
The Anti-SARS-CoV-2 IgG1 and IgG3 Antibody Isotypes with Limited Neutralizing Capacity against Omicron Elicited in a Latin Population a Switch toward IgG4 after Multiple Doses with the mRNA Pfizer–BioNTech Vaccine

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Antigen and Reagents
2.2. Study Cohorts
2.3. Ethical Statement
2.4. Detection of Anti-SARS-CoV-2 IgG Antibodies
2.5. Detection of SARS-CoV-2 IgG Subclasses
2.6. Neutralization Assay
2.7. Statistical Analysis
3. Results and Discussion
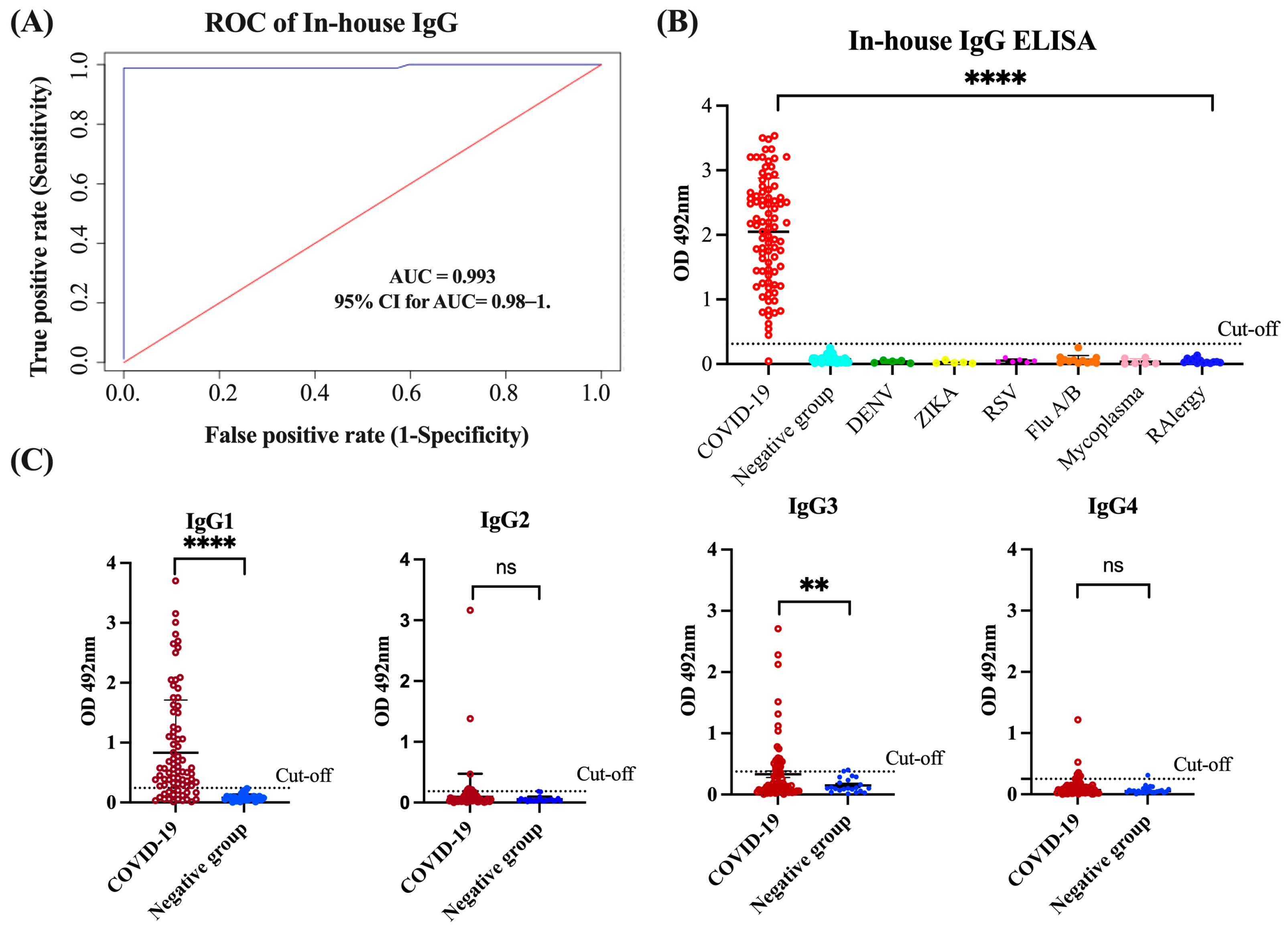
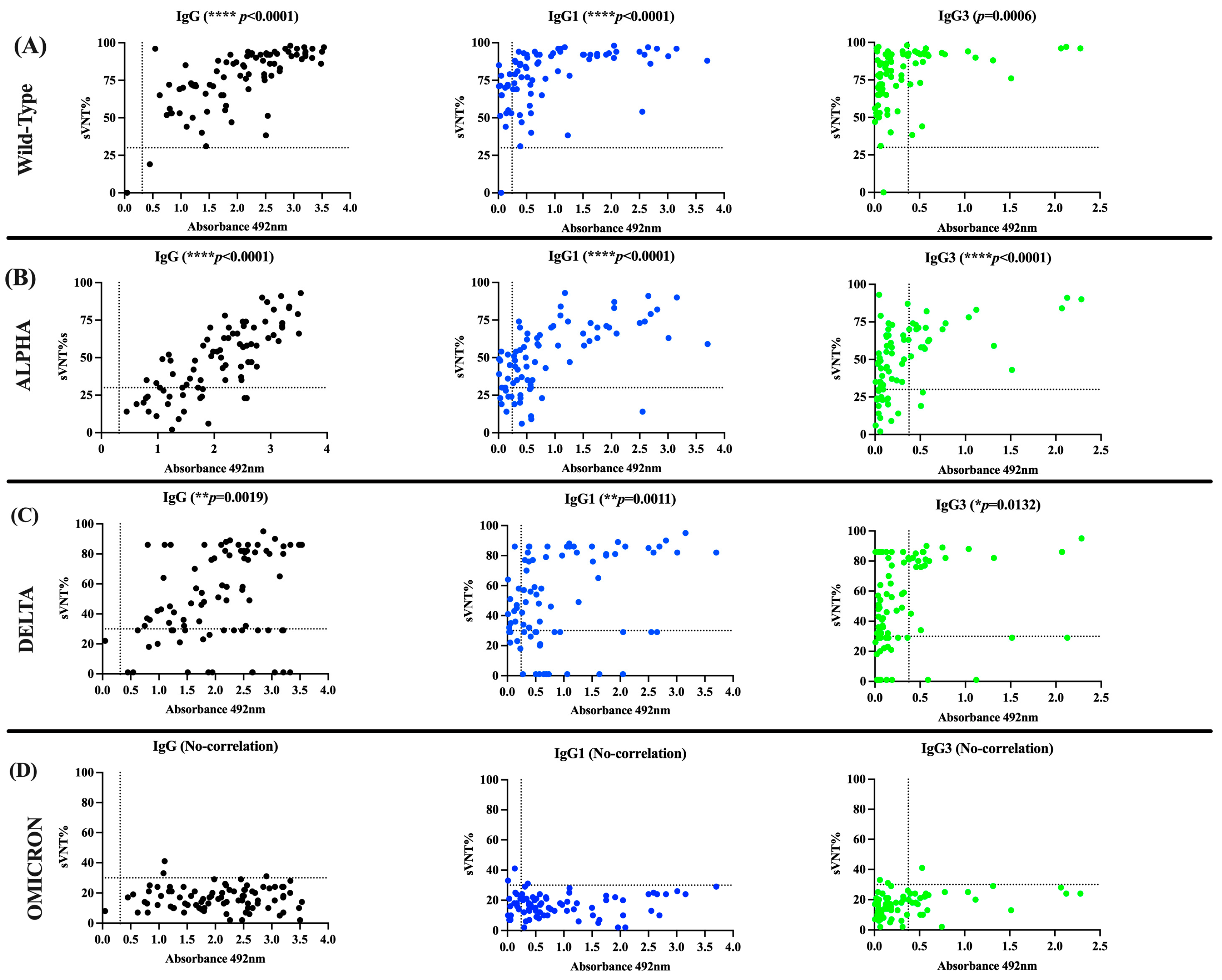
3.1. Assessment of In-House IgG and IgG Isotype ELISA Performance
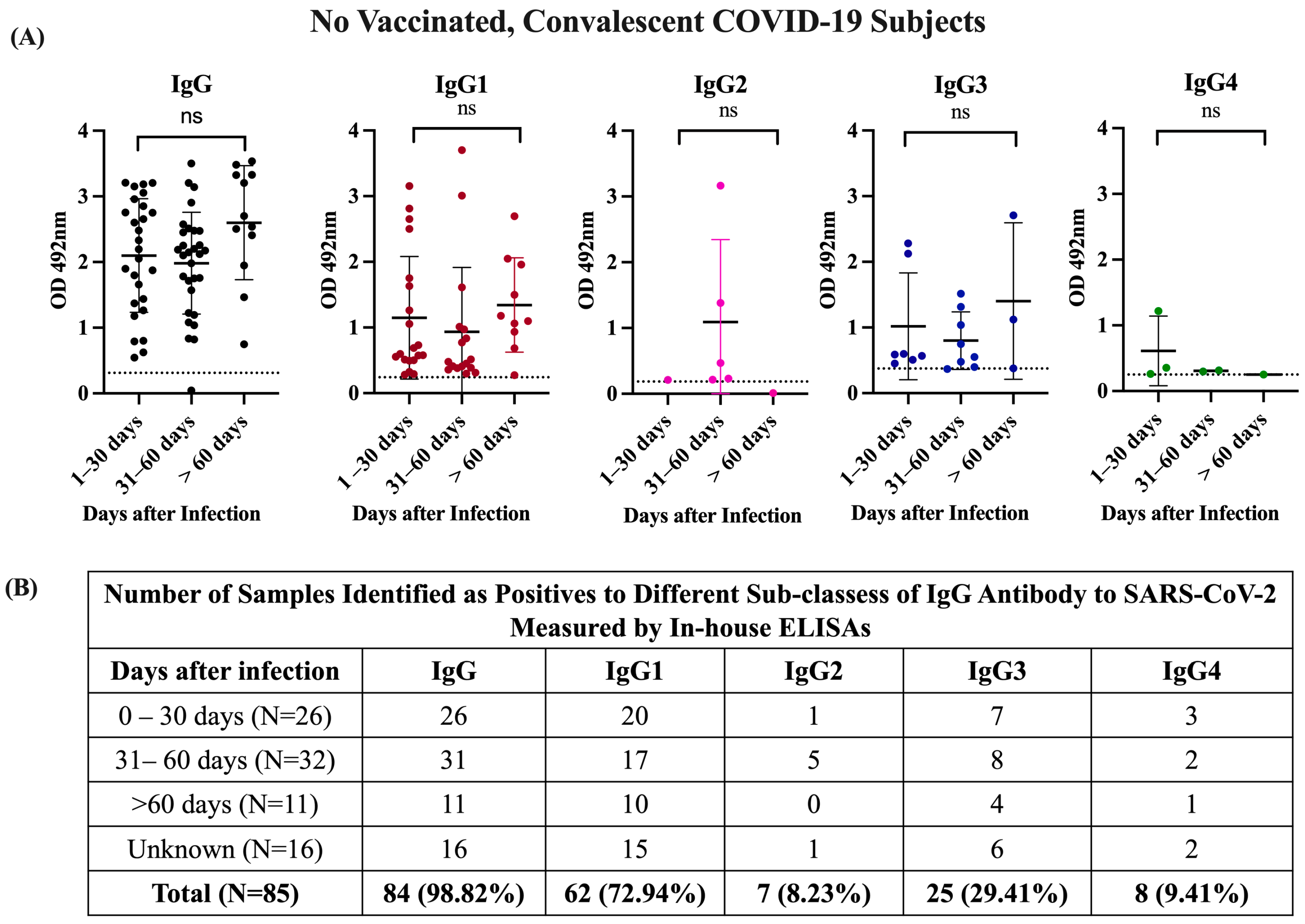
3.2. The Antibody Response in Unvaccinated Convalescent COVID-19 Subjects Is Dominated by IgG1 and IgG3 Isotypes, Which Neutralize the Wild-Type Strain and the Alpha and Delta VOCs but Are Poorly Effective against Omicron
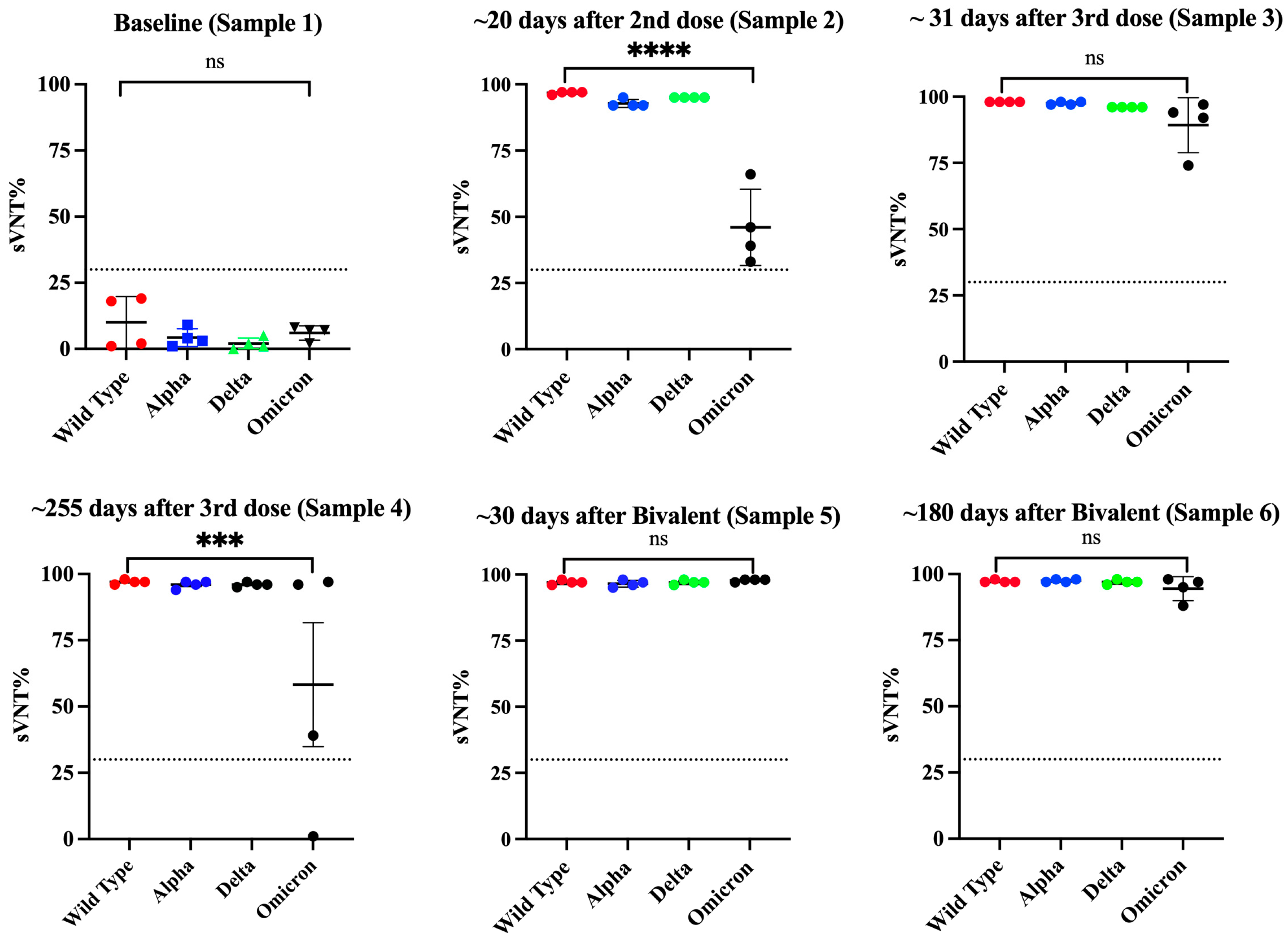
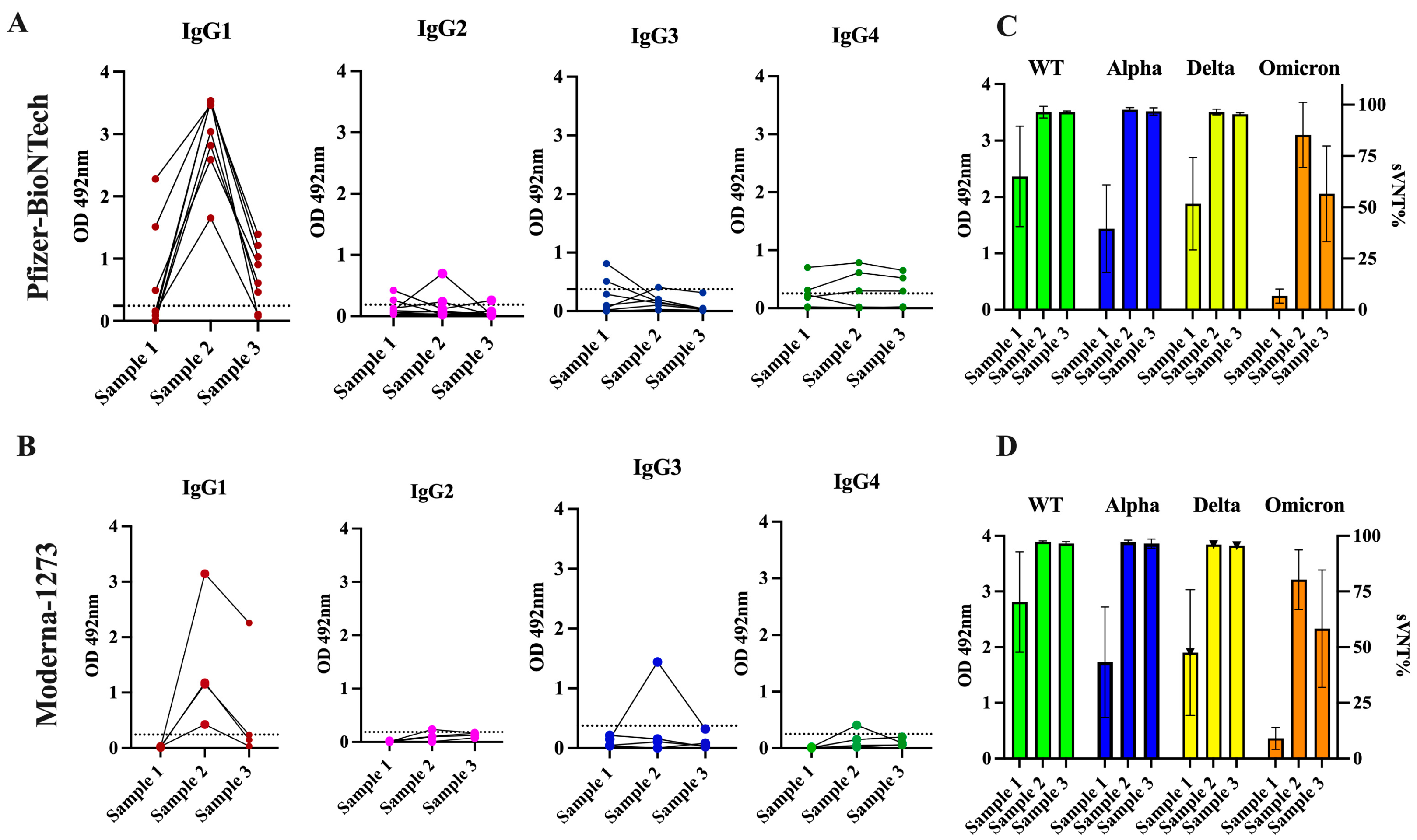
3.3. The Antibody Response in Previously Infected Subjects That Received Two Doses of the Pfizer–BioNTech or Moderna-1273 Vaccine Is Dominated by the IgG1 Isotype Which Has Potent Neutralizing Activity against the Alpha, Delta, and Omicron VOCs
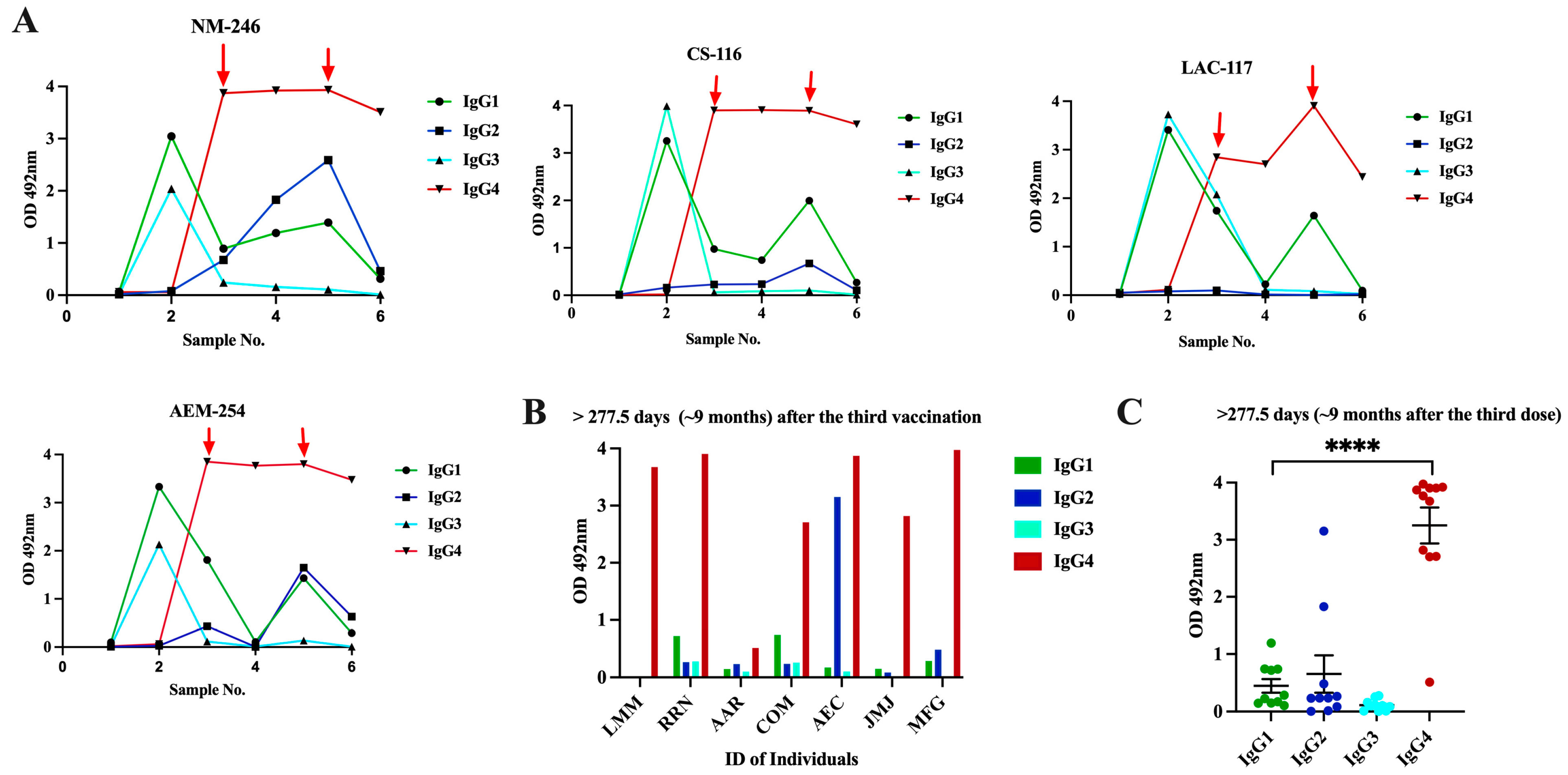
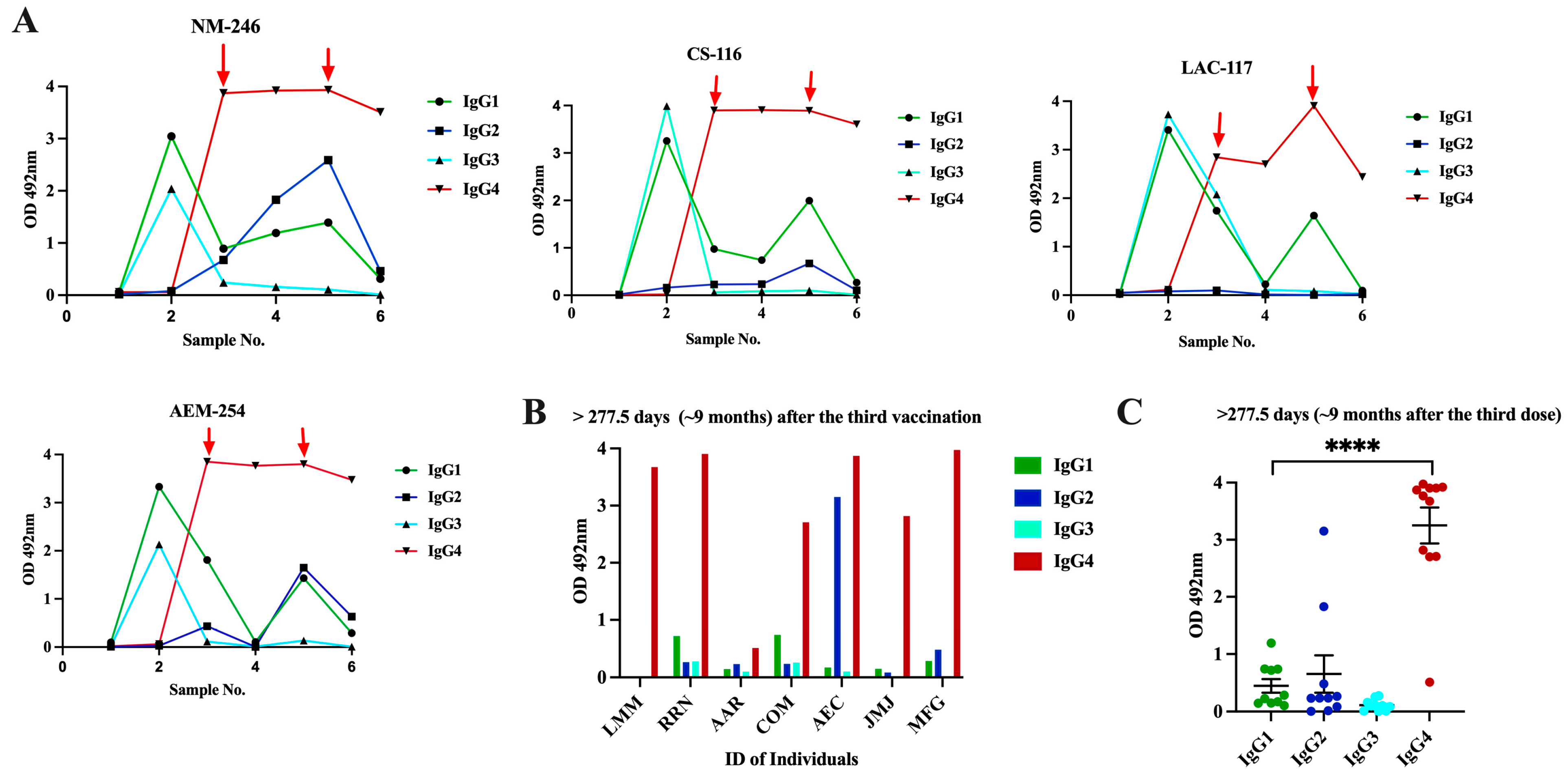
3.4. Class Switch toward IgG4 Occurs in Subjects That Receive Multiple Doses of Pfizer–BioNTech Vaccine, Which Is Sustained over the Time
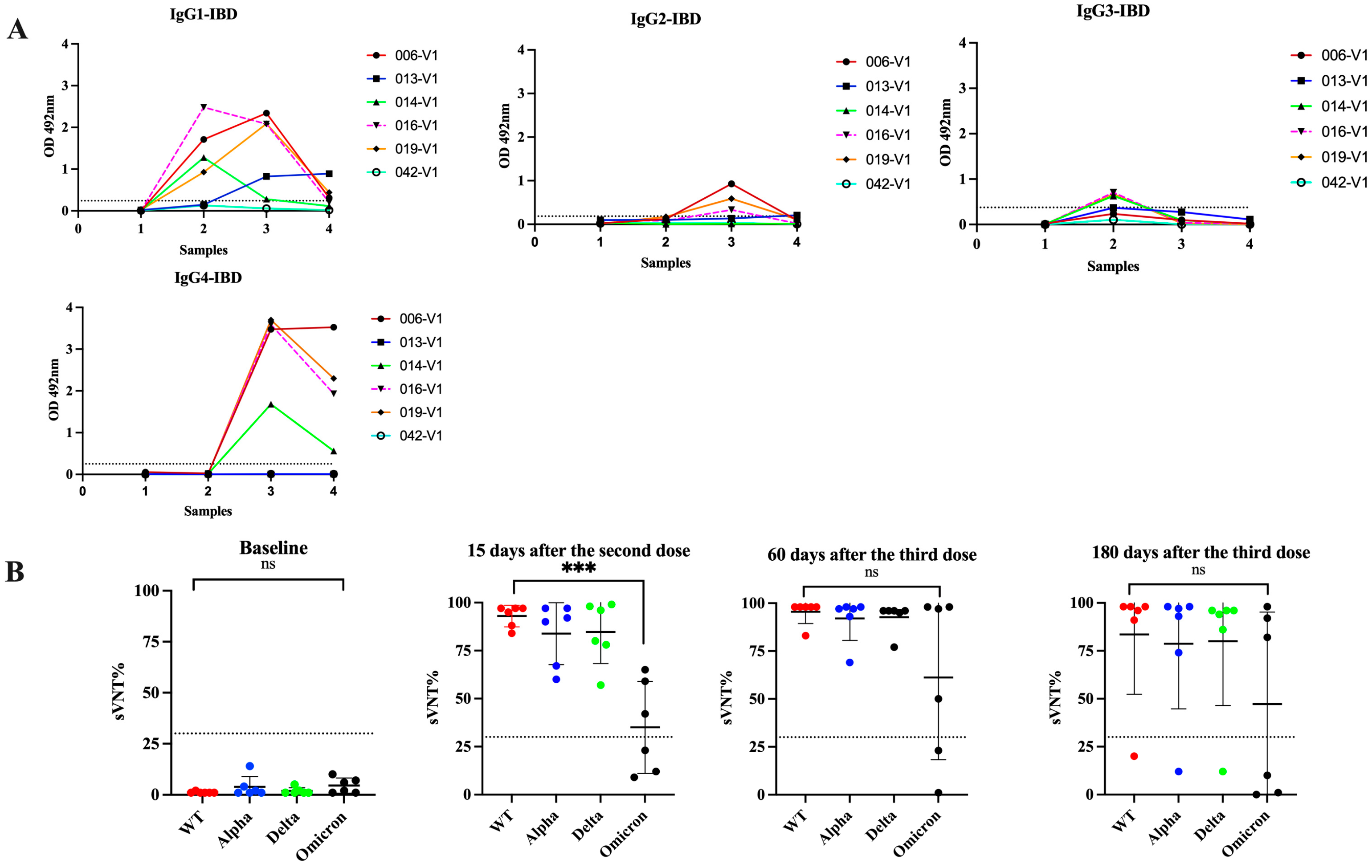
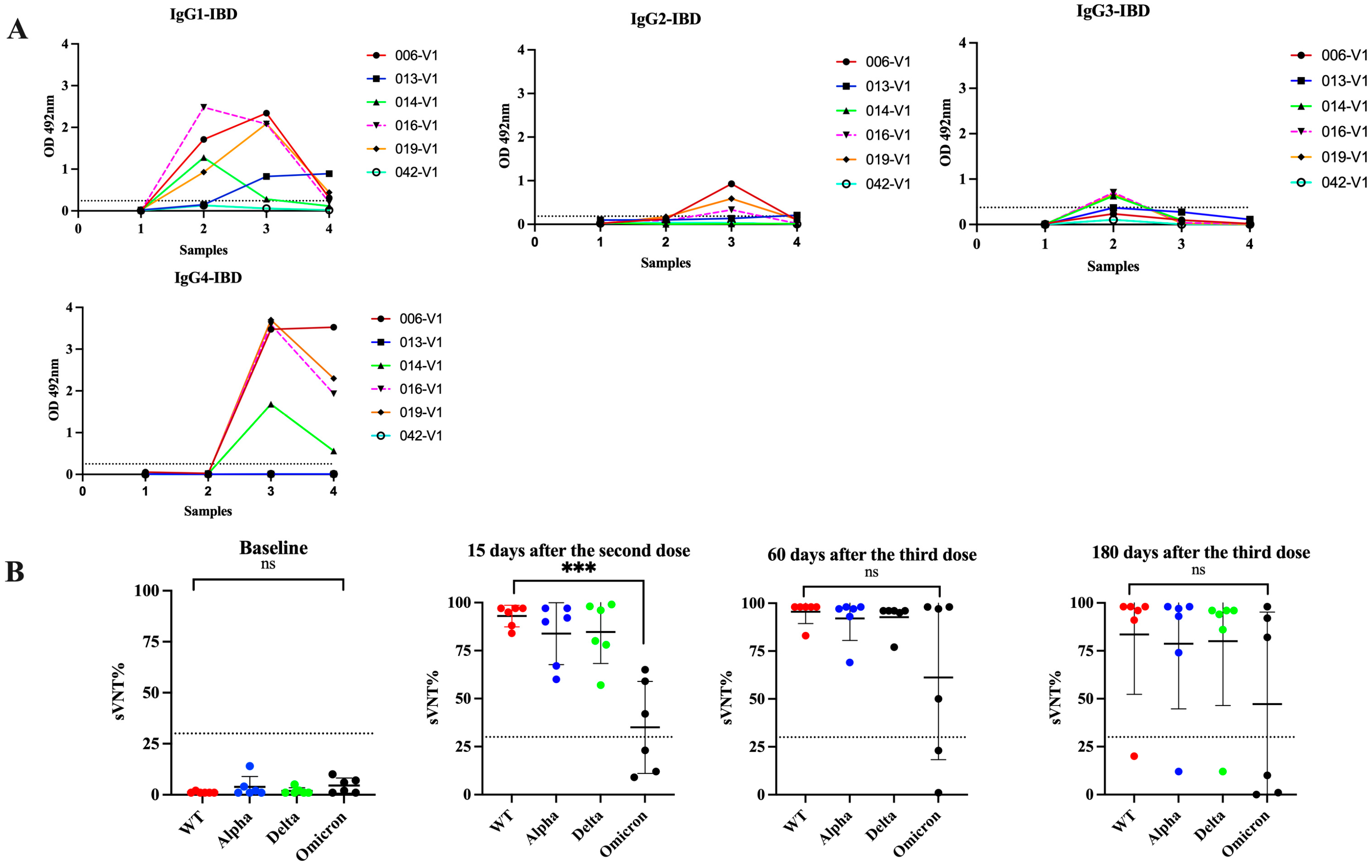
3.5. Switch in Class toward IgG4 Is Also Observed in Subjects with Inflammatory Bowel Disease and Received Multiple Vaccinations
4. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. WHO Coronavirus (COVID-19) Dashboard. 2022. Available online: https://covid19.who.int/ (accessed on 29 March 2022).
- Soto-Canetti, G.; Garcia, L.; Julia, A.E.; Gordian, E.I.; Bartolomei, J.A.; Camareno, N.; Rodriguez, J.F.; Montoya, M. Developing a Case Investigation and Contact-Tracing System in Puerto Rico, 2020. Am. J. Public Health 2022, 112, 223–226. [Google Scholar] [CrossRef]
- Amanat, F.; Stadlbauer, D.; Strohmeier, S.; Nguyen, T.H.O.; Chromikova, V.; McMahon, M.; Jiang, K.; Arunkumar, G.A.; Jurczyszak, D.; Polanco, J.; et al. A serological assay to detect SARS-CoV-2 seroconversion in humans. Nat. Med. 2020, 26, 1033–1036. [Google Scholar] [CrossRef]
- Lisboa Bastos, M.; Tavaziva, G.; Abidi, S.K.; Campbell, J.R.; Haraoui, L.P.; Johnston, J.C.; Lan, Z.; Law, S.; MacLean, E.; Trajman, A.; et al. Diagnostic accuracy of serological tests for covid-19: Systematic review and meta-analysis. BMJ 2020, 370, m2516. [Google Scholar] [CrossRef]
