
Epidemiological Analyses of the First Incursion of the Epizootic Hemorrhagic Disease Virus Serotype 8 in Tunisia, 2021–2022

 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Outbreak Dataset and Cluster Analysis
2.2. Environmental Risk Models
2.3. Evaluation of the Model
3. Results
3.1. Spatiotemporal and Cluster Analysis
3.2. Environmental Risk Models
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mertens, P.P.C.; Maan, S.; Samuel, A.; Attoui, H. Orbiviruses, Reoviridae, p 466–483. In Virus Taxonomy: Eighth Report of the International Committee on Taxonomy of Viruses; Elsevier/Academic Press: London, UK, 2005. [Google Scholar]
- Shope, R.E.; Macnamara, L.G.; Mangold, R. A Virus-Induced Epizootic Hemorrhagic Disease of the Virginia White-Tailed Deer (Odocoileus Virginianus). J. Exp. Med. 1960, 111, 155–170. [Google Scholar] [CrossRef]
- Rodrigues, T.C.S.; Viadanna, P.H.O.; Subramaniam, K.; Hawkins, I.K.; Jeon, A.B.; Loeb, J.C.; Krauer, J.M.C.; Lednicky, J.A.; Wisely, S.M.; Waltzek, T.B. Characterization of a Novel Reassortant Epizootic Hemorrhagic Disease Virus Serotype 6 Strain Isolated from Diseased White-Tailed Deer (Odocoileus Virginianus) on a Florida Farm. Viruses 2022, 14, 1012. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Cabello, L.; Utrilla-Trigo, S.; Lorenzo, G.; Ortego, J.; Calvo-Pinilla, E. Epizootic Hemorrhagic Disease Virus: Current Knowledge and Emerging Perspectives. Microorganisms 2023, 11, 1339. [Google Scholar] [CrossRef] [PubMed]
- Anthony, S.J.; Maan, S.; Maan, N.; Kgosana, L.; Bachanek-Bankowska, K.; Batten, C.; Darpel, K.E.; Sutton, G.; Attoui, H.; Mertens, P.P.C. Genetic and Phylogenetic Analysis of the Outer-Coat Proteins VP2 and VP5 of Epizootic Haemorrhagic Disease Virus (EHDV): Comparison of Genetic and Serological Data to Characterise the EHDV Serogroup. Virus Res. 2009, 145, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Ben Dhaou, S.; Sailleau, C.; Babay, B.; Viarouge, C.; Sghaier, S.; Zientara, S.; Hammami, S.; Bréard, E.; Ben Dhaou, S.; Sailleau, C.; et al. Molecular Characterisation of Epizootic Haemorrhagic Disease Virus Associated with a Tunisian Outbreak among Cattle in 2006. Acta Vet. Hung. 2016, 64, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Golender, N.; Khinich, Y.; Gorohov, A.; Abramovitz, I.; Bumbarov, V. Epizootic Hemorrhagic Disease Virus Serotype 6 Outbreak in Israeli Cattle in 2015. J. Vet. Diagn. Investig. 2017, 29, 885–888. [Google Scholar] [CrossRef]
