

Investigating the Genetic Diversity of Hepatitis Delta Virus in Hepatocellular Carcinoma (HCC): Impact on Viral Evolution and Oncogenesis in HCC

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Serum Samples
2.2. Analyses of HDV RNA from Serum Samples of HCC
2.3. HDV Genome and HDAg Expression Plasmids
2.4. Transfections and Post-Transfection Analyses
2.5. Western Blot Assays
2.6. Wound Healing Assay
2.7. Cell Invasion Assay
2.8. Immunofluorescence F-Actin Staining
3. Results
3.1. RT-PCR Sequencing Analyses of HDV RNA in the Sera of HCC
3.2. Genotypes of HDV Isolated from the Serum Samples of HCC Patients
3.3. Mixed-Genotype Infections and RNA Recombination in HDV-Associated HCC
3.4. Mutations Leading to Codon Changes in Single-Genotype Infections of HDV
3.5. Novel Mutations in Mixed-Genotype Infections of HDV
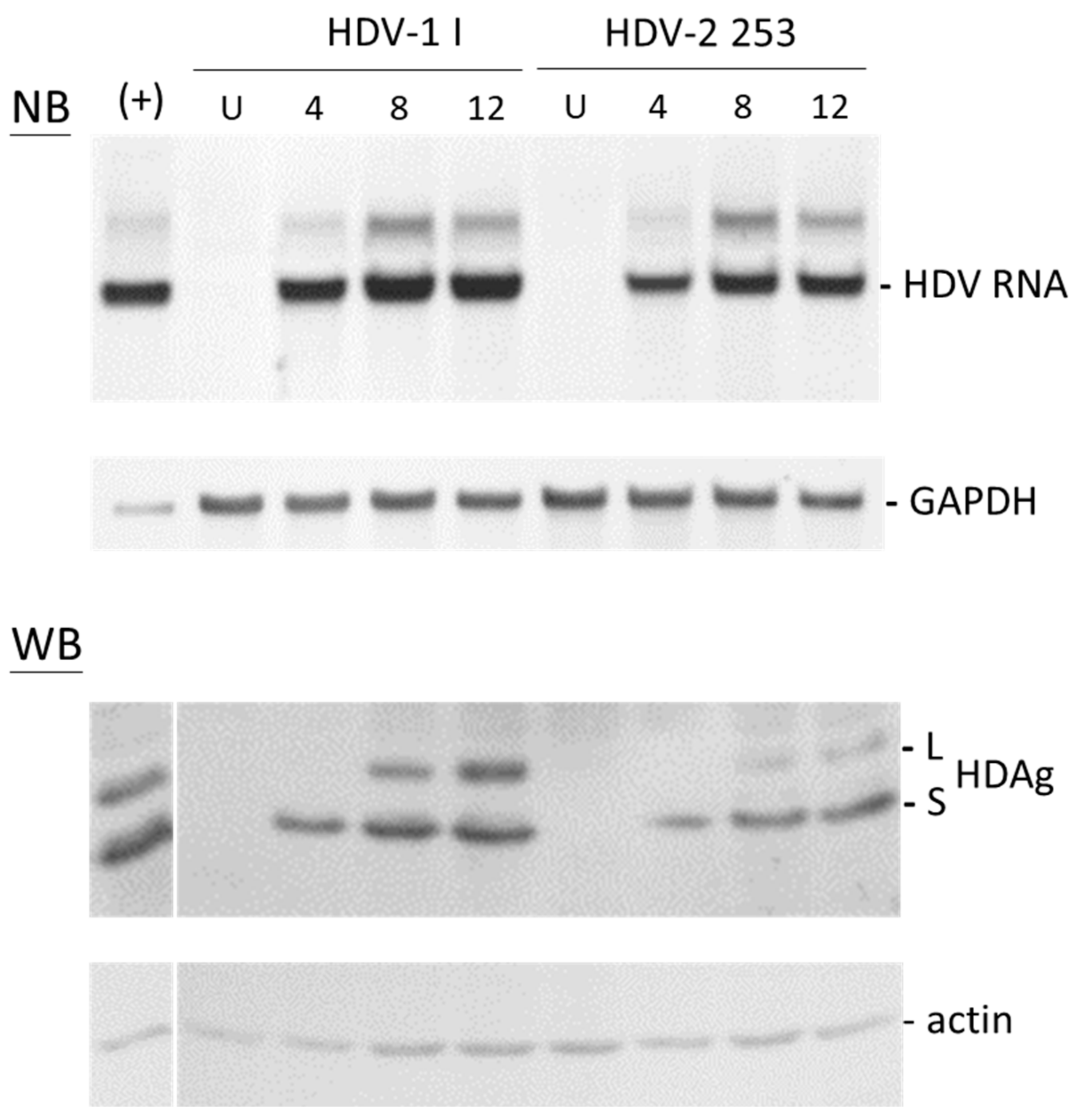
3.6. Replication of HDV-2 Clone 253 Isolated from the Serum Sample of HCC
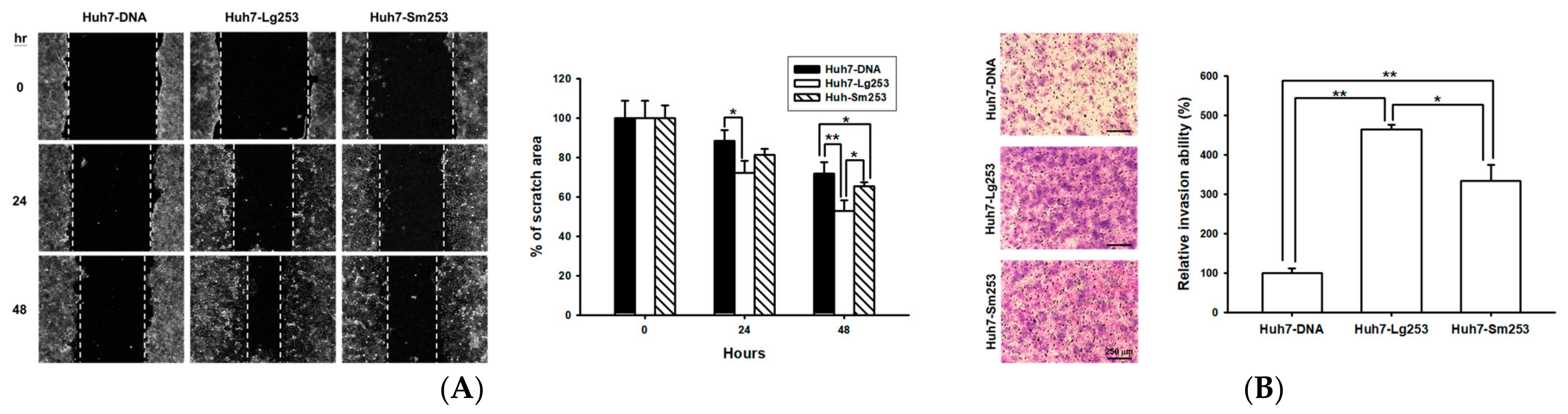
3.7. Both Forms of the HDAg Facilitate Cell Migration and Invasion
3.8. HDAg Boosts Cell Motility and Regulates the Expression of EMT Markers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rizzetto, M.; Hoyer, B.; Canese, M.G.; Shih, J.W.; Purcell, R.H.; Gerin, J.L. Delta Agent: Association of delta antigen with hepatitis B surface antigen and RNA in serum of delta-infected chimpanzees. Proc. Natl. Acad. Sci. USA 1980, 77, 6124–6128. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Shen, D.T.; Ji, D.Z.; Han, P.C.; Zhang, W.M.; Ma, J.F.; Chen, W.S.; Goyal, H.; Pan, S.; Xu, H.G. Prevalence and burden of hepatitis D virus infection in the global population: A systematic review and meta-analysis. Gut 2019, 68, 512–521. [Google Scholar] [CrossRef]
- Miao, Z.; Zhang, S.; Ou, X.; Li, S.; Ma, Z.; Wang, W.; Peppelenbosch, M.P.; Liu, J.; Pan, Q. Estimating the global prevalence, disease progression, and clinical outcome of hepatitis delta virus infection. J. Infect. Dis. 2020, 221, 1677–1687. [Google Scholar] [CrossRef] [PubMed]
- Stockdale, A.J.; Kreuels, B.; Henrion, M.Y.R.; Giorgi, E.; Kyomuhangi, I.; de Martel, C.; Hutin, Y.; Geretti, A.M. The global prevalence of hepatitis D virus infection: Systematic review and meta-analysis. J. Hepatol. 2020, 73, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Polaris Observatory Collaborators. Adjusted estimate of the prevalence of hepatitis delta virus in 25 countries and territories. J. Hepatol. 2024, 80, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.; Meyer, T.; Sapisochin, G.; Salem, R.; Saborowski, A. Hepatocellular carcinoma. Lancet 2022, 400, 1345–1362. [Google Scholar] [CrossRef]
- Alfaiate, D.; Clement, S.; Gomes, D.; Goossens, N.; Negro, F. Chronic hepatitis D and hepatocellular carcinoma: A systematic review and meta-analysis of observational studies. J. Hepatol. 2020, 73, 533–539. [Google Scholar] [CrossRef]