- Ramirez-Reveco, A.; Velasquez, G.; Aros, C.; Navarrete, G.; Villarroel-Espindola, F.; Navarrete, M.; Fica, A.; Plaza, A.; Castro, N.; Verdugo, C.; et al. Performance estimation of two in-house ELISA assays for COVID-19 surveillance through the combined detection of anti-SARS-CoV-2 IgA, IgM, and IgG immunoglobulin isotypes. PLoS ONE 2023, 18, e0270388. [Google Scholar] [CrossRef]
- Rungrojcharoenkit, K.; Suthangkornkul, R.; Utennam, D.; Buddhari, D.; Pinpaiboon, S.; Mongkolsirichaikul, D.; Fernandez, S.; Jones, A.R.; Cotrone, T.S.; Hunsawong, T. Standardization of in-house anti-IgG and IgA ELISAs for the detection of COVID-19. PLoS ONE 2023, 18, e0287107. [Google Scholar] [CrossRef]
- Sariol, C.A.A.; Pantoja, P.; Serrano-Collazo, C.; Rosa-Arocho, T.; Armina-Rodriguez, A.; Cruz, L.; Stone, E.T.T.; Arana, T.; Climent, C.; Latoni, G.; et al. Function Is More Reliable than Quantity to Follow Up the Humoral Response to the Receptor-Binding Domain of SARS-CoV-2-Spike Protein after Natural Infection or COVID-19 Vaccination. Viruses 2021, 13, 1972. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Marte, P.; Soto-Gonzalez, A.; Ramos-Tollinchi, L.; Torres-Jorge, S.; Ferre, M.; Rodriguez-Martino, E.; Torres, E.A.; Sariol, C.A. Inefficient Induction of Neutralizing Antibodies against SARS-CoV-2 Variants in Patients with Inflammatory Bowel Disease on Anti-Tumor Necrosis Factor Therapy after Receiving a Third mRNA Vaccine Dose. Vaccines 2022, 10, 1301. [Google Scholar] [CrossRef] [PubMed]
- Sariol, C.A.; Serrano-Collazo, C.; Ortiz, E.J.; Pantoja, P.; Cruz, L.; Arana, T.; Atehortua, D.; Pabon-Carrero, C.; Espino, A.M. Limited Impact of Delta Variant’s Mutations on the Effectiveness of Neutralization Conferred by Natural Infection or COVID-19 Vaccines in a Latino Population. Viruses 2021, 13, 2405. [Google Scholar] [CrossRef]
- Li, K.; Huang, B.; Wu, M.; Zhong, A.; Li, L.; Cai, Y.; Wang, Z.; Wu, L.; Zhu, M.; Li, J.; et al. Dynamic changes in anti-SARS-CoV-2 antibodies during SARS-CoV-2 infection and recovery from COVID-19. Nat. Commun. 2020, 11, 6044. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.M.; Algaissi, A.; Almahboub, S.A.; Alfaleh, M.A.; Abujamel, T.S.; Alamri, S.S.; Alluhaybi, K.A.; Hobani, H.I.; AlHarbi, R.H.; Alsulaiman, R.M.; et al. Early Humoral Response Correlates with Disease Severity and Outcomes in COVID-19 Patients. Viruses 2020, 12, 1390. [Google Scholar] [CrossRef]
- Trieu, M.C.; Bansal, A.; Madsen, A.; Zhou, F.; Saevik, M.; Vahokoski, J.; Brokstad, K.A.; Krammer, F.; Tondel, C.; Mohn, K.G.I.; et al. SARS-CoV-2-Specific Neutralizing Antibody Responses in Norwegian Health Care Workers after the First Wave of COVID-19 Pandemic: A Prospective Cohort Study. J. Infect. Dis. 2021, 223, 589–599. [Google Scholar] [CrossRef]