- Golender, N.; Hoffmann, B. The Molecular Epidemiology of Epizootic Hemorrhagic Disease Viruses Identified in Israel between 2015 and 2023. Epidemiologia 2024, 5, 90–105. [Google Scholar] [CrossRef]
- Sghaier, S.; Hammami, S.; Goffredo, M.; Hammami, M.; Portanti, O.; Lorusso, A.; Savini, G.; Delécolle, J.C. New Species of the Genus Culicoides (Diptera Ceratopogonidae) for Tunisia, with Detection of Bluetongue Viruses in Vectors. Vet. Ital. 2017, 53, 357–366. [Google Scholar] [CrossRef]
- Ahmed, S.; Mahmoud, M.A.E.-F.; Viarouge, C.; Sailleau, C.; Zientara, S.; Breard, E. Presence of Bluetongue and Epizootic Hemorrhagic Disease Viruses in Egypt in 2016 and 2017. Infect. Genet. Evol. 2019, 73, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Hassine, T.B.; Sghaier, S.; Thabet, S.; Ammar, H.H.; Hammami, S. Role of Eco-Climatic Factors in the Distribution of Bluetongue in Endemic Areas in Tunisia. Open Vet. J. 2022, 12, 114–123. [Google Scholar] [CrossRef]
- Sghaier, S.; Sailleau, C.; Marcacci, M.; Thabet, S.; Curini, V.; Ben Hassine, T.; Teodori, L.; Portanti, O.; Hammami, S.; Jurisic, L.; et al. Epizootic Haemorrhagic Disease Virus Serotype 8 in Tunisia, 2021. Viruses 2023, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Thabet, S.; Sghaier, S.; Ben Hassine, T.; Slama, D.; Ben Osmane, R.; Ben Omrane, R.; Mouelhi, W.; Spedicato, M.; Leone, A.; Teodori, L.; et al. Characterization of Epizootic Hemorrhagic Disease Virus Serotype 8 in Naturally Infected Barbary Deer (Cervus Elaphus Barbarus) and Culicoides (Diptera: Ceratopogonidae) in Tunisia. Viruses 2023, 15, 1567. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, A.; Cappai, S.; Loi, F.; Pinna, L.; Ruiu, A.; Puggioni, G.; Guercio, A.; Purpari, G.; Vicari, D.; Sghaier, S.; et al. Epizootic Hemorrhagic Disease Virus Serotype 8, Italy, 2022. Emerg. Infect. Dis. 2023, 29, 1063–1065. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Wilson, A.; Barber, J.; Veronesi, E.; Mellor, P.; Venter, G.; Gubbins, S. Temperature Dependence of the Extrinsic Incubation Period of Orbiviruses in Culicoides Biting Midges. PLoS ONE 2011, 6, e27987. [Google Scholar] [CrossRef] [PubMed]
