
Primordial Biochemicals Within Coacervate-Like Droplets and the Origins of Life



{kind=link}
Abstract
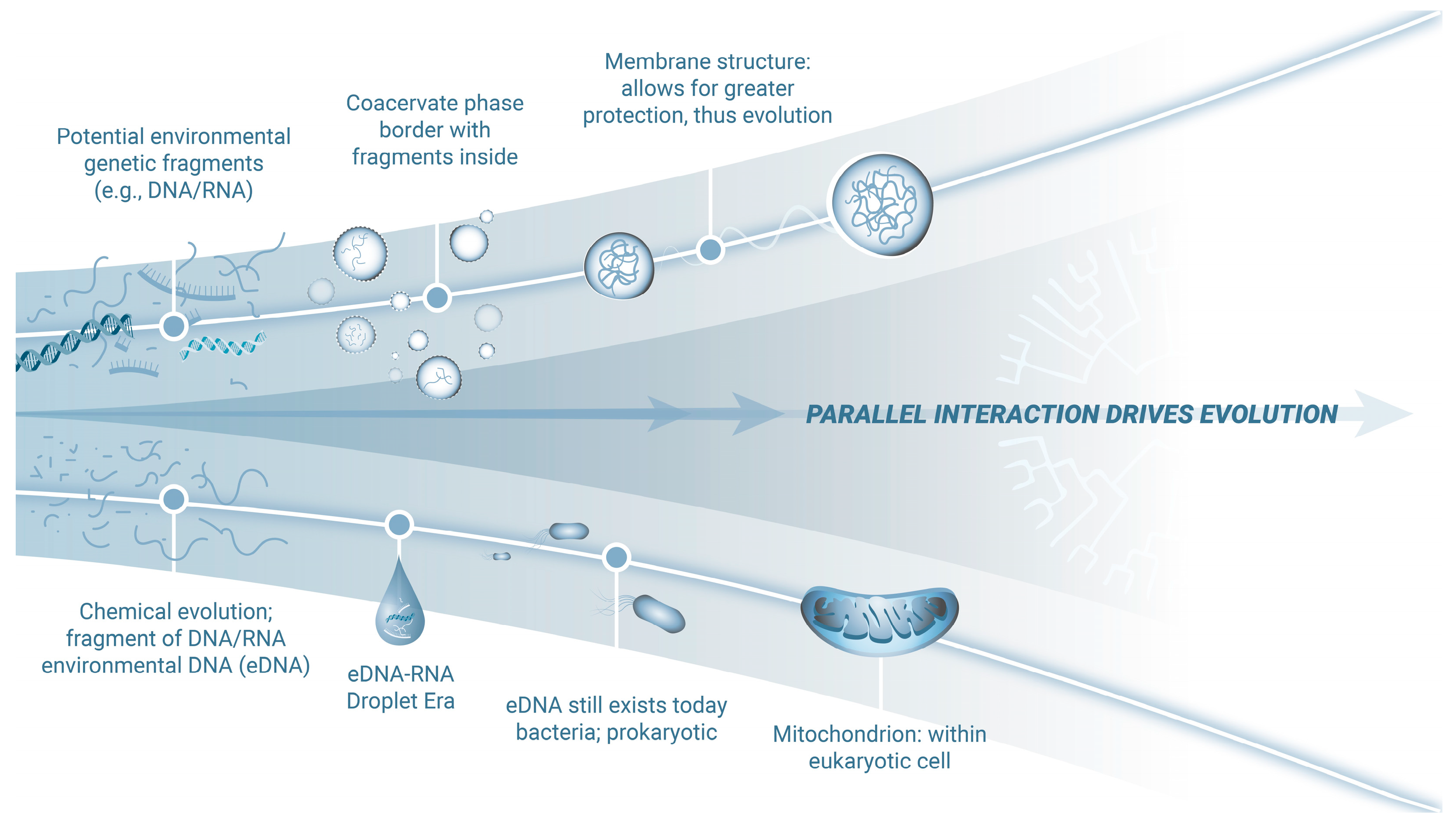
1. Environmental DNA and the Origins of Life
2. Environmental DNA and the Emergence of Viruses
3. Environmental DNA and the Evolution of Bacteria
4. Facilitating Genetic Evolution: The Droplet Era
5. Evolution of Chemical Communication
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stefano, G.B.; Kream, R.M. Prebiotic Formation of Protoalkaloids within Alkaline Oceanic Hydrothermal Vents in the Hadean Seafloor as a Prerequisite for Evolutionary Biodiversity. Med. Sci. Monit. 2020, 26, e928415. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, L.; McCauley, M.; Farrell, J.A.; Stammnitz, M.R.; Koda, S.A.; Mashkour, N.; Summers, V.; Osborne, T.; Whilde, J.; Duffy, D.J. Inadvertent human genomic bycatch and intentional capture raise beneficial applications and ethical concerns with environmental DNA. Nat. Ecol. Evol. 2023, 7, 873–888. [Google Scholar] [CrossRef]
- Duffy, D.J.; Martindale, M.Q. Perspectives on the expansion of human precision oncology and genomic approaches to sea turtle fibropapillomatosis. Commun. Biol. 2019, 2, 54. [Google Scholar] [CrossRef] [PubMed]
- Duffy, D.J.; Burkhalter, B. When is a lab animal not a lab animal? Lab. Anim. 2020, 49, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Wessner, D.R. The Origin of Viruses. Nat. Educ. 2010, 3, 37–39. [Google Scholar]
- Koonin, E.V.; Martin, W. On the origin of genomes and cells within inorganic compartments. Trends Genet. 2005, 21, 647–654. [Google Scholar] [CrossRef]
- Iyer, L.M.; Koonin, E.V.; Leipe, D.D.; Aravind, L. Origin and evolution of the archaeo-eukaryotic primase superfamily and related palm-domain proteins: Structural insights and new members. Nucleic Acids Res. 2005, 33, 3875–3896. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Halami, P.M. Presence of extracellular DNA & protein in biofilm formation by gentamicin-resistant Lactobacillus plantarum. Indian J. Med. Res. 2019, 149, 257–262. [Google Scholar] [CrossRef]
- Yang, K.; Wang, L.; Cao, X.; Gu, Z.; Zhao, G.; Ran, M.; Yan, Y.; Yan, J.; Xu, L.; Gao, C.; et al. The Origin, Function, Distribution, Quantification, and Research Advances of Extracellular DNA. Int. J. Mol. Sci. 2022, 23, 3690. [Google Scholar] [CrossRef]
- Atlante, A.; Valenti, D. Mitochondria Have Made a Long Evolutionary Path from Ancient Bacteria Immigrants within Eukaryotic Cells to Essential Cellular Hosts and Key Players in Human Health and Disease. Curr. Issues Mol. Biol. 2023, 45, 4451–4479. [Google Scholar] [CrossRef]
