
A Prospective Study Investigating Immune Checkpoint Molecule and CD39 Expression on Peripheral Blood Cells for the Prognostication of COVID-19 Severity and Mortality

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Data Extraction and Outcome Measure
2.3. Peripheral Blood Mononuclear Cell Isolation
2.4. Flow Cytometry
2.5. Statistics
3. Results
3.1. Patient Characteristics
3.2. Flow Cytometry
3.3. Univariable and Multivariable Analysis
4. Discussion
Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fact Sheet: End of the COVID-19 Public Health Emergency. Available online: https://www.hhs.gov/about/news/2023/05/09/fact-sheet-end-of-the-covid-19-public-health-emergency.html (accessed on 2 May 2024).
- Tian, W.; Jiang, W.; Yao, J.; Nicholson, C.J.; Li, R.H.; Sigurslid, H.H.; Wooster, L.; Rotter, J.I.; Guo, X.; Malhotra, R. Predictors of mortality in hospitalized COVID-19 patients: A systematic review and meta-analysis. J. Med. Virol. 2020, 92, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Kojima, S.; Kawamoto, A.; Fukushima, M. COVID-19 pathogenesis, prognostic factors, and treatment strategy: Urgent recommendations. J. Med. Virol. 2021, 93, 2694–2704. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, D.Q.; Zou, B.; Yang, H.; Hui, W.Z.; Rui, F.; Yee, N.T.S.; Liu, C.; Nerurkar, S.N.; Kai, J.C.Y.; et al. Epidemiology of COVID-19: A systematic review and meta-analysis of clinical characteristics, risk factors, and outcomes. J. Med. Virol. 2021, 93, 1449–1458. [Google Scholar] [CrossRef]
- Gambichler, T.; Schuleit, N.; Susok, L.; Becker, J.C.; Scheel, C.H.; Torres-Reyes, C.; Overheu, O.; Reinacher-Schick, A.; Schmidt, W. Prognostic Performance of Inflammatory Biomarkers Based on Complete Blood Counts in COVID-19 Patients. Viruses 2023, 15, 1920. [Google Scholar] [CrossRef]
- Park, L.M.; Lannigan, J.; Jaimes, M.C. OMIP-069: Forty-Color Full Spectrum Flow Cytometry Panel for Deep Immunophenotyping of Major Cell Subsets in Human Peripheral Blood. Cytom. A 2020, 97, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Gambichler, T.; Reuther, J.; Scheel, C.H.; Becker, J.C. On the use of immune checkpoint inhibitors in patients with viral infections including COVID-19. J. Immunother. Cancer 2020, 8, e001145. [Google Scholar] [CrossRef] [PubMed]