- Paweska, J.T.; Venter, G.J.; Hamblin, C. A Comparison of the Susceptibility of Culicoides Imicola and C. Bolitinos to Oral Infection with Eight Serotypes of Epizootic Haemorrhagic Disease Virus. Med. Vet. Entomol. 2005, 19, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Quaglia, M.; Foxi, C.; Satta, G.; Puggioni, G.; Bechere, R.; De Ascentis, M.; D’Alessio, S.G.; Spedicato, M.; Leone, A.; Pisciella, M.; et al. Culicoides Species Responsible for the Transmission of Epizootic Haemorrhagic Disease Virus (EHDV) Serotype 8 in Italy. Vet. Ital. 2023, 59, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Merrill, M.M.; Boughton, R.K.; Lollis, L.O.; Sayler, K.A.; Wisely, S.M. Epidemiology of Bluetongue Virus and Epizootic Hemorrhagic Disease Virus in Beef Cattle on a Ranch in South-Central Florida. Vector Borne Zoonotic Dis. 2019, 19, 752–757. [Google Scholar] [CrossRef]
- Lv, M.-N.; Zhu, J.-B.; Liao, S.-Q.; Yang, Z.-X.; Lin, X.-H.; Qi, N.-S.; Chen, Q.-L.; Wu, C.-Y.; Li, J.; Cai, H.-M.; et al. Seroprevalence of Epizootic Hemorrhagic Disease Virus in Guangdong Cattle Farms during 2013–2017, China. Viruses 2023, 15, 1263. [Google Scholar] [CrossRef]
- McGregor, B.L.; Erram, D.; Alto, B.W.; Lednicky, J.A.; Wisely, S.M.; Burkett-Cadena, N.D. Vector Competence of Florida Culicoides Insignis (Diptera: Ceratopogonidae) for Epizootic Hemorrhagic Disease Virus Serotype-2. Viruses 2021, 13, 410. [Google Scholar] [CrossRef]
- El Moustaid, F.; Thornton, Z.; Slamani, H.; Ryan, S.J.; Johnson, L.R. Predicting Temperature-Dependent Transmission Suitability of Bluetongue Virus in Livestock. Parasites Vectors 2021, 14, 382. [Google Scholar] [CrossRef]
- Anselin, L. Local Indicators of Spatial Association—LISA. Geogr. Anal. 1995, 27, 93–115. [Google Scholar] [CrossRef]
- Barbosa, A.M. fuzzySim: Applying Fuzzy Logic to Binary Similarity Indices in Ecology. Methods Ecol. Evol. 2015, 6, 853–858. [Google Scholar] [CrossRef]