- Kamal, H.; Fornes, R.; Simin, J.; Stål, P.; Duberg, A.S.; Brusselaers, N.; Aleman, S. Risk of hepatocellular carcinoma in hepatitis B and D virus co-infected patients: A systematic review and meta-analysis of longitudinal studies. J. Viral Hepat. 2021, 28, 1431–1442. [Google Scholar] [CrossRef]
- Lai, M.M. RNA replication without RNA-dependent RNA polymerase: Surprises from hepatitis delta virus. J. Virol. 2005, 79, 7951–7958. [Google Scholar] [CrossRef]
- Taylor, J.M. Virology of hepatitis D virus. Semin. Liver Dis. 2012, 32, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Casey, J.L. Control of ADAR1 editing of hepatitis delta virus RNAs. Curr. Top. Microbiol. Immunol. 2012, 353, 123–143. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.Y.; Chao, M.; Taylor, J. Initiation of replication of the human hepatitis delta virus genome from cloned DNA: Role of delta antigen. J. Virol. 1989, 63, 1945–1950. [Google Scholar] [CrossRef] [PubMed]
- Chang, F.L.; Chen, P.J.; Tu, S.J.; Wang, C.J.; Chen, D.S. The large form of hepatitis delta antigen is crucial for assembly of hepatitis delta virus. Proc. Natl. Acad. Sci. USA 1991, 88, 8490–8494. [Google Scholar] [CrossRef] [PubMed]
- Goodrum, G.; Pelchat, M. Insight into the contribution and disruption of host processes during HDV replication. Viruses 2018, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.J.; Sun, C.P.; Hsu, Y.C.; Chen, Y.W.; Wang, I.A.; Su, C.W.; Tao, M.H.; Wu, J.C. Statin inhibits large hepatitis delta antigen-Smad3-twist-mediated epithelial-to-mesenchymal transition and hepatitis D virus secretion. J. Biomed. Sci. 2020, 27, 65. [Google Scholar] [CrossRef] [PubMed]
- Dény, P. Hepatitis delta virus genetic variability: From genotypes I, II, III to eight major clades? Curr. Top. Microbiol. Immunol. 2006, 307, 151–171. [Google Scholar] [CrossRef]
- Le Gal, F.; Brichler, S.; Drugan, T.; Alloui, C.; Roulot, D.; Pawlotsky, J.M.; Deny, P.; Gordien, E. Genetic diversity and worldwide distribution of the deltavirus genus: A study of 2152 clinical strains. Hepatology 2017, 66, 1826–1841. [Google Scholar] [CrossRef]
- Casey, J.L.; Brown, T.L.; Colan, E.J.; Wignall, F.S.; Gerin, J.L. A genotype of hepatitis D virus that occurs in northern South America. Proc. Natl. Acad. Sci. USA 1993, 90, 9016–9020. [Google Scholar] [CrossRef]
- Su, C.W.; Huang, Y.H.; Huo, T.I.; Shih, H.H.; Sheen, I.J.; Chen, S.W.; Lee, P.C.; Lee, S.D.; Wu, J.C. Genotypes and viremia of hepatitis B and D viruses are associated with outcomes of chronic hepatitis D patients. Gastroenterology 2006, 130, 1625–1635. [Google Scholar] [CrossRef]