- Stefano, G.B.; Kream, R.M. Viruses Broaden the Definition of Life by Genomic Incorporation of Artificial Intelligence and Machine Learning Processes. Curr. Neuropharmacol. 2022, 20, 1888–1893. [Google Scholar] [CrossRef]
- Margulis, L.; Bermudes, D. Symbiosis as a mechanism of evolution: Status of cell symbiosis theory. Symbiosis 1985, 1, 101–124. [Google Scholar]
- Cutler, R.G. Evolutionary Biology of Senescence; Behnke, J.A., Finch, C.E., Moment, G.B., Eds.; Plenum Press: New York, NY, USA, 1978; 388p. [Google Scholar]
- Stefano, G.; Büttiker, P.; Weissenberger, S.; Esch, T.; Anders, M.; Raboch, J.; Kream, R.; Ptacek, R. Independent and sensory human mitochondrial functions reflecting symbiotic evolution. Front. Cell. Infect. Microbiol. 2023, 13, 1130197. [Google Scholar] [CrossRef] [PubMed]
- Stefano, G.B.; Kream, R.M. Mitochondrial DNA Heteroplasmy as an Informational Reservoir Dynamically Linked to Metabolic and Immunological Processes Associated with COVID-19 Neurological Disorders. Cell. Mol. Neurobiol. 2022, 42, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Bandyopadhaya, A.; Singh, V.K.; Kovacic, F.; Cha, S.; Oldham, W.M.; Tzika, A.A.; Rahme, L.G. The bacterial quorum sensing signal 2’-aminoacetophenone rewires immune cell bioenergetics through the Ppargc1a/Esrra axis to mediate tolerance to infection. eLife 2024, 13, RP97568. [Google Scholar] [CrossRef] [PubMed]
- Maurice, N.M.; Sadikot, R.T. Mitochondrial Dysfunction in Bacterial Infections. Pathogens 2023, 12, 1005. [Google Scholar] [CrossRef] [PubMed]
- Stefano, G.B. Antibiotics and Antiviral Agents Can Trigger Mitochondrial Dysfunction that Leads to Psychiatric Disorders. Mind Bull. Mind-Body Med. Res. 2023, 2, 8–10. [Google Scholar] [CrossRef]
- Stefano, G.B.; Samuel, J.; Kream, R.M. Antibiotics May Trigger Mitochondrial Dysfunction Inducing Psychiatric Disorders. Med. Sci. Monit. 2017, 23, 101–106. [Google Scholar] [CrossRef]
- Warstler, A.; Bean, J. Antimicrobial-Induced Cognitive Side Effects. Ment. Health Clin. 2016, 6, 207–214. [Google Scholar] [CrossRef]
- Jenkins, H.L.; Graham, R.; Porter, J.S.; Vieira, L.M.; De Almeida, A.C.S.; Hall, A.; O’Dea, A.; Coppard, S.E.; Waeschenbach, A. Unprecedented Frequency of Mitochondrial Introns in Colonial Bilaterians. Sci. Rep. 2022, 12, 10889. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Schon, K.R.; Elgar, G.; Orioli, A.; Tanguy, M.; Giess, A.; Tischkowitz, M.; Caulfield, M.J.; Chinnery, P.F. Nuclear-Embedded Mitochondrial DNA Sequences in 66,083 Human Genomes. Nature 2022, 611, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Karan, K.R.; Gu, W.; Klein, H.-U.; Sturm, G.; De Jager, P.L.; Bennett, D.A.; Hirano, M.; Picard, M.; Mills, R.E. Somatic Nuclear Mitochondrial DNA Insertions Are Prevalent in the Human Brain and Accumulate over Time in Fibroblasts. PLoS Biol. 2024, 22, e3002723. [Google Scholar] [CrossRef]