- Leta, S.; Fetene, E.; Mulatu, T.; Amenu, K.; Jaleta, M.B.; Beyene, T.J.; Negussie, H.; Revie, C.W. Modeling the Global Distribution of Culicoides Imicola: An Ensemble Approach. Sci. Rep. 2019, 9, 14187. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, B.P.; Estrada, A.; Sutherland, C.; Barbosa, A.M. Projected Northward Shifts in Eastern Red-backed Salamanders Due to Changing Climate. Ecol. Evol. 2023, 13, e9999. [Google Scholar] [CrossRef]
- Real, R.; Barbosa, A.M.; Vargas, J.M. Obtaining Environmental Favourability Functions from Logistic Regression. Environ. Ecol. Stat. 2006, 13, 237–245. [Google Scholar] [CrossRef]
- Barbosa, A.M.; Real, R.; Muñoz, A.-R.; Brown, J.A. New Measures for Assessing Model Equilibrium and Prediction Mismatch in Species Distribution Models. Divers. Distrib. 2013, 19, 1333–1338. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the Accuracy of Species Distribution Models: Prevalence, Kappa and the True Skill Statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S. Goodness of Fit Tests for the Multiple Logistic Regression Model. Commun. Stat. Theory Methods 1980, 9, 1043–4069. [Google Scholar] [CrossRef]
- Miller, M.E.; Hui, S.L.; Tierney, W.M. Validation Techniques for Logistic Regression Models. Stat. Med. 1991, 10, 1213–1226. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S.; Sturdivant, R.X. Applied Logistic Regression; John Wiley & Sons: Hoboken, NJ, USA, 2013; ISBN 9780470582473. [Google Scholar]
- Noronha, L.E.; Cohnstaedt, L.W.; Richt, J.A.; Wilson, W.C. Perspectives on the Changing Landscape of Epizootic Hemorrhagic Disease Virus Control. Viruses 2021, 13, 2268. [Google Scholar] [CrossRef]
- Assefa, A.; Tibebu, A.; Bihon, A.; Dagnachew, A.; Muktar, Y. Ecological Niche Modeling Predicting the Potential Distribution of African Horse Sickness Virus from 2020 to 2060. Sci. Rep. 2022, 12, 1748. [Google Scholar] [CrossRef]
- Areias-Guerreiro, J.; Mira, A.; Barbosa, A.M. How Well Can Models Predict Changes in Species Distributions? A 13-Year-Old Otter Model Revisited. Hystrix It. J. Mamm. 2016, 27, 1. [Google Scholar] [CrossRef]
- Barbosa, A.M.; Real, R.; Vargas, J.M. Use of Coarse-Resolution Models of Species’ Distributions to Guide Local Conservation Inferences. Conserv. Biol. 2010, 24, 1378–1387. [Google Scholar] [CrossRef]
