wei, S.; Niu, E.; Wu, Y.; Kondo, H.; Liu, X.; Sun, L. Facilitative and synergistic interactions between fungal and plant viruses. Proc. Natl. Acad. Sci. USA 2020, 117, 3779–3788. [Google Scholar] [CrossRef]
- Wu, S.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Ghabrial, S.A.; Xie, J. Virus-mediated suppression of host non-self recognition facilitates horizontal transmission of heterologous viruses. PLoS Pathog. 2017, 13, e1006234. [Google Scholar] [CrossRef]
- Jia, H.; Dong, K.; Zhou, L.; Wang, G.; Hong, N.; Jiang, D.; Xu, W. A dsRNA virus with filamentous viral particles. Nat. Commun. 2017, 8, 168. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Qu, Z.; Mehmood, M.A.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D. Schizotrophic Sclerotinia sclerotiorum-mediated root and rhizosphere microbiome alterations activate growth and disease resistance in wheat. Microbiol. Spectr. 2023, 11, e0098123. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, D. Understanding the diversity, evolution, ecology, and applications of mycoviruses. Annu. Rev. Microbiol. 2024, 78, 595–620. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef]
- Sato, Y.; Hisano, S.; López-Herrera, C.J.; Kondo, H.; Suzuki, N. Three-layered complex interactions among capsidless (+) ssRNA yadokariviruses, dsRNA viruses, and a fungus. mBio 2022, 13, e0168522. [Google Scholar] [CrossRef]
- Eusebio-Cope, A.; Sun, L.; Tanaka, T.; Chiba, S.; Kasahara, S.; Suzuki, N. The chestnut blight fungus for studies on virus/host and virus/virus interactions: From a natural to a model host. Virology 2015, 477, 164–175. [Google Scholar] [CrossRef]
- Khan, H.A.; Nerva, L.; Bhatti, M.F. The good, the bad and the cryptic: The multifaceted roles of mycoviruses and their potential applications for a sustainable agriculture. Virology 2023, 585, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Fu, Y.; Guoqing, L.; Ghabrial, S.A. Viruses of the plant pathogenic fungus Sclerotinia sclerotiorum. Adv. Virus Res. 2013, 86, 215–248. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Hai, D.; Huang, H.; Cheng, J.; Fu, Y.; Lin, Y.; Jiang, D.; Xie, J. Mycoviromic analysis unveils complex virus composition in a hypovirulent strain of Sclerotinia sclerotiorum. J. Fungi. 2022, 8, 649. [Google Scholar] [CrossRef]
- Pearson, M.N.; Bailey, A.M. Viruses of Botrytis. Adv. Virus Res. 2013, 86, 249–272. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, S.; Ruan, S.; Nzabanita, C.; Wang, Y.; Guo, L. A mycovirus VIGS vector confers hypovirulence to a plant pathogenic fungus to control wheat FHB. Adv. Sci. 2023, 10, e2302606. [Google Scholar] [CrossRef]
- Hillman, B.I.; Annisa, A.; Suzuki, N. Viruses of plant-interacting fungi. Adv. Virus Res. 2018, 100, 99–116. [Google Scholar] [CrossRef]
- Hai, D.; Li, J.; Jiang, D.; Cheng, J.; Fu, Y.; Xiao, X.; Yin, H.; Lin, Y.; Chen, T.; Li, B.; et al. Plants interfere with non-self recognition of a phytopathogenic fungus via proline accumulation to facilitate mycovirus transmission. Nat. Commun. 2024, 15, 4748. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.H.; Dawe, A.L.; Churbanov, A.; Smith, M.L.; Milgroom, M.G.; Nuss, D.L. Molecular characterization of vegetative incompatibility genes that restrict hypovirus transmission in the chestnut blight fungus Cryphonectria parasitica. Genetics 2012, 190, 113–127. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Contig Number | Contig Length | Name of Putative Viruses | Best Match | aa Identity | Genome Type | Family/Genus |
|---|---|---|---|---|---|---|---|
| 1 | Acontig113 | 1456 bp | Botrytis cinerea partitivirus 5 BcPV5 | Botrytis cinerea partitivirus3 QJQ28888.1 | 44.72% | dsRNA | Partitiviridae |
| 2 | Acontig729 | 924 bp | Gammapartitivirus sp.-XJ1 GVsp-XJ1 | Gammapartitivirus sp. UDL14413.1 | 78.18% | dsRNA | Partitiviridae |
| 3 | Acontig372 | 2116 bp | Botoulivirus sp-XJ2 BVsp-XJ2 | Botoulivirus sp. UJQ92024.1 | 52.89% | +ssRNA | Botourmiaviridae |
| 4 | Acontig412 | 6806 bp | Luoyang Fusar tick virus 2 LfTV2 | Luoyang Fusar tick virus 1 UYL95319.1 | 50.82% | +ssRNA | Fusariviridae |
| 5 | Acontig397 | 2486 bp | Leptosphaeria biglobosa narnavirus 17 LbNV17 | Leptosphaeria biglobosa narnavirus 14 WKR37707.1 | 68.16% | +ssRNA | Narnaviridae |
| 6 | Acontig519 | 2566 bp | Sclerotinia sclerotiorum narnavirus 6 SsNV6 | Sclerotinia sclerotiorum narnavirus 5 UZC46344.1 | 70.60% | +ssRNA | Narnaviridae |
| 7 | Acontig526 | 3116 bp | Cytosporam ribis mitovirus 3 CrMV3 | Cytosporaribis mitovirus 1 AIS37555.1 | 64.67% | +ssRNA | Narnaviridae |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mi, Y.; Chen, S.; Liu, K.; Tie, Z.; Ren, J.; Zhang, M.; Liu, Z.; Zhao, S.; Xi, H.; Zhang, X. Identification of Mycoviruses in Cytospora chrysosperma: Potential Biocontrol Agents for Walnut Canker. Viruses 2025, 17, 180. https://doi.org/10.3390/v17020180
Mi Y, Chen S, Liu K, Tie Z, Ren J, Zhang M, Liu Z, Zhao S, Xi H, Zhang X. Identification of Mycoviruses in Cytospora chrysosperma: Potential Biocontrol Agents for Walnut Canker. Viruses. 2025; 17(2):180. https://doi.org/10.3390/v17020180
Chicago/Turabian StyleMi, Yingjie, Shaohua Chen, Kexin Liu, Zhanjiang Tie, Junchao Ren, Mingli Zhang, Zheng Liu, Sifeng Zhao, Hui Xi, and Xuekun Zhang. 2025. "Identification of Mycoviruses in Cytospora chrysosperma: Potential Biocontrol Agents for Walnut Canker" Viruses 17, no. 2: 180. https://doi.org/10.3390/v17020180
APA StyleMi, Y., Chen, S., Liu, K., Tie, Z., Ren, J., Zhang, M., Liu, Z., Zhao, S., Xi, H., & Zhang, X. (2025). Identification of Mycoviruses in Cytospora chrysosperma: Potential Biocontrol Agents for Walnut Canker. Viruses, 17(2), 180. https://doi.org/10.3390/v17020180