- Wang, P. Significance of IgA antibody testing for early detection of SARS-CoV-2. J. Med. Virol. 2021, 93, 1888–1889. [Google Scholar] [CrossRef]
- Yu, H.Q.; Sun, B.Q.; Fang, Z.F.; Zhao, J.C.; Liu, X.Y.; Li, Y.M.; Sun, X.Z.; Liang, H.F.; Zhong, B.; Huang, Z.F.; et al. Distinct features of SARS-CoV-2-specific IgA response in COVID-19 patients. Eur. Respir. J. 2020, 56, 2001526. [Google Scholar] [CrossRef] [PubMed]
- Xiang, T.; Liang, B.; Fang, Y.; Lu, S.; Li, S.; Wang, H.; Li, H.; Yang, X.; Shen, S.; Zhu, B.; et al. Declining Levels of Neutralizing Antibodies against SARS-CoV-2 in Convalescent COVID-19 Patients One Year Post Symptom Onset. Front. Immunol. 2021, 12, 708523. [Google Scholar] [CrossRef]
- Zhu, L.; Xu, X.; Zhu, B.; Guo, X.; Xu, K.; Song, C.; Fu, J.; Yu, H.; Kong, X.; Peng, J.; et al. Kinetics of SARS-CoV-2 Specific and Neutralizing Antibodies over Seven Months after Symptom Onset in COVID-19 Patients. Microbiol. Spectr. 2021, 9, e0059021. [Google Scholar] [CrossRef] [PubMed]
- Hantz, S. Biological diagnosis of Sars-CoV-2 infection: Strategies and interpretation of results. Rev. Francoph. Lab. 2020, 2020, 48–56. [Google Scholar] [CrossRef]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG subclasses and allotypes: From structure to effector functions. Front. Immunol. 2014, 5, 520. [Google Scholar] [CrossRef] [PubMed]
- Husain-Syed, F.; Vadasz, I.; Wilhelm, J.; Walmrath, H.D.; Seeger, W.; Birk, H.W.; Jennert, B.; Dietrich, H.; Herold, S.; Trauth, J.; et al. Immunoglobulin deficiency as an indicator of disease severity in patients with COVID-19. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 320, L590–L599. [Google Scholar] [CrossRef]
- Patil, H.P.; Rane, P.S.; Shrivastava, S.; Palkar, S.; Lalwani, S.; Mishra, A.C.; Arankalle, V.A. Antibody (IgA, IgG, and IgG Subtype) Responses to SARS-CoV-2 in Severe and Nonsevere COVID-19 Patients. Viral Immunol. 2021, 34, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Yates, J.L.; Ehrbar, D.J.; Hunt, D.T.; Girardin, R.C.; Dupuis, A.P., 2nd; Payne, A.F.; Sowizral, M.; Varney, S.; Kulas, K.E.; Demarest, V.L.; et al. Serological analysis reveals an imbalanced IgG subclass composition associated with COVID-19 disease severity. Cell Rep. Med. 2021, 2, 100329. [Google Scholar] [CrossRef]
- Kellner, C.; Otte, A.; Cappuzzello, E.; Klausz, K.; Peipp, M. Modulating cytotoxic effector functions by Fc engineering to improve cancer tharpy. Transfus. Med. Hemother. 2017, 44, 327–336. [Google Scholar] [CrossRef]