- Elith, J. Quantitative Methods for Modeling Species Habitat: Comparative Performance and an Application to Australian Plants. In Quantitative Methods for Conservation Biology; Springer: Berlin/Heidelberg, Germany, 2000; pp. 39–58. ISBN 978-0-387-95486-8. [Google Scholar]
- Wintle, B.A.; Elith, J.; Potts, J.M. Fauna Habitat Modelling and Mapping: A Review and Case Study in the Lower Hunter Central Coast Region of NSW. Austral Ecol. 2005, 30, 719–738. [Google Scholar] [CrossRef]
- Estrada, A.; Real, R. A Stepwise Assessment of Parsimony and Fuzzy Entropy in Species Distribution Modelling. Entropy 2021, 23, 1014. [Google Scholar] [CrossRef]
- García-Carrasco, J.-M.; Muñoz, A.-R.; Olivero, J.; Segura, M.; Real, R. Mapping the Risk for West Nile Virus Transmission, Africa. Emerg. Infect. Dis. 2022, 28, 777–785. [Google Scholar] [CrossRef]
- Peterson, A.T. Biogeography of Diseases: A Framework for Analysis. Naturwissenschaften 2008, 95, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, E.J.; Mellor, P.S.; Baylis, M. Using Climate Data to Map the Potential Distribution of Culicoides Imicola (Diptera: Ceratopogonidae) in Europe. Rev. Sci. Tech. OIE 2001, 20, 731–740. [Google Scholar] [CrossRef]
- Cuéllar, A.C.; Kjær, L.J.; Baum, A.; Stockmarr, A.; Skovgard, H.; Nielsen, S.A.; Andersson, M.G.; Lindström, A.; Chirico, J.; Lühken, R.; et al. Modelling the Monthly Abundance of Culicoides Biting Midges in Nine European Countries Using Random Forests Machine Learning. Parasit. Vectors 2020, 13, 194. [Google Scholar] [CrossRef]
- Leta, S.; Fetene, E.; Mulatu, T.; Amenu, K.; Jaleta, M.B.; Beyene, T.J.; Negussie, H.; Kriticos, D.; Revie, C.W. Updating the Global Occurrence of Culicoides Imicola, a Vector for Emerging Viral Diseases. Sci. Data 2019, 6, 185. [Google Scholar] [CrossRef]
- Guichard, S.; Guis, H.; Tran, A.; Garros, C.; Balenghien, T.; Kriticos, D.J. Worldwide Niche and Future Potential Distribution of Culicoides Imicola, a Major Vector of Bluetongue and African Horse Sickness Viruses. PLoS ONE 2014, 9, e112491. [Google Scholar] [CrossRef]