- Alahdal, M.; Elkord, E. Exhaustion and over-activation of immune cells in COVID-19: Challenges and therapeutic opportunities. Clin. Immunol. 2022, 245, 109177. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Liu, G.; Zhong, J.; Zheng, K.; Xiao, H.; Li, C.; Song, X.; Li, Y.; Xu, C.; Wu, H.; et al. Immune Checkpoints in Viral Infections. Viruses 2020, 12, 1051. [Google Scholar] [CrossRef] [PubMed]
- Al-Mterin, M.A.; Alsalman, A.; Elkord, E. Inhibitory Immune Checkpoint Receptors and Ligands as Prognostic Biomarkers in COVID-19 Patients. Front. Immunol. 2022, 13, 870283. [Google Scholar] [CrossRef]
- Barnova, M.; Bobcakova, A.; Urdova, V.; Kosturiak, R.; Kapustova, L.; Dobrota, D.; Jesenak, M. Inhibitory immune checkpoint molecules and exhaustion of T cells in COVID-19. Physiol. Res. 2021, 70, S227–S247. [Google Scholar] [CrossRef]
- Gambichler, T.; Reuther, J.; Scheel, C.H.; Susok, L.; Kern, P.; Becker, J.C. Cancer and Immune Checkpoint Inhibitor Treatment in the Era of SARS-CoV-2 Infection. Cancers 2020, 12, 3383. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Jeong, S.; Jeon, K.; Park, M.J.; Song, W. Prognostic impacts of soluble immune checkpoint regulators and cytokines in patients with SARS-CoV-2 infection. Front. Immunol. 2022, 13, 903419. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Ampudia Mesias, E.; Pluhar, G.E.; Rathe, S.K.; Largaespada, D.A.; Sham, Y.Y.; Moertel, C.L.; Olin, M.R. Cd200 checkpoint reversal: A novel approach to immunotherapy. Clin. Cancer Res. 2020, 26, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Avendaño-Ortiz, J.; Lozano-Rodríguez, R.; Martín-Quirós, A.; Terrón, V.; Maroun-Eid, C.; Montalbán-Hernández, K.; Valentín-Quiroga, J.; García-Garrido, M.Á.; Del Val, E.M.; del Balzo-Castillo, Á.; et al. The immune checkpoints storm in COVID-19: Role as severity markers at emergency department admission. Clin. Transl. Med. 2021, 11, e573. [Google Scholar] [CrossRef] [PubMed]
- Jubel, J.M.; Barbati, Z.R.; Burger, C.; Wirtz, D.C.; Schildberg, F.A. The Role of PD-1 in Acute and Chronic Infection. Front. Immunol. 2020, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- Ahn, E.; Araki, K.; Hashimoto, M.; Li, W.; Riley, J.L.; Cheung, J.; Sharpe, A.H.; Freeman, G.J.; Irving, B.A.; Ahmed, R. Role of PD-1 during effector CD8 T cell differentiation. Proc. Natl. Acad. Sci. USA 2018, 115, 4749–4754. [Google Scholar] [CrossRef] [PubMed]