- Damelang, T.; Rogerson, S.J.; Kent, S.J.; Chung, A.W. Role of IgG3 in infections diseases. Trends Immunol. 2019, 40, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Tejedor Vaquero, S.; de Campos-Mata, L.; Ramada, J.M.; Diaz, P.; Navarro-Barriuso, J.; Ribas-Llaurado, C.; Rodrigo Melero, N.; Carolis, C.; Cerutti, A.; Gimeno, R.; et al. The mRNA-1273 Vaccine Induces Cross-Variant Antibody Responses to SARS-CoV-2 with Distinct Profiles in Individuals with or without Pre-Existing Immunity. Front. Immunol. 2021, 12, 737083. [Google Scholar] [CrossRef]
- Rahman, S.; Rahman, M.M.; Miah, M.; Begum, M.N.; Sarmin, M.; Mahfuz, M.; Hossain, M.E.; Rahman, M.Z.; Chisti, M.J.; Ahmed, T.; et al. COVID-19 reinfections among naturally infected and vaccinated individuals. Sci. Rep. 2022, 12, 1438. [Google Scholar] [CrossRef]
- Ren, X.; Zhou, J.; Guo, J.; Hao, C.; Zheng, M.; Zhang, R.; Huang, Q.; Yao, X.; Li, R.; Jin, Y. Reinfection in patients with COVID-19: A systematic review. Glob. Health Res. Policy 2022, 7, 12. [Google Scholar] [CrossRef]
- Fernandes, Q.; Inchakalody, V.P.; Merhi, M.; Mestiri, S.; Taib, N.; Moustafa Abo El-Ella, D.; Bedhiafi, T.; Raza, A.; Al-Zaidan, L.; Mohsen, M.O.; et al. Emerging COVID-19 variants and their impact on SARS-CoV-2 diagnosis, therapeutics and vaccines. Ann. Med. 2022, 54, 524–540. [Google Scholar] [CrossRef] [PubMed]
- Espino, A.M.; Pantoja, P.; Sariol, C.A. Validation and performance of a quantitative IgG assay for the screening of SARS-CoV-2 antibodies. bioRxv 2020. [Google Scholar] [CrossRef]
- Taylor, S.C.; Hurst, B.; Charlton, C.L.; Bailey, A.; Kanji, J.N.; McCarthy, M.K.; Morrison, T.E.; Huey, L.; Annen, K.; DomBourian, M.G.; et al. A New SARS-CoV-2 Dual-Purpose Serology Test: Highly Accurate Infection Tracing and Neutralizing Antibody Response Detection. J. Clin. Microbiol. 2021, 59, e02438-20. [Google Scholar] [CrossRef] [PubMed]
- Hosmer, D.W., Jr.; Lemeshow, S.; Sturdivant, R.X. Applied Logistic Regression; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Obermeier, P.; Muehlhans, S.; Hoppe, C.; Karsch, K.; Tief, F.; Seeber, L.; Chen, X.; Conrad, T.; Boettcher, S.; Diedrich, S.; et al. Enabling Precision Medicine with Digital Case Classification at the Point-of-Care. EBioMedicine 2016, 4, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Linnet, K. Evaluation of regression procedures for methods comparison studies. Clin. Chem. 1993, 39, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Wieckowska, B.; Kubiak, K.B.; Jozwiak, P.; Moryson, W.; Stawinska-Witoszynska, B. Cohen’s Kappa Coefficient as a Measure to Assess Classification Improvement following the Addition of a New Marker to a Regression Model. Int. J. Environ. Res. Public Health 2022, 19, 10213. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.R.; Koch, G.G. An application of hierarchical kappa-type statistics in the assessment of majority agreement among multiple observers. Biometrics 1977, 33, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Valdes-Fernandez, B.N.; Duconge, J.; Espino, A.M.; Ruano, G. Personalized health and the coronavirus vaccines-Do individual genetics matter? Bioessays 2021, 43, e2100087. [Google Scholar] [CrossRef] [PubMed]
