
Strain-Dependent Assessment of Powassan Virus Transmission to Ixodes scapularis Ticks
 ,
,
Abstract
1. Introduction
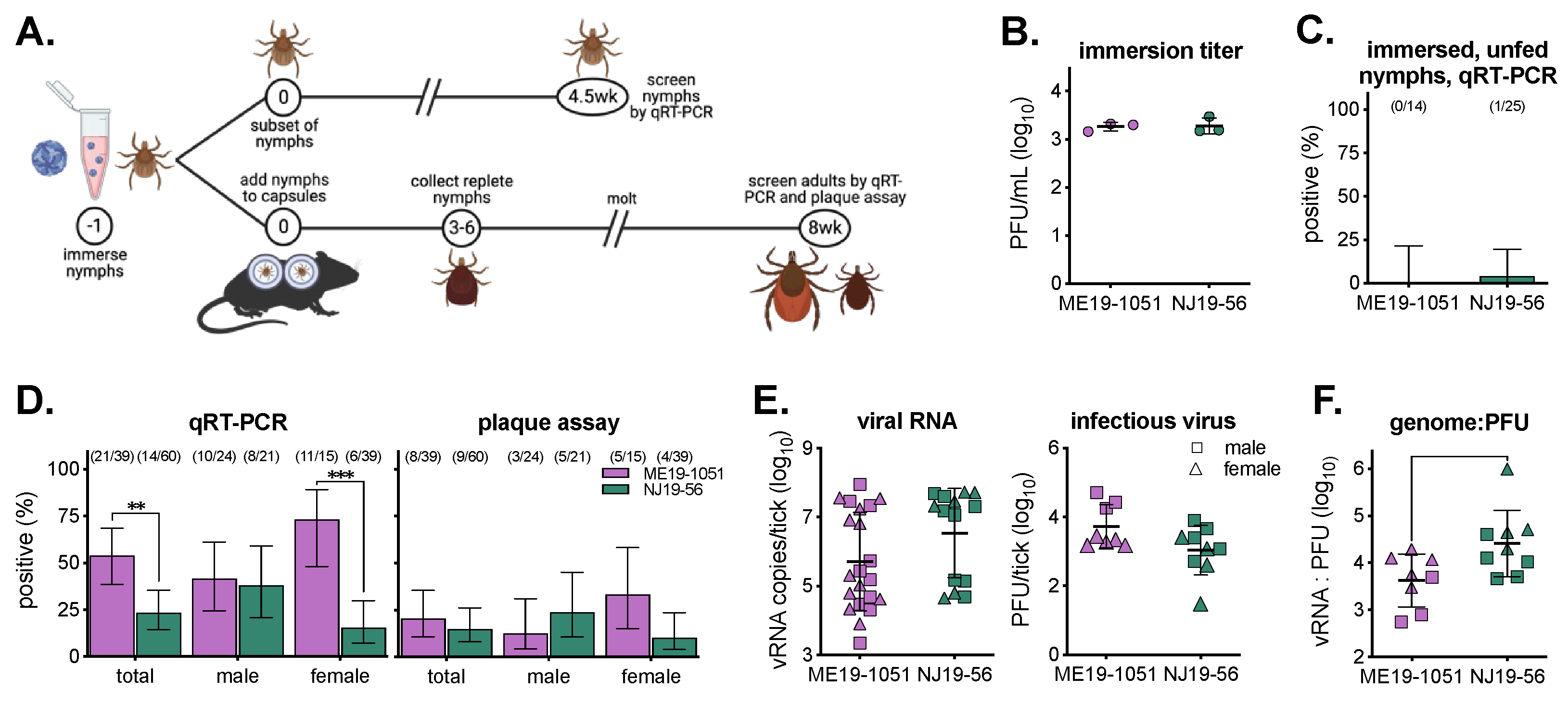
2. Materials and Methods
3. Results
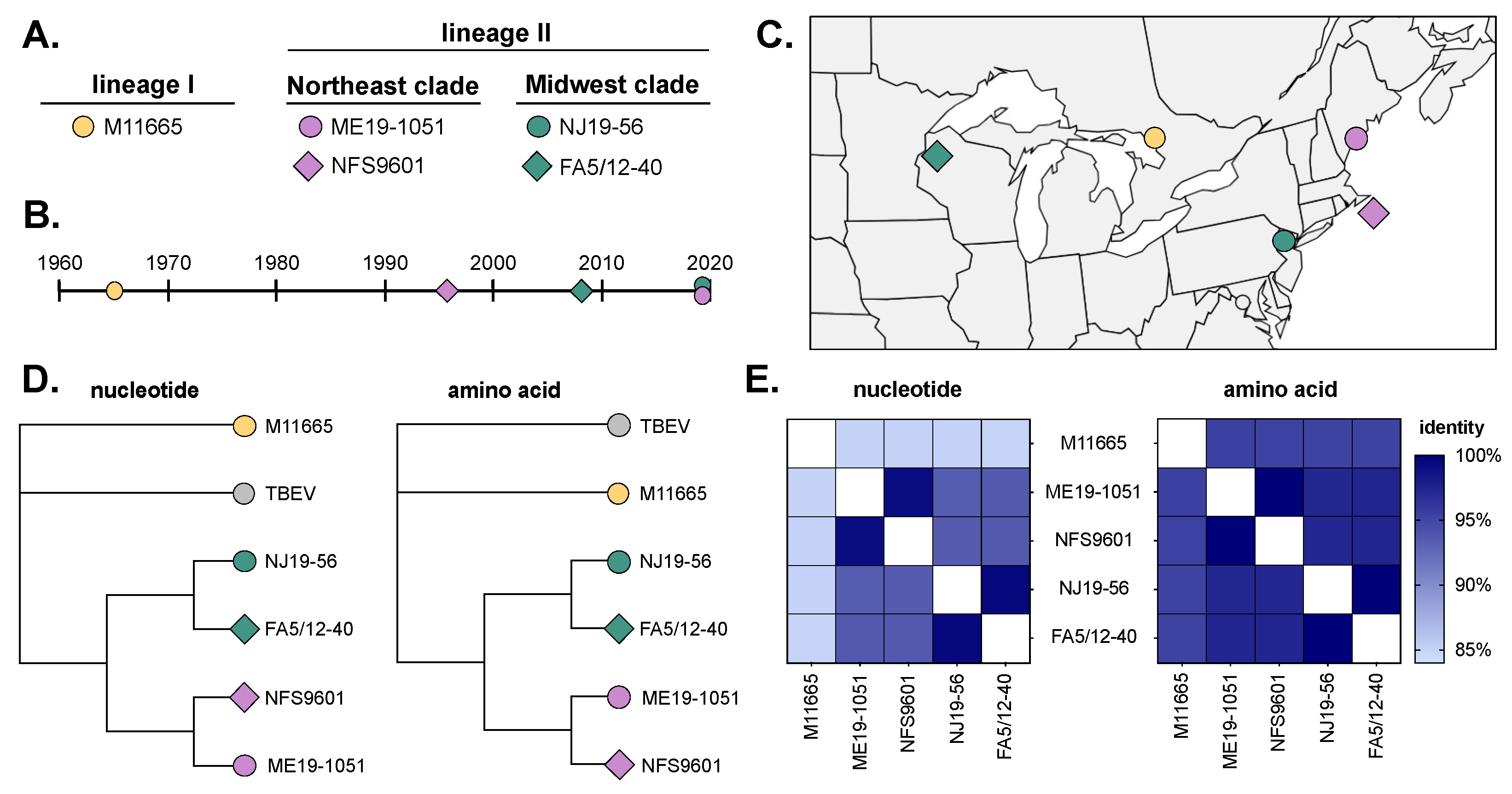
3.1. Virus Isolates Used in This Study
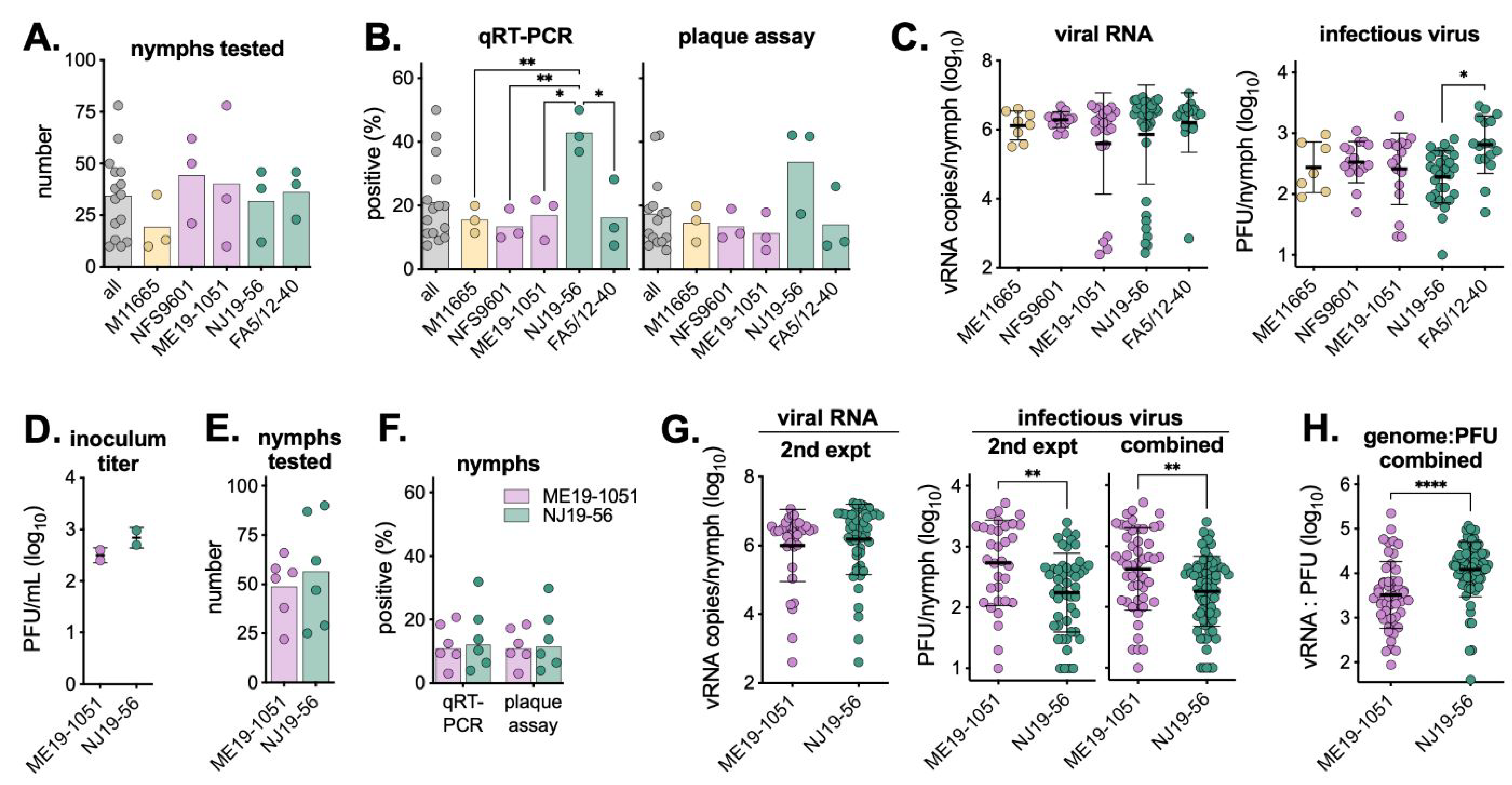
3.2. Viremic Transmission to Ticks