- Schönrich, G.; Raftery, M.J. The PD-1/PD-L1 Axis and Virus Infections: A Delicate Balance. Front. Cell Infect. Microbiol. 2019, 9, 207. [Google Scholar] [CrossRef] [PubMed]
- Avery, L.; Filderman, J.; Szymczak-Workman, A.L.; Kane, L.P. Tim-3 co-stimulation promotes short-lived effector T cells, restricts memory precursors, and is dispensable for T cell exhaustion. Proc. Natl. Acad. Sci. USA 2018, 115, 2455–2460. [Google Scholar] [CrossRef] [PubMed]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Juno, J.A.; Tan, H.X.; Lee, W.S.; Reynaldi, A.; Kelly, H.G.; Wragg, K.; Esterbauer, R.; Kent, H.E.; Batten, C.J.; Mordant, F.L.; et al. Humoral and circulating follicular helper T cell responses in recovered patients with COVID-19. Nat. Med. 2020, 26, 1428–1434. [Google Scholar] [CrossRef]
- Boppana, S.; Qin, K.; Files, J.K.; Russell, R.M.; Stoltz, R.; Bibollet-Ruche, F.; Bansal, A.; Erdmann, N.; Hahn, B.H.; Goepfert, P.A. SARS-CoV-2-specific circulating T follicular helper cells correlate with neutralizing antibodies and increase during early convalescence. PLoS Pathog. 2021, 17, e1009761. [Google Scholar] [CrossRef] [PubMed]
- Notarbartolo, S.; Ranzani, V.; Bandera, A.; Gruarin, P.; Bevilacqua, V.; Putignano, A.R.; Gobbini, A.; Galeota, E.; Manara, C.; Bombaci, M.; et al. Integrated longitudinal immunophenotypic, transcriptional and repertoire analyses delineate immune responses in COVID-19 patients. Sci. Immunol. 2021, 6, eabg5021. [Google Scholar] [CrossRef] [PubMed]
- Cohen, K.W.; Linderman, S.L.; Moodie, Z.; Czartoski, J.; Lai, L.; Mantus, G.; Norwood, C.; Nyhoff, L.E.; Edara, V.V.; Floyd, K.; et al. Longitudinal analysis shows durable and broad immune memory after SARS-CoV-2 infection with persisting antibody responses and memory B and T cells. Cell Rep. Med. 2021, 2, 100354. [Google Scholar] [CrossRef]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Liu, Y.; Wang, G.; et al. Reduction and Functional Exhaustion of T Cells in Patients With Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef] [PubMed]