- Bahoussi, A.N.; Wang, P.H.; Guo, Y.Y.; Rabbani, N.; Wu, C.; Xing, L. Global distribution and natural recombination of hepatitis D virus: Implication of Kyrgyzstan emerging HDVs in the clinical outcomes. Viruses 2022, 14, 1467. [Google Scholar] [CrossRef]
- Wang, T.C.; Chao, M. RNA recombination of hepatitis delta virus in natural mixed-genotype infection and transfected cultured cells. J. Virol. 2005, 79, 2221–2229. [Google Scholar] [CrossRef] [PubMed]
- Sy, B.T.; Nguyen, H.M.; Toan, N.L.; Song, L.H.; Tong, H.V.; Wolboldt, C.; Binh, V.Q.; Kremsner, P.G.; Velavan, T.P.; Bock, C.T. Identification of a natural intergenotypic recombinant hepatitis delta virus genotype 1 and 2 in Vietnamese HBsAg-positive patients. J. Viral Hepat. 2015, 22, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Lee, C.C.; Lin, S.H.; Huang, P.J.; Li, H.P.; Chang, Y.S.; Tang, P.; Chao, M. RNA recombination in Hepatitis delta virus: Identification of a novel naturally occurring recombinant. J. Microbiol. Immunol. Infect. Wei Mian Yu Gan Ran Za Zhi 2017, 50, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Zhang, S.; Ma, Z.; Hakim, M.S.; Wang, W.; Peppelenbosch, M.P.; Pan, Q. Recombinant identification, molecular classification and proposed reference genomes for hepatitis delta virus. J. Viral Hepat. 2019, 26, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.Y.; Wu, Y.L.; Wang, C.S.; Ko, C.Y.; Chen, C.H.; Chen, P.J.; Peng, P.H.; Hsu, C.W. Performance of commercially available anti-HDV enzyme-linked immunosorbent assays in Taiwan. Virol. J. 2020, 17, 76. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.W.; Hsu, H.Y.; Chen, C.H.; Chao, M. Unbranched rod-like RNA is required for RNA editing of hepatitis delta virus genotype 2 and genotype 4. Virus Res. 2023, 338, 199239. [Google Scholar] [CrossRef]
- Makarova, O.; Kamberov, E.; Margolis, B. Generation of deletion and point mutations with one primer in a single cloning step. Biotechniques 2000, 29, 970–972. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, H.; Taketa, K.; Miyano, K.; Yamane, T.; Sato, J. Growth of human hepatoma cells lines with differentiated functions in chemically defined medium. Cancer Res. 1982, 42, 3858–3863. Available online: https://pubmed.ncbi.nlm.nih.gov/6286115/ (accessed on 19 November 2014).
- Chao, M.; Lin, C.C.; Lin, F.M.; Li, H.P.; Iang, S.B. Whole-genome analysis of genetic recombination of hepatitis delta virus: Molecular domain in delta antigen determining trans-activating efficiency. J. Gen. Virol. 2015, 96, 3460–3469. [Google Scholar] [CrossRef]