- Thameur, B.H.; Soufiène, S.; Haj Ammar, H.; Hammami, S. Spatial Distribution and Habitat Selection of Culicoides Imicola: The Potential Vector of Bluetongue Virus in Tunisia. Onderstepoort J. Vet. Res. 2021, 88, e1–e9. [Google Scholar] [CrossRef]
- Sailleau, C.; Zanella, G.; Breard, E.; Viarouge, C.; Desprat, A.; Vitour, D.; Adam, M.; Lasne, L.; Martrenchar, A.; Bakkali-Kassimi, L.; et al. Co-Circulation of Bluetongue and Epizootic Haemorrhagic Disease Viruses in Cattle in Reunion Island. Vet. Microbiol. 2012, 155, 191–197. [Google Scholar] [CrossRef]
- Viarouge, C.; Lancelot, R.; Rives, G.; Bréard, E.; Miller, M.; Baudrimont, X.; Doceul, V.; Vitour, D.; Zientara, S.; Sailleau, C. Identification of Bluetongue Virus and Epizootic Hemorrhagic Disease Virus Serotypes in French Guiana in 2011 and 2012. Vet. Microbiol. 2014, 174, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Kedmi, M.; Levi, S.; Galon, N.; Bomborov, V.; Yadin, H.; Batten, C.; Klement, E. No Evidence for Involvement of Sheep in the Epidemiology of Cattle Virulent Epizootic Hemorrhagic Disease Virus. Vet. Microbiol. 2011, 148, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Boyer, T.C.; Ward, M.P.; Singer, R.S. Climate, Landscape, and the Risk of Orbivirus Exposure in Cattle in Illinois and Western Indiana. Am. J. Trop. Med. Hyg. 2010, 83, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Kedmi, M.; Van Straten, M.; Ezra, E.; Galon, N.; Klement, E. Assessment of the Productivity Effects Associated with Epizootic Hemorrhagic Disease in Dairy Herds. J. Dairy. Sci. 2010, 93, 2486–2495. [Google Scholar] [CrossRef] [PubMed]
- Stevens, G.; McCluskey, B.; King, A.; O’Hearn, E.; Mayr, G. Review of the 2012 Epizootic Hemorrhagic Disease Outbreak in Domestic Ruminants in the United States. PLoS ONE 2015, 10, e0133359. [Google Scholar] [CrossRef]
- Christensen, S.A.; Ruder, M.G.; Williams, D.M.; Porter, W.F.; Stallknecht, D.E. The Role of Drought as a Determinant of Hemorrhagic Disease in the Eastern United States. Glob. Chang. Biol. 2020, 26, 3799–3808. [Google Scholar] [CrossRef] [PubMed]
- Dorak, S.J.; Varga, C.; Ruder, M.G.; Gronemeyer, P.; Rivera, N.A.; Dufford, D.R.; Skinner, D.J.; Roca, A.L.; Novakofski, J.; Mateus-Pinilla, N.E. Spatial Epidemiology of Hemorrhagic Disease in Illinois Wild White-Tailed Deer. Sci. Rep. 2022, 12, 6888. [Google Scholar] [CrossRef] [PubMed]