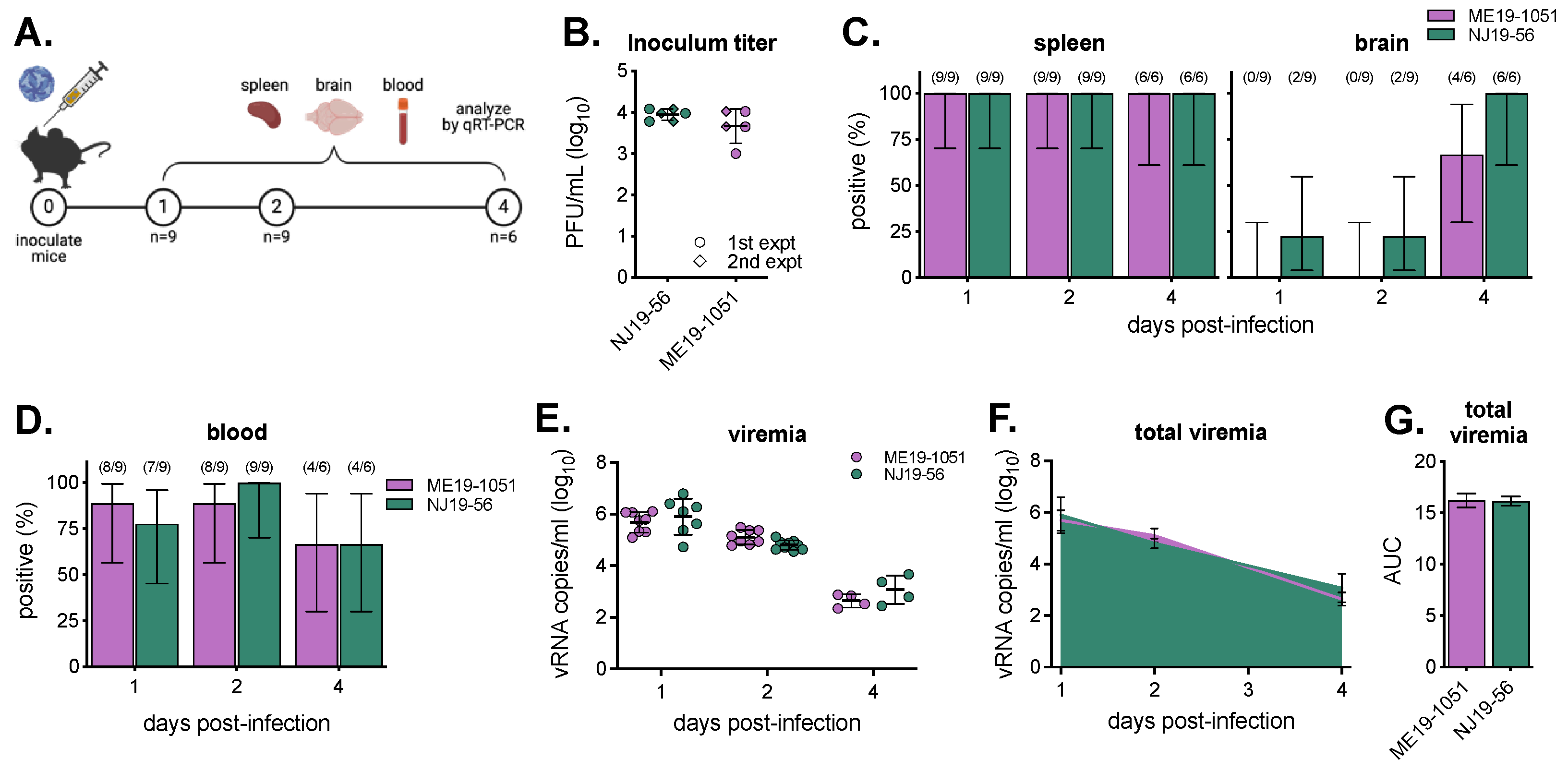
3.3. Viremia in BALB/c Mice
3.4. Artificial Infection of Ticks
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dennis, D.T.; Nekomoto, T.S.; Victor, J.C.; Paul, W.S.; Piesman, J. Reported distribution of Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae) in the United States. J. Med. Entomol. 1998, 35, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Eisen, L. The Blacklegged Tick, Ixodes scapularis: An Increasing Public Health Concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Sonenshine, D.E. Range Expansion of Tick Disease Vectors in North America: Implications for Spread of Tick-Borne Disease. Int. J. Environ. Res. Public Health 2018, 15, 478. [Google Scholar] [CrossRef] [PubMed]
- Ebel, G.D. Update on Powassan virus: Emergence of a North American tick-borne flavivirus. Annu. Rev. Entomol. 2010, 55, 95–110. [Google Scholar] [CrossRef]
- Powassan Virus. 2023. Available online: https://www.cdc.gov/powassan/data-maps/current-year-data.html (accessed on 14 March 2023).