- Chiang, K.C.; Chang, K.S.; Hsu, S.Y.; Sung, H.C.; Feng, T.H.; Chao, M.; Juang, H.H. Human heme oxygenase-1 induced by interleukin-6 via JAK/STAT3 pathways is a tumor suppressor gene in hepatoma cells. Antioxidants 2020, 9, 251. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.C.; Chao, M. Molecular cloning and expression of the hepatitis delta virus genotype IIb genome. Biochem. Biophys. Res. Commun. 2003, 303, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.S.; Chen, S.T.; Sung, H.C.; Hsu, S.Y.; Lin, W.Y.; Hou, C.P.; Lin, Y.H.; Feng, T.H.; Tsui, K.H.; Juang, H.H. WNT1 inducible signaling pathway protein 1 is a stroma-specific secreting protein inducing a fibroblast contraction and carcinoma cell growth in the human prostate. Int. J. Mol. Sci. 2022, 23, 11437. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.; Changchien, C.S.; Chung, J.C.; Liaw, Y.F. Characterization of a new genotype II hepatitis delta virus from Taiwan. J. Med. Virol. 1996, 49, 145–154. [Google Scholar] [CrossRef]
- Lin, F.M.; Lee, C.M.; Wang, T.C.; Chao, M. Initiation of RNA replication of cloned Taiwan-3 isolate of hepatitis delta virus genotype II in cultured cells. Biochem. Biophys. Res. Commun. 2003, 306, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.; Wang, T.C.; Lin, C.C.; Wang, R.Y.L.; Lin, W.B.; Lee, S.E.; Cheng, Y.Y.; Yeh, C.T.; Iang, S.B. Analyses of a whole-genome inter-clade recombination map of hepatitis delta virus suggest a host polymerase-driven and viral RNA structure-promoted template-switching mechanism for viral RNA recombination. Oncotarget 2017, 8, 60841–60859. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Yang, Z.W.; Iang, S.B.; Chao, M. Reduced genetic distance and high replication levels increase the RNA recombination rate of hepatitis delta virus. Virus Res. 2015, 195, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.; Lin, Y.Y.; Chen, P.J. Recombination between hepatitis D viruses. In Hepatitis D. Virology, Management and Methodology; Rizzetto, M., Smedile, A., Eds.; Il Pensiero Scientifico: Rome, Italy, 2019; pp. 135–154. ISBN 8849006462, 9788849006469. [Google Scholar]
- Hsieh, S.Y.; Chao, M.; Coates, L.; Taylor, J. Hepatitis delta virus genome replication: A polyadenylated mRNA for delta antigen. J. Virol. 1990, 64, 3192–3198. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.Y.; Goldberg, J.; Coates, L.; Mason, W.; Gerin, J.; Taylor, J. Molecular cloning of hepatitis delta virus RNA from an infected woodchuck liver: Sequence, structure, and applications. J. Virol. 1988, 62, 1855–1861. [Google Scholar] [CrossRef]
- Wu, J.C. Functional and clinical significance of hepatitis D virus genotype II infection. Curr. Top. Microbiol. Immunol. 2006, 307, 173–186. [Google Scholar] [CrossRef]
- Chang, S.Y.; Yang, C.L.; Ko, W.S.; Liu, W.C.; Lin, C.Y.; Wu, C.H.; Su, Y.C.; Chang, S.F.; Chen, M.Y.; Sheng, W.H.; et al. Molecular epidemiology of hepatitis D virus infection among injecting drug users with and without human immunodeficiency virus infection in Taiwan. J. Clin. Microbiol. 2011, 49, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.H.; Lee, S.S.; Yu, M.L.; Chang, T.T.; Su, C.W.; Hu, B.S.; Chen, Y.S.; Huang, C.K.; Lai, C.H.; Lin, J.N.; et al. Changing hepatitis D virus epidemiology in a hepatitis B virus endemic area with a national vaccination program. Hepatology 2015, 61, 1870–1879. [Google Scholar] [CrossRef] [PubMed]