- Rha, M.S.; Jeong, H.W.; Ko, J.H.; Choi, S.J.; Seo, I.H.; Lee, J.S.; Sa, M.; Kim, A.R.; Joo, E.J.; Ahn, J.Y.; et al. PD-1-Expressing SARS-CoV-2-Specific CD8+ T Cells Are Not Exhausted, but Functional in Patients with COVID-19. Immunity 2021, 54, 44–52.e3. [Google Scholar] [CrossRef]
- Elsaghir, A.; El-Sabaa, E.M.; Ahmed, A.K.; Abdelwahab, S.F.; Sayed, I.M.; El-Mokhtar, M.A. The Role of Cluster of Differentiation 39 (CD39) and Purinergic Signaling Pathway in Viral Infections. Pathogens 2023, 12, 279. [Google Scholar] [CrossRef]
- Franciosi, M.L.M.; Lima, M.D.M.; Schetinger, M.R.C.; Cardoso, A.M. Possible role of purinergic signaling in COVID-19. Mol. Cell Biochem. 2021, 476, 2891–2898. [Google Scholar] [CrossRef]
- Timperi, E.; Barnaba, V. CD39 Regulation and Functions in T Cells. Int. J. Mol. Sci. 2021, 22, 8068. [Google Scholar] [CrossRef]
- Modabber, Z.; Shahbazi, M.; Akbari, R.; Bagherzadeh, M.; Firouzjahi, A.; Mohammadnia-Afrouzi, M. TIM-3 as a potential exhaustion marker in CD4+ T cells of COVID-19 patients. Immun. Inflamm. Dis. 2021, 9, 1707–1715. [Google Scholar] [CrossRef]
- Farfour, E.; Lesprit, P.; Visseaux, B.; Pascreau, T.; Jolly, E.; Houhou, N.; Mazaux, L.; Asso-Bonnet, M.; Vasse, M.; SARS-CoV-2 Foch Hospital Study Group. The Allplex 2019-nCoV (Seegene) assay: Which performances are for SARS-CoV-2 infection diagnosis? Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1997–2000. [Google Scholar] [CrossRef]
- Marshall, J.C.; Murthy, S.; Diaz, J.; Adhikari, N.K.; Angus, D.C.; Arabi, Y.M.; Baillie, K.; Bauer, M.; Berry, S.; Blackwood, B.; et al. A minimal common outcome measure set for COVID-19 clinical research. Lancet Infect. Dis. 2020, 20, e192–e197. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.B.; Briggs, K.T.; Marino, J.P.; Ravid, K.; Robson, S.C.; Mosser, D.M. TLR stimulation initiates a CD39-based autoregulatory mechanism that limits macrophage inflammatory responses. Blood 2013, 122, 1935–1945. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purine and purinergic receptors. Brain Neurosci. Adv. 2018, 2, 2398212818817494. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.-F.; Guidotti, G. CD39 Is an Ecto-(Ca2+, Mg2+)-apyrase (*). J. Biol. Chem. 1996, 271, 9898–9901. [Google Scholar]