- Alba, A.; Casal, J.; Domingo, M. Possible Introduction of Bluetongue into the Balearic Islands, Spain, in 2000, via Air Streams. Vet. Rec. 2004, 155, 460–461. [Google Scholar] [CrossRef] [PubMed]
- Behar, A.; Friedgut, O.; Rotenberg, D.; Zalesky, O.; Izhaki, O.; Yulzary, A.; Rot, A.; Wolkomirsky, R.; Zamir, L.; Hmd, F.; et al. Insights on Transmission, Spread, and Possible Endemization of Selected Arboviruses in Israel—Interim Results from Five-Year Surveillance. Vet. Sci. 2022, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Rajko-Nenow, P.; Howson, E.L.A.; Clark, D.; Hilton, N.; Ambagala, A.; Svitek, N.; Flannery, J.; Batten, C. Development of a Novel Loop Mediated Isothermal Amplification Assay (LAMP) for the Rapid Detection of Epizootic Haemorrhagic Disease Virus. Viruses 2021, 13, 2187. [Google Scholar] [CrossRef]
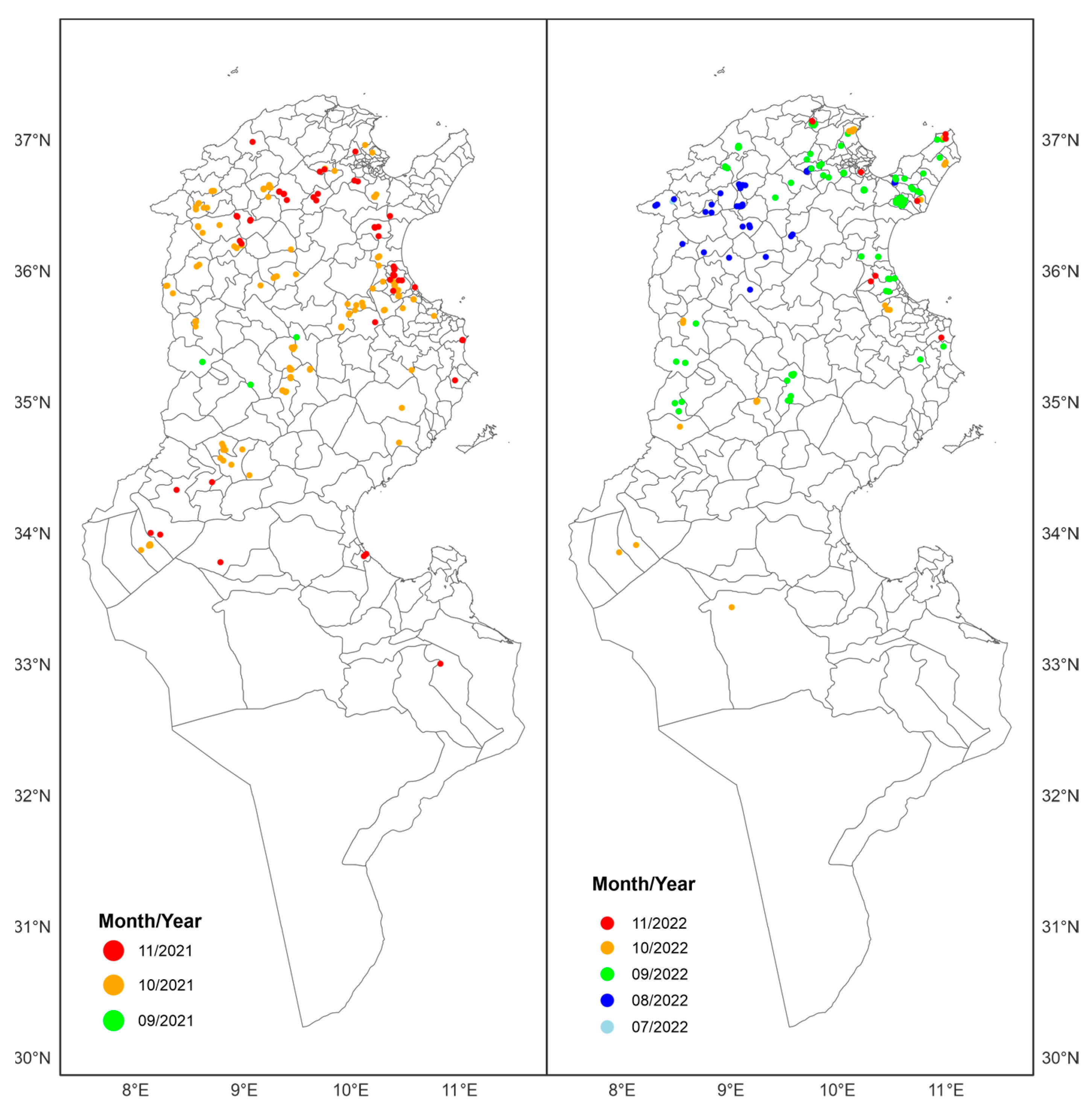

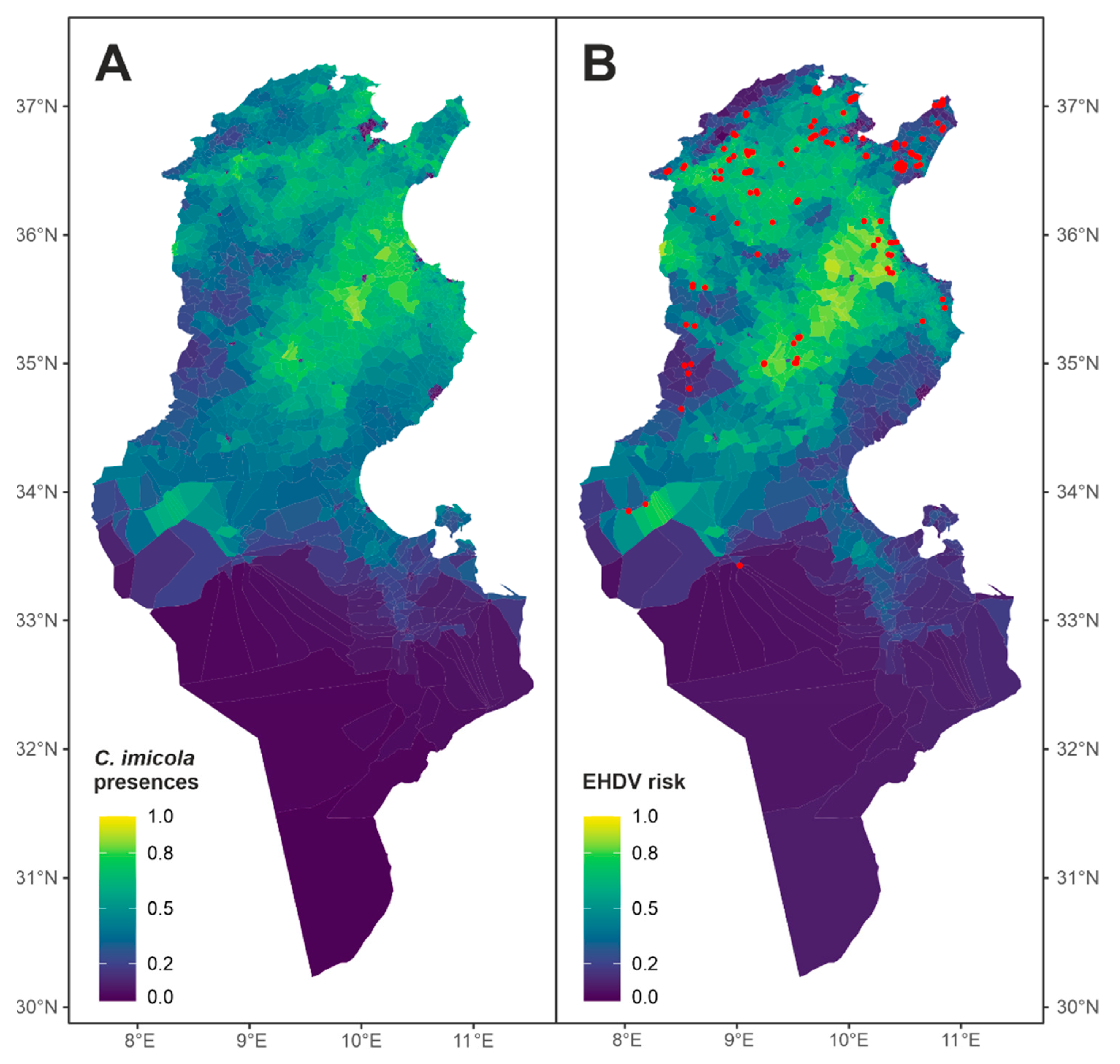
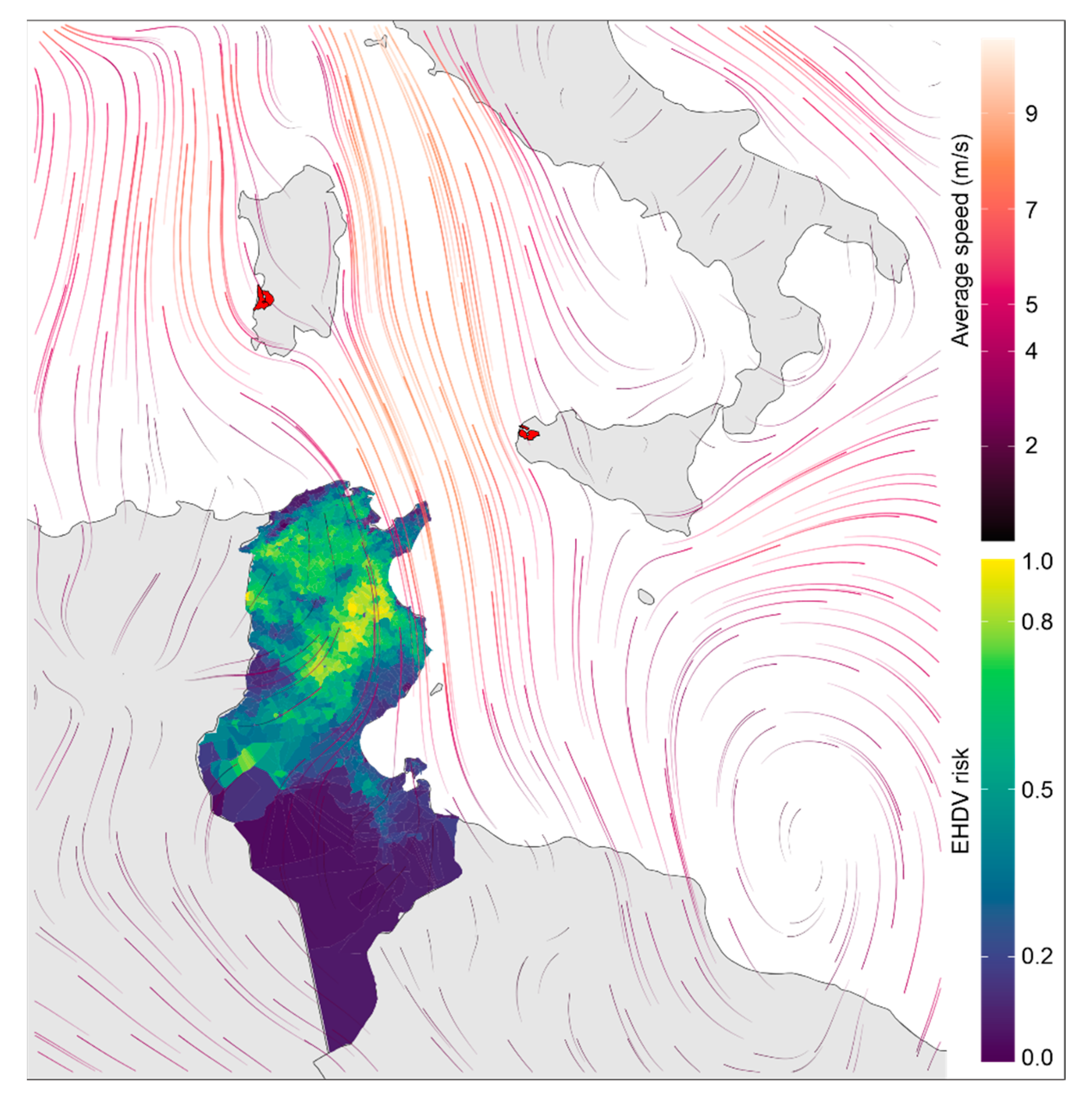

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Variable | Code |
|---|---|---|
| Topography | Altitude 1 | alt |
| Slope 1 | slope | |
| Climatic | Day land surface temperature mean (2021) 2 | dlst_mean |
| Night land surface temperature mean (2021) 2 | nlst_mean | |
| Land surface temperature day-night mean (2021) 2 | lst_dn_mean | |
| Land surface temperature difference Day-Night (2021) 2 | lst_dn_diff | |
| Water availability | Distance to rivers 3 | dist_river |
| Percentage of areas equipped for irrigation 4 | irrig | |
| Livestock | Cattledensity 5 | cattle |
| Sheepdensity 5 | sheep | |
| Anthropic | Population density 6 | dens_pop |
| Vegetation | Normalized Difference Vegetation Index mean (2021) 7 | NDVI_mean |
| Normalized Difference Vegetation Index minimum (2021) 7 | NDVI_min | |
| Normalized Difference Vegetation Index maximum (2021) 7 | NDVI_max | |
| Normalized Difference Vegetation Index difference between min and max (2021) 7 | NDVI_diff |
| Culicoides imicola Model | |||
|---|---|---|---|
| Estimate | Wald | Significance | |
| Intercept | −9.878 | 2.428 × 101 | 8.338 × 10−7 |