- Isho, B.; Abe, K.T.; Zuo, M.; Jamal, A.J.; Rathod, B.; Wang, J.H.; Li, Z.; Chao, G.; Rojas, O.L.; Bang, Y.M.; et al. Persistence of serum and saliva antibody responses to SARS-CoV-2 spike antigens in COVID-19 patients. Sci. Immunol. 2020, 5, eabe5511. [Google Scholar] [CrossRef] [PubMed]
- Chia, W.N.; Tan, C.W.; Foo, R.; Kang, A.E.Z.; Peng, Y.; Sivalingam, V.; Tiu, C.; Ong, X.M.; Zhu, F.; Young, B.E.; et al. Serological differentiation between COVID-19 and SARS infections. Emerg. Microbes Infect. 2020, 9, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, B.A.; Oliveira, L.C.; Sabino, E.C.; Okay, T.S. SARS-CoV-2 and the COVID-19 disease: A mini review on diagnostic methods. Rev. Inst. Med. Trop. Sao Paulo 2020, 62, e44. [Google Scholar] [CrossRef] [PubMed]
- Karger, A.B.; Brien, J.D.; Christen, J.M.; Dhakal, S.; Kemp, T.J.; Klein, S.L.; Pinto, L.A.; Premkumar, L.; Roback, J.D.; Binder, R.A.; et al. The Serological Sciences Network (SeroNet) for COVID-19: Depth and Breadth of Serology Assays and Plans for Assay Harmonization. mSphere 2022, 7, e0019322. [Google Scholar] [CrossRef]
- Luo, H.; Jia, T.; Chen, J.; Zeng, S.; Qiu, Z.; Wu, S.; Li, X.; Lei, Y.; Wang, X.; Wu, W.; et al. The Characterization of Disease Severity Associated IgG Subclasses Response in COVID-19 Patients. Front. Immunol. 2021, 12, 632814. [Google Scholar] [CrossRef]
- Korobova, Z.R.; Zueva, E.V.; Arsentieva, N.A.; Batsunov, O.K.; Liubimova, N.E.; Khamitova, I.V.; Kuznetsova, R.N.; Rubinstein, A.A.; Savin, T.V.; Stanevich, O.V.; et al. Changes in Anti-SARS-CoV-2 IgG Subclasses over Time and in Association with Disease Severity. Viruses 2022, 14, 941. [Google Scholar] [CrossRef]
- Rispens, T.; Huijbers, M.G. The unique properties of IgG4 and its roles in health and disease. Nat. Rev. Immunol. 2023, 23, 763–778. [Google Scholar] [CrossRef]
- Santiago, G.A.; Flores, B.; Gonzalez, G.L.; Charriez, K.N.; Huertas, L.C.; Volkman, H.R.; Van Belleghem, S.M.; Rivera-Amill, V.; Adams, L.E.; Marzan, M.; et al. Genomic surveillance of SARS-CoV-2 in Puerto Rico enabled early detection and tracking of variants. Commun. Med. 2022, 2, 100. [Google Scholar] [CrossRef] [PubMed]
- Arevalo-Rodriguez, I.; Buitrago-Garcia, D.; Simancas-Racines, D.; Zambrano-Achig, P.; Del Campo, R.; Ciapponi, A.; Sued, O.; Martinez-Garcia, L.; Rutjes, A.; Low, N.; et al. False-negative results of initial RT-PCR assays for COVID-19: A systematic review. PLoS ONE 2020, 15, e0242958. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.; Yip, C.C.; To, K.K.; Tang, T.H.; Wong, S.C.; Leung, K.H.; Fung, A.Y.; Ng, A.C.; Zou, Z.; Tsoi, H.W.; et al. Improved Molecular Diagnosis of COVID-19 by the Novel, Highly Sensitive and Specific COVID-19-RdRp/Hel Real-Time Reverse Transcription-PCR Assay Validated In Vitro and with Clinical Specimens. J. Clin. Microbiol. 2020, 58, e00310-20. [Google Scholar] [CrossRef] [PubMed]