- Kronlage, M.; Song, J.; Sorokin, L.; Isfort, K.; Schwerdtle, T.; Leipziger, J.; Robaye, B.; Conley, P.B.; Kim, H.C.; Sargin, S.; et al. Autocrine purinergic receptor signaling is essential for macrophage chemotaxis. Sci. Signal. 2010, 3, ra55. [Google Scholar] [CrossRef] [PubMed]
- Dorneles, G.P.; Teixeira, P.C.; da Silva, I.M.; Schipper, L.L.; Santana Filho, P.C.; Rodrigues Junior, L.C.; Bonorino, C.; Peres, A.; Fonseca, S.G.; Monteiro, M.C.; et al. Alterations in CD39/CD73 axis of T cells associated with COVID-19 severity. J. Cell Physiol. 2022, 237, 3394–3407. [Google Scholar] [CrossRef] [PubMed]
- Shahbaz, S.; Xu, L.; Sligl, W.; Osman, M.; Bozorgmehr, N.; Mashhouri, S.; Redmond, D.; Perez Rosero, E.; Walker, J.; Elahi, S. The Quality of SARS-CoV-2-Specific T Cell Functions Differs in Patients with Mild/Moderate versus Severe Disease, and T Cells Expressing Coinhibitory Receptors Are Highly Activated. J. Immunol. 2021, 207, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Pietrobon, A.J.; Andrejew, R.; Custódio, R.W.A.; Oliveira, L.D.M.; Scholl, J.N.; Teixeira, F.M.E.; De Brito, C.A.; Glaser, T.; Kazmierski, J.; Aoki, V.; et al. Dysfunctional purinergic signaling correlates with disease severity in COVID-19 patients. Front. Immunol. 2022, 13, 1012027. [Google Scholar] [CrossRef] [PubMed]
- Simsek, A.; Kizmaz, M.A.; Cagan, E.; Dombaz, F.; Tezcan, G.; Asan, A.; Demir, H.I.; Bal, S.H.; Ermis, D.Y.; Dilektaslı, A.G.; et al. Assessment of CD39 expression in regulatory T-cell subsets by disease severity in adult and juvenile COVID-19 cases. J. Med. Virol. 2022, 94, 2089–2101. [Google Scholar] [CrossRef]
- Díaz-García, E.; García-Tovar, S.; Alfaro, E.; Zamarrón, E.; Mangas, A.; Galera, R.; Ruíz-Hernández, J.J.; Solé-Violán, J.; Rodríguez-Gallego, C.; Van-Den-Rym, A.; et al. Role of CD39 in COVID-19 Severity: Dysregulation of Purinergic Signaling and Thromboinflammation. Front. Immunol. 2022, 13, 847894. [Google Scholar] [CrossRef]
- Shahbazi, M.; Moulana, Z.; Sepidarkish, M.; Bagherzadeh, M.; Rezanejad, M.; Mirzakhani, M.; Jafari, M.; Mohammadnia-Afrouzi, M. Pronounce expression of Tim-3 and CD39 but not PD1 defines CD8 T cells in critical COVID-19 patients. Microb. Pathog. 2021, 153, 104779. [Google Scholar] [CrossRef] [PubMed]