- Mc, L.D.; Donohue, W.L. Powassan virus: Isolation of virus from a fatal case of encephalitis. Can. Med. Assoc. J. 1959, 80, 708–711. [Google Scholar]
- Campbell, O.; Krause, P.J. The emergence of human Powassan virus infection in North America. Ticks Tick. Borne Dis. 2020, 11, 101540. [Google Scholar] [CrossRef] [PubMed]
- Telford, S.R., 3rd; Armstrong, P.M.; Katavolos, P.; Foppa, I.; Garcia, A.S.; Wilson, M.L.; Spielman, A. A new tick-borne encephalitis-like virus infecting New England deer ticks, Ixodes dammini. Emerg. Infect. Dis. 1997, 3, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Nofchissey, R.A.; Deardorff, E.R.; Blevins, T.M.; Anishchenko, M.; Bosco-Lauth, A.; Berl, E.; Lubelczyk, C.; Mutebi, J.P.; Brault, A.C.; Ebel, G.D.; et al. Seroprevalence of Powassan virus in New England deer, 1979–2010. Am. J. Trop. Med. Hyg. 2013, 88, 1159–1162. [Google Scholar] [CrossRef]
- Aliota, M.T.; Dupuis, A.P., 2nd; Wilczek, M.P.; Peters, R.J.; Ostfeld, R.S.; Kramer, L.D. The prevalence of zoonotic tick-borne pathogens in Ixodes scapularis collected in the Hudson Valley, New York State. Vector Borne Zoonotic Dis. 2014, 14, 245–250. [Google Scholar] [CrossRef]
- Klinepeter, K. Powassan Virus Identified in Ticks; Pennsylvania Department of Health: Harrisburg, PA, USA, 2022.
- Smalley, R.t.; Zafar, H.; Land, J.; Samour, A.; Hance, D.; Brennan, R.E. Detection of Borrelia miyamotoi and Powassan Virus Lineage II (Deer Tick Virus) from Odocoileus virginianus Harvested Ixodes scapularis in Oklahoma. Vector Borne Zoonotic Dis. 2022, 22, 209–216. [Google Scholar] [CrossRef]
- Cumbie, A.N.; Whitlow, A.M.; Eastwood, G. First Evidence of Powassan Virus (Flaviviridae) in Ixodes scapularis in Appalachian Virginia, USA. Am. J. Trop. Med. Hyg. 2021, 106, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Whitlow, A.M.; Cumbie, A.N.; Eastwood, G. Pathogen prevalence in Amblyomma americanum and Ixodes scapularis ticks from central Appalachian Virginia, U.S.A. J. Vector Ecol. 2022, 47, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Vogels, C.B.F.; Brackney, D.E.; Dupuis, A.P., 2nd; Robich, R.M.; Fauver, J.R.; Brito, A.F.; Williams, S.C.; Anderson, J.F.; Lubelczyk, C.B.; Lange, R.E.; et al. Phylogeographic reconstruction of the emergence and spread of Powassan virus in the northeastern United States. Proc. Natl. Acad. Sci. USA 2023, 120, e2218012120. [Google Scholar] [CrossRef] [PubMed]
- McMinn, R.J.; Langsjoen, R.M.; Bombin, A.; Robich, R.M.; Ojeda, E.; Normandin, E.; Goethert, H.K.; Lubelczyk, C.B.; Schneider, E.; Cosenza, D.; et al. Phylodynamics of deer tick virus in North America. Virus Evol. 2023, 9, vead008. [Google Scholar] [CrossRef]
- Brackney, D.E.; Vogels, C.B.F. The known unknowns of Powassan virus ecology. J. Med. Entomol. 2023, 60, 1142–1148. [Google Scholar] [CrossRef]