- Loeffelholz, M.J.; Tang, Y.W. Laboratory diagnosis of emerging human coronavirus infections-the state of the art. Emerg. Microbes Infect. 2020, 9, 747–756. [Google Scholar] [CrossRef]
- Tang, Y.W.; Schmitz, J.E.; Persing, D.H.; Stratton, C.W. Laboratory Diagnosis of COVID-19: Current Issues and Challenges. J. Clin. Microbiol. 2020, 58, e00512-20. [Google Scholar] [CrossRef] [PubMed]
- Giovanetti, M.; Benedetti, F.; Campisi, G.; Ciccozzi, A.; Fabris, S.; Ceccarelli, G.; Tambone, V.; Caruso, A.; Angeletti, S.; Zella, D.; et al. Evolution patterns of SARS-CoV-2: Snapshot on its genome variants. Biochem. Biophys. Res. Commun. 2021, 538, 88–91. [Google Scholar] [CrossRef]
- Roy, U. Comparative structural analyses of selected spike protein-RBD mutations in SARS-CoV-2 lineages. Immunol. Res. 2022, 70, 143–151. [Google Scholar] [CrossRef]
- Chakraborty, C.; Bhattacharya, M.; Sharma, A.R.; Mallik, B. Omicron (B.1.1.529)—A new heavily mutated variant: Mapped location and probable properties of its mutations with an emphasis on S-glycoprotein. Int. J. Biol. Macromol. 2022, 219, 980–997. [Google Scholar] [CrossRef]
- Verbeke, R.; Lentacker, I.; De Smedt, S.C.; Dewitte, H. The dawn of mRNA vaccines: The COVID-19 case. J. Control Release 2021, 333, 511–520. [Google Scholar] [CrossRef]
- De Greef, J.; Scohy, A.; Zech, F.; Aboubakar, F.; Pilette, C.; Gerard, L.; Pothen, L.; Yildiz, H.; Belkhir, L.; Yombi, J.C. Determinants of IgG antibodies kinetics after severe and critical COVID-19. J. Med. Virol. 2021, 93, 5416–5424. [Google Scholar] [CrossRef]
- Wei, J.; Stoesser, N.; Matthews, P.C.; Ayoubkhani, D.; Studley, R.; Bell, I.; Bell, J.I.; Newton, J.N.; Farrar, J.; Diamond, I.; et al. Antibody responses to SARS-CoV-2 vaccines in 45,965 adults from the general population of the United Kingdom. Nat. Microbiol. 2021, 6, 1140–1149. [Google Scholar] [CrossRef]
- Choe, P.G.; Hong, J.; Park, J.; Chang, E.; Kang, C.K.; Kim, N.J.; Lee, C.H.; Park, W.B.; Oh, M.D. Persistent Antibody Responses Up to 18 Months after Mild Severe Acute Respiratory Syndrome Coronavirus 2 Infection. J. Infect. Dis. 2022, 226, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Altarawneh, H.N.; Chemaitelly, H.; Hasan, M.R.; Ayoub, H.H.; Qassim, S.; AlMukdad, S.; Coyle, P.; Yassine, H.M.; Al-Khatib, H.A.; Benslimane, F.M.; et al. Protection against the Omicron Variant from Previous SARS-CoV-2 Infection. N. Engl. J. Med. 2022, 386, 1288–1290. [Google Scholar] [CrossRef] [PubMed]