- da Silva, G.B.; Manica, D.; da Silva, A.P.; Kosvoski, G.C.; Hanauer, M.; Assmann, C.E.; Simões, J.L.B.; Pillat, M.M.; de Lara, J.D.; Marafon, F.; et al. High levels of extracellular ATP lead to different inflammatory responses in COVID-19 patients according to the severity. J. Mol. Med. 2022, 100, 645–663. [Google Scholar] [CrossRef] [PubMed]
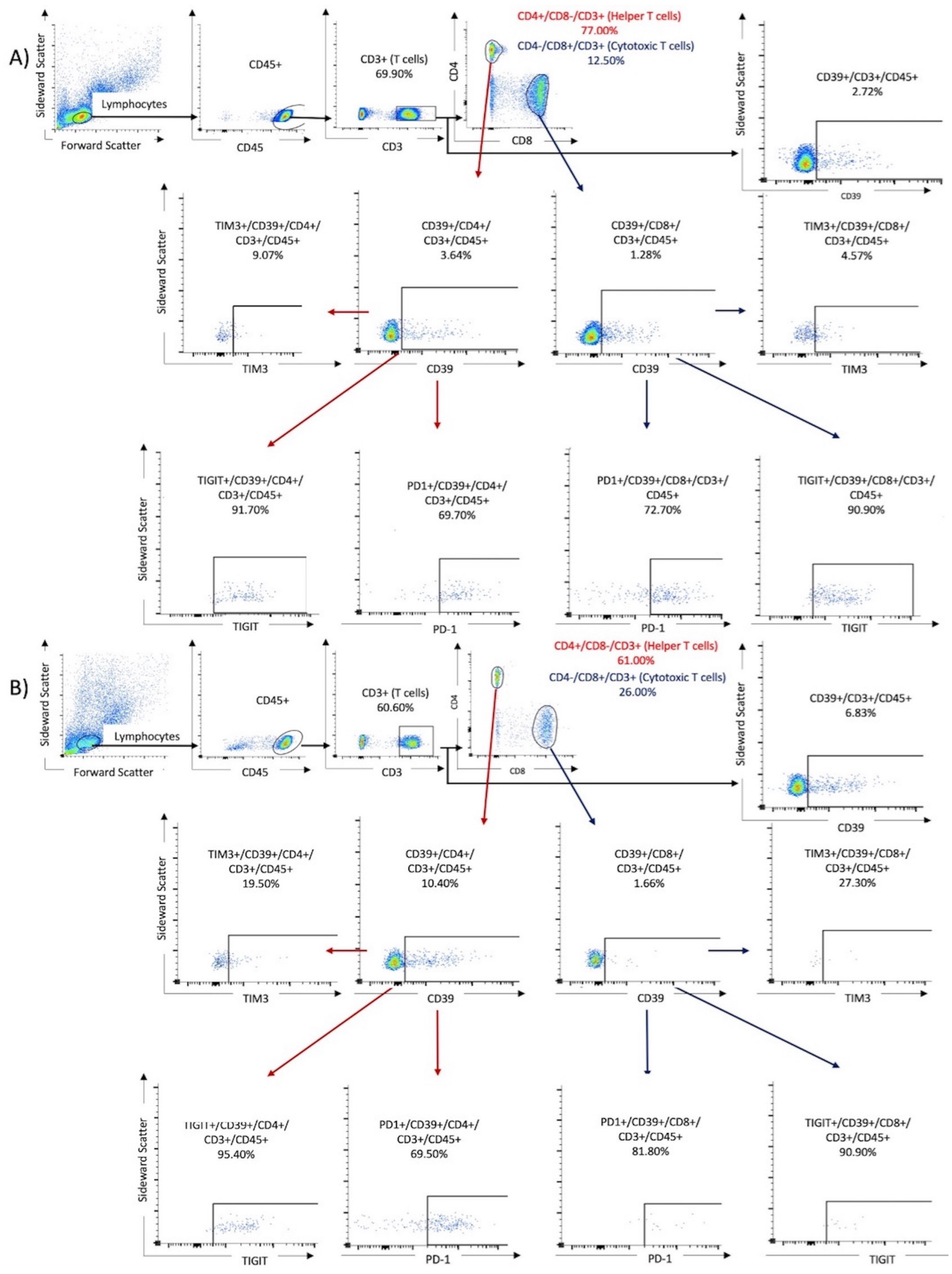

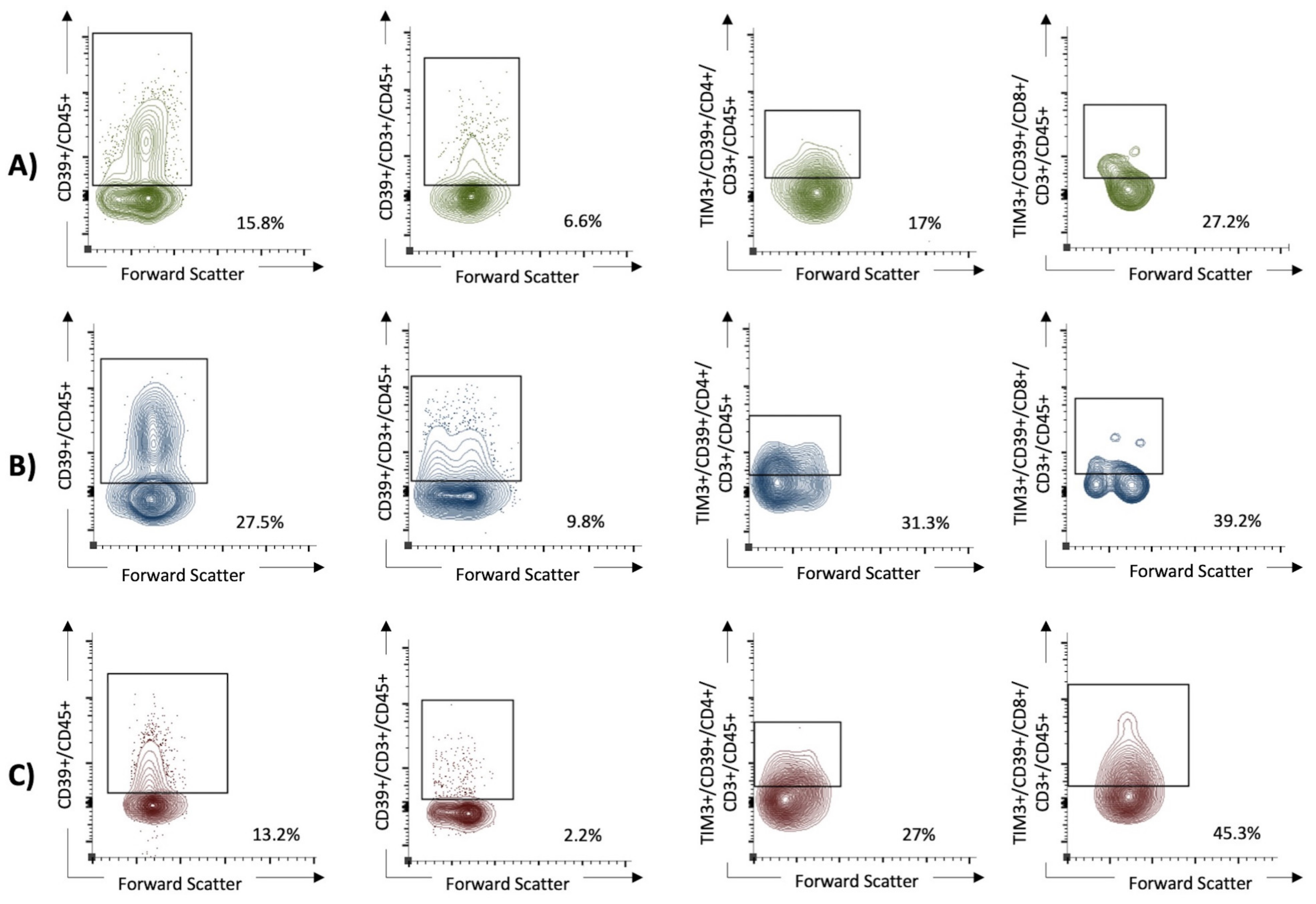

{kind=link}
{kind=link}
{kind=link}
| Parameter | Data |
|---|---|
| Sex | |
| COVID-19 patients | 47 m/29 f (61.8%/38.2%) |
| Healthy controls | 8 m/4 f (66.7%/33.3%); p = 0.99 |
| Median (range) age | |
| COVID-19 patients | 64 years (22–87) |
| Healthy controls | 63 years (47–73); p = 0.61 |
| SARS-CoV-2 variant | |
| Delta/omicron | 6/70 (7.9%/92.1%) |
| At least one vaccination | |
| No/yes | 8/68 (10.5%/89.5%) |
| Two or more relevant comorbidities * | |
| No/yes | 29/47 (38.2%/61.8%) |
| Smoking | |
| No/yes | 56/20 (73.7%/26.3%) |
| Breathing support | |
| No/yes | 61/15 (80.3%/19.7%) |
| COVID-19 pneumonia ** | |
| No/yes | 64/12 (84.2%/15.8%) |
| Systemic treatment | |
| No/yes | 42/34 (55.3%/44.7%) |
| Length of in-patient treatment | |
| Median (range) | 7.5 days (1–78) |
| WHO clinical progression scale | |
| Mild/moderate | |
| I | 54 (71.1%) |
| II | 13 (17.1%) |
| Severe/death | |
| III | 6 (7.9%) |
| IV | 3 (3.9%) |
| Tube 1 Exhausted T Cells |
| CD45-PerCp-Cy5-5 |
| CD3-BV605 |
| CD4-BV786 |
| CD8-BUV395 |
| PD1(CD279)-BUV737 |
| CD39-PE-594 |
| TIM3(CD366)-BB515 |
| TIGIT-BV421 |
| Tube 2 Exhausted NK Cells |
| CD45-PerCp-Cy5-5 |
| CD56-BV786 |
| CD16-BUV395 |
| PD1(CD279)-BUV737 |
| LAG3(CD223)-PE-CF-594 |
| TIM3(CD366)-BB515 |
| TIGIT-BV421 |
| Tube 3 Monocytes |
| CD45-PerCp-Cy5-5 |
| CD11-BV605 |
| CD14-CF594 |
| CD16-BUV395 |
| CD66a-BV786 |
| CD200R1-BV421 |
| Cell Surface Markers | Controls | COVID-19 | p-Value |
|---|---|---|---|
| CD3+CD45+ | 69.3 (49.1–78.9) | 70.1 (0.9–93.7) | =0.78 |
| CD3+CD4+CD45+ | 64.2 (1.3–89.4) | 69.5 (0.6–96) | =0.25 |
| CD8+CD3+CD45+ | 26.4 (14.8–79.1) | 19.1 (3.1–66) | =0.011 * |
| CD56+CD45+ | 89.8 (77.7–93.6) | 65.5 (17.8–99) | <0.0001 * |
| CD16+CD45+ | 25.7 (12.6–37.9) | 21.7 (0.77–51.3) | =0.85 |
| CD56+CD16+CD45+ | 96.3 (89.5–98.6) | 89.1 (58.2–98.1) | <0.0001 * |
| CD39+CD3+CD45+ | 1.7 (0.57–6.4) | 5.9 (0.08–40.1) | =0.0004 * |