- Dziri, S.; Rodriguez, C.; Gerber, A.; Brichler, S.; Alloui, C.; Roulot, D.; Dény, P.; Pawlotsky, J.M.; Gordien, E.; Le Gal, F. Variable in vivo hepatitis D virus (HDV) RNA editing rates according to the HDV genotype. Viruses 2021, 13, 1572. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.C.; Huang, I.A.; Huang, Y.H.; Chen, J.Y.; Sheen, I.J. Mixed genotypes infection with hepatitis D virus. J. Med. Virol. 1999, 57, 64–67. [Google Scholar] [CrossRef]
- Kao, J.H.; Chen, P.J.; Lai, M.Y.; Chen, D.S. Hepatitis D virus genotypes in intravenous drug users in taiwan: Decreasing prevalence and lack of correlation with hepatitis B virus genotypes. J. Clin. Microbiol. 2002, 40, 3047–3049. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.C.; Choo, K.B.; Chen, C.M.; Chen, T.Z.; Huo, T.I.; Lee, S.D. Genotyping of hepatitis D virus by restriction-fragment length polymorphism and relation to outcome of hepatitis D. Lancet 1995, 346, 939–941. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.Y.; Taylor, J. Regulation of polyadenylation of hepatitis delta virus antigenomic RNA. J. Virol. 1991, 65, 6438–6446. [Google Scholar] [CrossRef] [PubMed]
- Fouad, R.; Abdo, M.; Eldeen, H.G.; Sabry, D.; Atef, M.; Ahmed, R.; Zayed, N. Influence of delta virus infection on the virologic status in Egyptian patients with chronic hepatitis B virus genotype D. J. Med. Virol. 2016, 88, 837–842. [Google Scholar] [CrossRef]
- Barros, L.M.; Gomes-Gouvea, M.S.; Pinho, J.R.; Alvarado-Mora, M.V.; Dos Santos, A.; Mendes-Correa, M.C.; Caldas, A.J.; Sousa, M.T.; Santos, M.D.; Ferreira, A.S. Hepatitis delta virus genotype 8 infection in Northeast Brazil: Inheritance from African slaves? Virus Res. 2011, 160, 333–339. [Google Scholar] [CrossRef]
- Demirel, A.; Uraz, S.; Deniz, Z.; Daglilar, E.; Basar, O.; Tahan, V.; Ozaras, R. Epidemiology of hepatitis D virus infection in Europe: Is it vanishing? J. Viral Hepat. 2024, 31, 120–128. [Google Scholar] [CrossRef]
- Roulot, D.; Brichler, S.; Layese, R.; BenAbdesselam, Z.; Zoulim, F.; Thibault, V.; Scholtes, C.; Roche, B.; Castelnau, C.; Poynard, T.; et al. Origin, HDV genotype and persistent viremia determine outcome and treatment response in patients with chronic hepatitis delta. J. Hepatol. 2020, 73, 1046–1062. [Google Scholar] [CrossRef] [PubMed]