- Accorsi, E.K.; Britton, A.; Fleming-Dutra, K.E.; Smith, Z.R.; Shang, N.; Derado, G.; Miller, J.; Schrag, S.J.; Verani, J.R. Association Between 3 Doses of mRNA COVID-19 Vaccine and Symptomatic Infection Caused by the SARS-CoV-2 Omicron and Delta Variants. JAMA 2022, 327, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kruger, N.; Schulz, S.; Cossmann, A.; Rocha, C.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Moldenhauer, A.S.; Winkler, M.S.; et al. The Omicron variant is highly resistant against antibody-mediated neutralization: Implications for control of the COVID-19 pandemic. Cell 2022, 185, 447–456.e11. [Google Scholar] [CrossRef]
- Subramanian, S.V.; Kumar, A. Increases in COVID-19 are unrelated to levels of vaccination across 68 countries and 2947 counties in the United States. Eur. J. Epidemiol. 2021, 36, 1237–1240. [Google Scholar] [CrossRef]
- Maslinska, M.; Dmowska-Chalaba, J.; Jakubaszek, M. The Role of IgG4 in Autoimmunity and Rheumatic Diseases. Front. Immunol. 2021, 12, 787422. [Google Scholar] [CrossRef]
- Irrgang, P.; Gerling, J.; Kocher, K.; Lapuente, D.; Steininger, P.; Habenicht, K.; Wytopil, M.; Beileke, S.; Schafer, S.; Zhong, J. Class switch towards non-inflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination. Sci. Immunol. 2022, 8, eade2798. [Google Scholar] [CrossRef] [PubMed]
- Fernandes-Siqueira, L.O.; Sousa, B.G.; Cleto, C.E.; Wermelinger, L.S.; Caetano, B.L.L.; Pacheco, A.R.; Costa, S.M.; Almeida, F.C.L.; Ferreira, G.C.; Salmon, D.; et al. IgA quantification as a good predictor of the neutralizing antibodies levels after vaccination against SARS-CoV-2. J. Clin. Virol. Plus 2022, 2, 100121. [Google Scholar] [CrossRef]
- Hale, M.; Netland, J.; Chen, Y.; Thouvenel, C.D.; Smith, K.N.; Rich, L.M.; Vanderwall, E.R.; Miranda, M.C.; Eggenberger, J.; Hao, L.; et al. IgM antibodies derived from memory B cells are potent cross-variant neutralizers of SARS-CoV-2. J. Exp. Med. 2022, 219, e20220849. [Google Scholar] [CrossRef]
- Kim, K.O.; Jang, B.I. Management of inflammatory bowel disease in the COVID-19 era. Intest. Res. 2022, 20, 3–10. [Google Scholar] [CrossRef]
- Alexander, J.L.; Kennedy, N.A.; Ibraheim, H.; Anandabaskaran, S.; Saifuddin, A.; Castro Seoane, R.; Liu, Z.; Nice, R.; Bewshea, C.; D’Mello, A.; et al. COVID-19 vaccine-induced antibody responses in immunosuppressed patients with inflammatory bowel disease (VIP): A multicentre, prospective, case-control study. Lancet Gastroenterol. Hepatol. 2022, 7, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C.; Schuurman, J. IgG4 breaking the rules. Immunology 2002, 105, 9–19. [Google Scholar] [CrossRef]