| CD39+CD4+CD3+CD45+ | 2.5 (0.19–26.1) | 5.8 (0–41.3) | =0.016 * |
| CD39+CD8+CD3+CD45+ | 0.82 (0.03–6.2) | 2.3 (0–100) | =0.0089 * |
| Cell Surface Markers | Controls | COVID-19 | p-Value |
|---|---|---|---|
| PD1+CD16+CD45+ | 9.8 (2.1–24.8) | 59.7 (29.3–94.7) | <0.0001 * |
| PD1+CD56+CD45+ | 9.1 (0.72–28.3) | 62.4 (35.9–94.4) | <0.0001 * |
| PD1+CD3+CD45+ | 20.8 (8.4–58.6) | 71.1 (40.1–96.7) | <0.0001 * |
| PD1+CD4+CD3+CD45+ | 17.2 (7.5–71.7) | 72.3 (38.1–98.5) | <0.0001 * |
| PD1+CD8+CD3+CD45+ | 27.5 (8.6–58.3) | 76.4 (0.6–98.2) | <0.0001 * |
| PD1+CD39+CD4+CD3+CD45+ | 30.7 (15.9–57.9) | 69.1 (0–99.4) | <0.0001 * |
| PD1+CD39+CD8+CD3+CD45+ | 37.3 (0–100) | 74.5 (0–100) | =0.0002 * |
| TIM3+CD16+CD45+ | 50.9 (31.5–73.1) | 66.6 (11.7–91.2) | =0.0066 * |
| TIM3+CD56+CD45+ | 28.6 (12.1–45.8) | 41.9 (11.3–67.9) | =0.0021 * |
| TIM3+CD3+CD45+ | 16.3 (4.9–32.3) | 28.2 (13.4–82.8) | =0.0001 * |
| TIM3+CD4+CD3+CD45+ | 17 (1.1–25.7) | 15.3 (4.4–84.9) | =0.64 |
| TIM3+CD8+CD3+CD45+ | 19.8 (1.1–32.4) | 21.9 (3.6–86,8) | =0.41 |
| TIM3+CD39+CD4+CD3+CD45+ | 27 (5.3–51.5) | 14.9 (0–80.7) | =0.0088 * |
| TIM3+CD39+CD8+CD3+CD45+ | 45.3 (15.6–57.1) | 25.8 (0–96.7) | =0.060 |
| LAG3+CD16+CD45+ | 2.5 (0.9–16.8) | 16.8 (2.1–59.6) | <0.0001 * |
| LAG3+CD56+CD45+ | 2.7 (0.54–13.9) | 10.9 (0.41–51.8) | =0.0004 * |
| Parameter | Univariable Analysis WHO clinical progression scale (class III and IV) | Multivariable Analysis WHO clinical progression scale (class III and IV) |
| LDH | AUC 0.79, p < 0.0001 Criterion: >252, Youden index: 0.41 | Did not remain in the model |
| C-reactive protein | AUC 0.90, p < 0.0001 Criterion: >39.8, Youden index: 0.78 | Did not remain in the model |
| Age | AUC 0.74, p = 0.0009 Criterion: >63, Youden index: 0.41 | Did not remain in the model |
| CD39+CD45+ | AUC 0.71, p = 0.041 Criterion: >30.4, Youden index: 0.41 | OR 51.4, 95% CI 1.5 to 1763 p = 0.029 |
| TIM3+CD39+CD4+CD3+CD45+ | AUC 0.73, p = 0.024 Criterion: >18.8, Youden index: 0.54 | OR 22.6, 95% CI 1.8 to 277 p = 0.015 |
| TIM3+CD39+CD8+CD3+CD45+ | AUC 0.73, p = 0.020 Criterion: >39, Youden index: 0.47 | Did not remain in the model |
| Cell Surface Markers | Controls | COVID-19 | p-Value |
|---|---|---|---|
| TIGIT+CD16+CD45+ | 38.1 (13.5–68.7) | 73.4 (13.2–96.1) | <0.0001 |