- Wood, C.L.; Lafferty, K.D. Biodiversity and disease: A synthesis of ecological perspectives on Lyme disease transmission. Trends Ecol. Evol. 2013, 28, 239–247. [Google Scholar] [CrossRef]
- Kwasnik, M.; Rola, J.; Rozek, W. Tick-Borne Encephalitis-Review of the Current Status. J. Clin. Med. 2023, 12, 6603. [Google Scholar] [CrossRef]
- Brackney, D.E.; Pesko, K.N.; Brown, I.K.; Deardorff, E.R.; Kawatachi, J.; Ebel, G.D. West Nile virus genetic diversity is maintained during transmission by Culex pipiens quinquefasciatus mosquitoes. PLoS ONE 2011, 6, e24466. [Google Scholar] [CrossRef] [PubMed]
- Migne, C.V.; Honig, V.; Bonnet, S.I.; Palus, M.; Rakotobe, S.; Galon, C.; Heckmann, A.; Vyletova, E.; Devillers, E.; Attoui, H.; et al. Evaluation of two artificial infection methods of live ticks as tools for studying interactions between tick-borne viruses and their tick vectors. Sci. Rep. 2022, 12, 491. [Google Scholar] [CrossRef]
- Mateos-Hernandez, L.; Rakotobe, S.; Defaye, B.; Cabezas-Cruz, A.; Simo, L. A Capsule-Based Model for Immature Hard Tick Stages Infestation on Laboratory Mice. J. Vis. Exp. 2020, 161, e61430. [Google Scholar] [CrossRef]
- Chernesky, M.A.; McLean, D.M. Localization of Powassan virus in Dermacentor andersoni ticks by immunofluorescence. Can. J. Microbiol. 1969, 15, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Cozens, D.W.; Armstrong, P.M.; Brackney, D.E. Vector competence of human-biting ticks Ixodes scapularis, Amblyomma americanum and Dermacentor variabilis for Powassan virus. Parasites Vectors 2021, 14, 466. [Google Scholar] [CrossRef] [PubMed]
- Raney, W.R.; Herslebs, E.J.; Langohr, I.M.; Stone, M.C.; Hermance, M.E. Horizontal and Vertical Transmission of Powassan Virus by the Invasive Asian Longhorned Tick, Haemaphysalis longicornis, Under Laboratory Conditions. Front. Cell Infect. Microbiol. 2022, 12, 923914. [Google Scholar] [CrossRef] [PubMed]
- Costero, A.; Grayson, M.A. Experimental transmission of Powassan virus (Flaviviridae) by Ixodes scapularis ticks (Acari:Ixodidae). Am. J. Trop. Med. Hyg. 1996, 55, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Bakker, J.W.; Esser, H.J.; Sprong, H.; Godeke, G.J.; Hoornweg, T.E.; de Boer, W.F.; Pijlman, G.P.; Koenraadt, C.J.M. Differential susceptibility of geographically distinct Ixodes ricinus populations to tick-borne encephalitis virus and louping ill virus. Emerg. Microbes Infect. 2024, 13, 2321992. [Google Scholar] [CrossRef]
- Godsey, M.S.; Savage, H.M.; Burkhalter, K.L.; Bosco-Lauth, A.M.; Delorey, M.J. Transmission of Heartland Virus (Bunyaviridae: Phlebovirus) by Experimentally Infected Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 2016, 53, 1226–1233. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, M.; Sajid, M.S.; Saqib, M.; Anjum, F.R.; Tayyab, M.H.; Rizwan, H.M.; Rashid, M.I.; Rashid, I.; Iqbal, A.; Siddique, R.M.; et al. Potential Mechanisms of Transmission of Tick-Borne Viruses at the Virus-Tick Interface. Front. Microbiol. 2022, 13, 846884. [Google Scholar] [CrossRef] [PubMed]