- Spaan, M.; Carey, I.; Bruce, M.; Shang, D.; Horner, M.; Dusheiko, G.; Agarwal, K. Hepatitis delta genotype 5 is associated with favourable disease outcome and better response to treatment compared to genotype 1. J. Hepatol. 2020, 72, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Shih, H.H.; Sheen, I.J.; Su, C.W.; Peng, W.L.; Lin, L.H.; Wu, J.C. Hepatitis D virus isolates with low replication and epithelial-mesenchymal transition-inducing activity are associated with disease remission. J. Virol. 2012, 86, 9044–9054. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lempp, F.A.; Schlund, F.; Walter, L.; Decker, C.C.; Zhang, Z.; Ni, Y.; Urban, S. Assembly and infection efficacy of hepatitis B virus surface protein exchanges in 8 hepatitis D virus genotype isolates. J. Hepatol. 2021, 75, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Kato, N.; Ono-Nita, S.K.; Yoshida, H.; Otsuka, M.; Shiratori, Y.; Omata, M. Large isoform of hepatitis delta antigen activates serum response factor-associated transcription. J. Biol. Chem. 2000, 275, 37311–37316. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Jeong, S.H.; Hwang, S.B. Large hepatitis delta antigen modulates transforming growth factor-beta signaling cascades: Implication of hepatitis delta virus-induced liver fibrosis. Gastroenterology 2007, 132, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Oh, S.H.; Kang, S.M.; Lim, Y.S.; Hwang, S.B. Hepatitis delta virus large antigen sensitizes to TNF-alpha-induced NF-kappaB signaling. Mol. Cells 2009, 28, 49–55. [Google Scholar] [CrossRef]
- Williams, V.; Brichler, S.; Khan, E.; Chami, M.; Deny, P.; Kremsdorf, D.; Gordien, E. Large hepatitis delta antigen activates STAT-3 and NF-kappaB via oxidative stress. J. Viral Hepat. 2012, 19, 744–753. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juang, H.-H.; Hsu, C.-W.; Chang, K.-S.; Iang, S.-B.; Lin, Y.-H.; Chao, M. Investigating the Genetic Diversity of Hepatitis Delta Virus in Hepatocellular Carcinoma (HCC): Impact on Viral Evolution and Oncogenesis in HCC. Viruses 2024, 16, 817. https://doi.org/10.3390/v16060817
Juang H-H, Hsu C-W, Chang K-S, Iang S-B, Lin Y-H, Chao M. Investigating the Genetic Diversity of Hepatitis Delta Virus in Hepatocellular Carcinoma (HCC): Impact on Viral Evolution and Oncogenesis in HCC. Viruses. 2024; 16(6):817. https://doi.org/10.3390/v16060817
Chicago/Turabian StyleJuang, Horng-Heng, Chao-Wei Hsu, Kang-Shuo Chang, Shan-Bei Iang, Yang-Hsiang Lin, and Mei Chao. 2024. "Investigating the Genetic Diversity of Hepatitis Delta Virus in Hepatocellular Carcinoma (HCC): Impact on Viral Evolution and Oncogenesis in HCC" Viruses 16, no. 6: 817. https://doi.org/10.3390/v16060817
APA StyleJuang, H.-H., Hsu, C.-W., Chang, K.-S., Iang, S.-B., Lin, Y.-H., & Chao, M. (2024). Investigating the Genetic Diversity of Hepatitis Delta Virus in Hepatocellular Carcinoma (HCC): Impact on Viral Evolution and Oncogenesis in HCC. Viruses, 16(6), 817. https://doi.org/10.3390/v16060817