| TIGIT+CD56+CD45+ | 26 (16.1–55) | 48.1 (5.3–84) | =0.0009 * |
| TIGIT+CD3+CD45+ | 68.9 (3.8–91.9) | 83 (19.2–97.9) | =0.10 |
| TIGIT+CD4+CD3+CD45+ | 94.3 (19.1–99.6) | 93.3 (10.6–99.7) | =0.47 |
| TIGIT+CD8+CD3+CD45+ | 50.7 (12–88) | 49 (11.5–90.8) | =0.95 |
| TIGIT+CD39+CD4+CD3+CD45+ | 99.6 (67–100) | 92.1 (0–100) | =0.0017 * |
| TIGIT+CD39+CD8+CD3+CD45+ | 68.4 (43.8–100) | 75 (0–100) | =0.30 |
| CD200R+CD16+CD45+ | 1.9 (0.9–19.3) | 14.4 (0.45–90.8) | <0.0001 * |
| CD200R+CD11c+CD45+ | 12.2 (2.8–34) | 30.3 (1.4–93.7) | =0.0005 * |
| CD200R+CD14+CD45+ | 20 (3.1–48.2) | 44.2 (0–92.3) | =0.0003 * |
| CD200R+CD66+CD45+ | 19.5 (2.5–48.2) | 67.2 (1.1–95.2) | <0.0001 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambichler, T.; Rüth, J.; Goesmann, S.; Höxtermann, S.; Skrygan, M.; Susok, L.; Becker, J.C.; Overheu, O.; Schmidt, W.; Reinacher-Schick, A. A Prospective Study Investigating Immune Checkpoint Molecule and CD39 Expression on Peripheral Blood Cells for the Prognostication of COVID-19 Severity and Mortality. Viruses 2024, 16, 810. https://doi.org/10.3390/v16050810
Gambichler T, Rüth J, Goesmann S, Höxtermann S, Skrygan M, Susok L, Becker JC, Overheu O, Schmidt W, Reinacher-Schick A. A Prospective Study Investigating Immune Checkpoint Molecule and CD39 Expression on Peripheral Blood Cells for the Prognostication of COVID-19 Severity and Mortality. Viruses. 2024; 16(5):810. https://doi.org/10.3390/v16050810
Chicago/Turabian StyleGambichler, Thilo, Jonas Rüth, Silke Goesmann, Stefan Höxtermann, Marina Skrygan, Laura Susok, Jürgen C. Becker, Oliver Overheu, Wolfgang Schmidt, and Anke Reinacher-Schick. 2024. "A Prospective Study Investigating Immune Checkpoint Molecule and CD39 Expression on Peripheral Blood Cells for the Prognostication of COVID-19 Severity and Mortality" Viruses 16, no. 5: 810. https://doi.org/10.3390/v16050810
APA StyleGambichler, T., Rüth, J., Goesmann, S., Höxtermann, S., Skrygan, M., Susok, L., Becker, J. C., Overheu, O., Schmidt, W., & Reinacher-Schick, A. (2024). A Prospective Study Investigating Immune Checkpoint Molecule and CD39 Expression on Peripheral Blood Cells for the Prognostication of COVID-19 Severity and Mortality. Viruses, 16(5), 810. https://doi.org/10.3390/v16050810