- Brackney, D.E.; Armstrong, P.M. Transmission and evolution of tick-borne viruses. Curr. Opin. Virol. 2016, 21, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, P.A.; Jones, L.D.; Labuda, M.; Kaufman, W.R. Adaptations of arboviruses to ticks. J. Med. Entomol. 1994, 31, 1–9. [Google Scholar] [CrossRef]
- Sonenshine, D.E.; Roe, R.M. Biology of Ticks, 2nd ed.; Oxford University Press: New York, NY, USA, 2014. [Google Scholar]
- Lejal, E.; Moutailler, S.; Simo, L.; Vayssier-Taussat, M.; Pollet, T. Tick-borne pathogen detection in midgut and salivary glands of adult Ixodes ricinus. Parasites Vectors 2019, 12, 152. [Google Scholar] [CrossRef]
- Narasimhan, S.; Rajeevan, N.; Liu, L.; Zhao, Y.O.; Heisig, J.; Pan, J.; Eppler-Epstein, R.; Deponte, K.; Fish, D.; Fikrig, E. Gut microbiota of the tick vector Ixodes scapularis modulate colonization of the Lyme disease spirochete. Cell Host Microbe 2014, 15, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Ochwoto, M.; Offerdahl, D.K.; Leung, J.M.; Schwartz, C.L.; Long, D.; Rosenke, R.; Stewart, P.E.; Saturday, G.A.; Bloom, M.E. Cytoarchitecture of ex vivo midgut cultures of unfed Ixodes scapularis infected with a tick-borne flavivirus. Ticks Tick. Borne Dis. 2024, 15, 102301. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.R.; Brunner, J.L.; Scoles, G.A.; Owen, J.P. Factors affecting larval tick feeding success: Host, density and time. Parasites Vectors 2015, 8, 340. [Google Scholar] [CrossRef] [PubMed]
- Grubaugh, N.D.; Ruckert, C.; Armstrong, P.M.; Bransfield, A.; Anderson, J.F.; Ebel, G.D.; Brackney, D.E. Transmission bottlenecks and RNAi collectively influence tick-borne flavivirus evolution. Virus Evol. 2016, 2, vew033. [Google Scholar] [CrossRef] [PubMed]
- Klasse, P.J. Molecular determinants of the ratio of inert to infectious virus particles. Prog. Mol. Biol. Transl. Sci. 2015, 129, 285–326. [Google Scholar] [CrossRef]
- Sanjuan, R. Collective properties of viral infectivity. Curr. Opin. Virol. 2018, 33, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, R.; Lipkin, W.I. Discovery and Surveillance of Tick-Borne Pathogens. J. Med. Entomol. 2021, 58, 1525–1535. [Google Scholar] [CrossRef]
- Anderson, J.F.; Armstrong, P.M. Prevalence and genetic characterization of Powassan virus strains infecting Ixodes scapularis in Connecticut. Am. J. Trop. Med. Hyg. 2012, 87, 754–759. [Google Scholar] [CrossRef]
- Hermance, M.E.; Hart, C.E.; Esterly, A.T.; Reynolds, E.S.; Bhaskar, J.R.; Thangamani, S. Development of a small animal model for deer tick virus pathogenesis mimicking human clinical outcome. PLoS Neglected Trop. Dis. 2020, 14, e0008359. [Google Scholar] [CrossRef]
- Jasperse, B.A.; Mattocks, M.D.; Noll, K.E.; Ferris, M.T.; Heise, M.T.; Lazear, H.M. Neuroinvasive Flavivirus Pathogenesis Is Restricted by Host Genetic Factors in Collaborative Cross Mice, Independently of Oas1b. J. Virol. 2023, 97, e0071523. [Google Scholar] [CrossRef]