| Sheep | 1.367 × 10−4 | 3.100 | 7.828 × 10−2 |
| Irrigation | 1.365 | 5.889 | 1.523 × 10−2 |
| Dens_pob | −1.096 × 10−3 | 5.335 | 2.090 × 10−2 |
| NLST_mean | 4.143 × 10−1 | 1.014 × 101 | 1.451 × 10−3 |
| Dist_river | −9.285 × 10−6 | 4.005 | 4.538 × 10−2 |
| Epizootic hemorrhagic disease virus Model | |||
| Estimate | Wald | Significance | |
| Intercept | −1.594 × 101 | 2.471 × 101 | 6.679 × 10−7 |
| C. imicola_F | 4.124 | 2.974 × 101 | 4.932 × 10−8 |
| lst_dn_diff | 3.567 × 10−1 | 1.898 × 101 | 1.318 × 10−5 |
| Slope | −3.698 × 10−1 | 1.327 × 101 | 2.693 × 10−4 |
| NDVI_diff | 3.807 | 8.927 | 2.810 × 10−3 |
| nlst_mean | −2.725 × 10−1 | 3.855 | 4.958 × 10−2 |
| Culicoides imicola Model | EHDV Risk Model | |
|---|---|---|
| Sensitivity | 0.739 | 0.711 |
| Specificity | 0.607 | 0.637 |
| Under-prediction Rate | 0.010 | 0.023 |
| Over-prediction Rate | 0.957 | 0.907 |
| Kappa | 0.039 | 0.084 |
| CCR | 0.610 | 0.640 |
| TSS | 0.346 | 0.348 |
| AUC | 0.746 | 0.771 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Hassine, T.; García-Carrasco, J.-M.; Sghaier, S.; Thabet, S.; Lorusso, A.; Savini, G.; Hammami, S. Epidemiological Analyses of the First Incursion of the Epizootic Hemorrhagic Disease Virus Serotype 8 in Tunisia, 2021–2022. Viruses 2024, 16, 362. https://doi.org/10.3390/v16030362
Ben Hassine T, García-Carrasco J-M, Sghaier S, Thabet S, Lorusso A, Savini G, Hammami S. Epidemiological Analyses of the First Incursion of the Epizootic Hemorrhagic Disease Virus Serotype 8 in Tunisia, 2021–2022. Viruses. 2024; 16(3):362. https://doi.org/10.3390/v16030362
Chicago/Turabian StyleBen Hassine, Thameur, José-María García-Carrasco, Soufien Sghaier, Sarah Thabet, Alessio Lorusso, Giovanni Savini, and Salah Hammami. 2024. "Epidemiological Analyses of the First Incursion of the Epizootic Hemorrhagic Disease Virus Serotype 8 in Tunisia, 2021–2022" Viruses 16, no. 3: 362. https://doi.org/10.3390/v16030362
APA StyleBen Hassine, T., García-Carrasco, J.-M., Sghaier, S., Thabet, S., Lorusso, A., Savini, G., & Hammami, S. (2024). Epidemiological Analyses of the First Incursion of the Epizootic Hemorrhagic Disease Virus Serotype 8 in Tunisia, 2021–2022. Viruses, 16(3), 362. https://doi.org/10.3390/v16030362