- van Swaay, D.; Tang, T.Y.; Mann, S.; de Mello, A. Microfluidic Formation of Membrane-Free Aqueous Coacervate Droplets in Water. Angew. Chem. Int. Ed. Engl. 2015, 54, 8398–8401. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Douglas, J.F.; Tirrell, M.; Karim, A. Manipulation of coacervate droplets with an electric field. Proc. Natl. Acad. Sci. USA 2022, 119, e2203483119. [Google Scholar] [CrossRef]
- Moulik, S.P.; Rakshit, A.K.; Pan, A.; Naskar, B. An Overview of Coacervates: The Special Disperse State of Amphiphilic and Polymeric Materials in Solution. Colloids Interfaces 2022, 6, 45. [Google Scholar] [CrossRef]
- Stefano, G.B. (Ed.) Conformational matching: A possible evolutionary force in the evolvement of signal systems. In CRC Handbook of Comparative Opioid and Related Neuropeptide Mechanisms; CRC Press Inc.: Boca Raton, FL, USA, 1986; Volume 2, pp. 271–277. [Google Scholar]
- Stefano, G.B. The evolvement of signal systems: Conformational matching a determining force stabilizing families of signal molecules. Comp. Biochem. Physiol. C 1988, 90, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Büttiker, P.; Boukherissa, A.; Weissenberger, S.; Ptacek, R.; Anders, M.; Raboch, J.; Stefano, G.B. Cognitive Impact of Neurotropic Pathogens: Investigating Molecular Mimicry through Computational Methods. Cell. Mol. Neurobiol. 2024, 44, 72. [Google Scholar] [CrossRef] [PubMed]
- Stefano, G.B.; Büttiker, P.; Weissenberger, S.; Anders, M.; Raboch, J.; Kream, R.M. Viruses May Be Redefined as Self-Replicating Entities: Expanding the Definition of Life. Mind-Bull. Mind-Body Med. Res. 2024, 3, 2–8. [Google Scholar] [CrossRef]
- Darwin, C. Origin of the Species; The Easton Press: Norwalk, CT, USA, 1976. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefano, G.B.; Kream, R.M. Primordial Biochemicals Within Coacervate-Like Droplets and the Origins of Life. Viruses 2025, 17, 146. https://doi.org/10.3390/v17020146
Stefano GB, Kream RM. Primordial Biochemicals Within Coacervate-Like Droplets and the Origins of Life. Viruses. 2025; 17(2):146. https://doi.org/10.3390/v17020146
Chicago/Turabian StyleStefano, George B., and Richard M. Kream. 2025. "Primordial Biochemicals Within Coacervate-Like Droplets and the Origins of Life" Viruses 17, no. 2: 146. https://doi.org/10.3390/v17020146
APA StyleStefano, G. B., & Kream, R. M. (2025). Primordial Biochemicals Within Coacervate-Like Droplets and the Origins of Life. Viruses, 17(2), 146. https://doi.org/10.3390/v17020146