- Nemeth, N.M.; Root, J.J.; Hartwig, A.E.; Bowen, R.A.; Bosco-Lauth, A.M. Powassan Virus Experimental Infections in Three Wild Mammal Species. Am. J. Trop. Med. Hyg. 2021, 104, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Cull, B.; Wang, X.R. Methods for the Study of Ticks, Mosquitoes, and their Transmitted Pathogens: Toward a Greater Understanding of Vector Biology and Arthropod-Microbe Interactions. J. Vis. Exp. 2023, 193, e64986. [Google Scholar] [CrossRef] [PubMed]
- Garcia Guizzo, M.; Meneses, C.; Amado Cecilio, P.; Hessab Alvarenga, P.; Sonenshine, D.; Ribeiro, J.M. Optimizing tick artificial membrane feeding for Ixodes scapularis. Sci. Rep. 2023, 13, 16170. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, J.M.; Offerdahl, D.K.; Bloom, M.E. The Use of Ex Vivo Organ Cultures in Tick-Borne Virus Research. ACS Infect. Dis. 2018, 4, 247–256. [Google Scholar] [CrossRef]
- Rochlin, I.; Chu, D.; Gmelin, M.; Le, J.; Furie, M.B.; Thanassi, D.G.; Keun Kim, H. Optimization of artificial membrane feeding system for lone star ticks, Amblyomma americanum (Acari: Ixodidae), and experimental infection with Rickettsia amblyommatis (Rickettsiales: Rickettsiaceae). J. Med. Entomol. 2024, 61, 442–453. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate ID | Lineage | Location | Year | Source | Passage History | GenBank Accession |
|---|---|---|---|---|---|---|
| M11665 | I | Ontario | 1965 | I. cookei | P1SM1V1B1 | OP823404 |
| NFS9601 | II | Nantucket, MA | 1996 | I. scapularis | SM1B2 | HM440559 |
| ME19-1051 | II | Cape Elizabeth, ME | 2019 | I. scapularis | B1 | OP823442 |
| FA5/12-40 | II | Spooner, WI | 2008 | I. scapularis | B2 | OP823475 |
| NJ19-56 | II | Hardwick, NJ | 2019 | I. scapularis | B1 | OP823460 |
| M11665 | I | Ontario | 1965 | I. cookei | P1SM1V1B1 | OP823404 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McMinn, R.J.; Gallichotte, E.N.; Courtney, S.; Telford, S.R.; Ebel, G.D. Strain-Dependent Assessment of Powassan Virus Transmission to Ixodes scapularis Ticks. Viruses 2024, 16, 830. https://doi.org/10.3390/v16060830
McMinn RJ, Gallichotte EN, Courtney S, Telford SR, Ebel GD. Strain-Dependent Assessment of Powassan Virus Transmission to Ixodes scapularis Ticks. Viruses. 2024; 16(6):830. https://doi.org/10.3390/v16060830
Chicago/Turabian StyleMcMinn, Rebekah J., Emily N. Gallichotte, Samantha Courtney, Sam R. Telford, and Gregory D. Ebel. 2024. "Strain-Dependent Assessment of Powassan Virus Transmission to Ixodes scapularis Ticks" Viruses 16, no. 6: 830. https://doi.org/10.3390/v16060830
APA StyleMcMinn, R. J., Gallichotte, E. N., Courtney, S., Telford, S. R., & Ebel, G. D. (2024). Strain-Dependent Assessment of Powassan Virus Transmission to Ixodes scapularis Ticks. Viruses, 16(6), 830. https://doi.org/10.3390/v16060830