- Aalberse, R.C.; Stapel, S.O.; Schuurman, J.; Rispens, T. Immunoglobulin G4: An odd antibody. Clin. Exp. Allergy 2009, 39, 469–477. [Google Scholar] [CrossRef]
- Huijbers, M.G.; Plomp, J.J.; van der Maarel, S.M.; Verschuuren, J.J. IgG4-mediated autoimmune diseases: A niche of antibody-mediated disorders. Ann. N. Y. Acad. Sci. 2018, 1413, 92–103. [Google Scholar] [CrossRef]
- Karagiannis, P.; Gilbert, A.E.; Josephs, D.H.; Ali, N.; Dodev, T.; Saul, L.; Correa, I.; Roberts, L.; Beddowes, E.; Koers, A.; et al. IgG4 subclass antibodies impair antitumor immunity in melanoma. J. Clin. Investig. 2013, 123, 1457–1474. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, P.; Gilbert, A.E.; Nestle, F.O.; Karagiannis, S.N. IgG4 antibodies and cancer-associated inflammation: Insights into a novel mechanism of immune escape. OncoImmunology 2013, 2, e24889. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Kojima, M.; Takata, K.; Morito, T.; Asaoku, H.; Takeuchi, T.; Mizobuchi, K.; Fujihara, M.; Kuraoka, K.; Nakai, T.; et al. Systemic IgG4-related lymphadenopathy: A clinical and pathologic comparison to multicentric Castleman’s disease. Mod. Pathol. 2009, 22, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Zen, Y.; Inoue, D.; Kitao, A.; Onodera, M.; Abo, H.; Miyayama, S.; Gabata, T.; Matsui, O.; Nakanuma, Y. IgG4-related lung and pleural disease: A clinicopathologic study of 21 cases. Am. J. Surg. Pathol. 2009, 33, 1886–1893. [Google Scholar] [CrossRef]
- Stone, J.H.; Khosroshahi, A.; Hilgenberg, A.; Spooner, A.; Isselbacher, E.M.; Stone, J.R. IgG4-related systemic disease and lymphoplasmacytic aortitis. Arthritis Rheum. 2009, 60, 3139–3145. [Google Scholar] [CrossRef]
- Billeskov, R.; Beikzadeh, B.; Berzofsky, J.A. The effect of antigen dose on T cell-targeting vaccine outcome. Hum. Vaccin. Immunother. 2019, 15, 407–411. [Google Scholar] [CrossRef]
- Deaths by Vaccination Status in England. Office for National Statistics. Available online: https://wwwonsgovuk/peoplepopulationandcommunity/birthsdeathsandmarriages/deaths/datasets/deathsbyvaccinationstatusengland (accessed on 9 November 2023).
- Aarstad, J.; Kvitastein, O.A. Is There a Link between the 2021 COVID-19 Vaccination Uptake in Europe and 2022 Excess All-Cause Mortality? Asian Pac. J. Health Sci. 2023, 10, 25–31. [Google Scholar]
- Uversky, V.N.; Redwan, E.M.; Makis, W.; Rubio-Casillas, A. IgG4 Antibodies Induced by Repeated Vaccination May Generate Immune Tolerance to the SARS-CoV-2 Spike Protein. Vaccines 2023, 11, 991. [Google Scholar] [CrossRef] [PubMed]
- Shuai, H.; Chan, J.F.; Hu, B.; Chai, Y.; Yuen, T.T.; Yin, F.; Huang, X.; Yoon, C.; Hu, J.C.; Liu, H.; et al. Attenuated replication and pathogenicity of SARS-CoV-2 B.1.1.529 Omicron. Nature 2022, 603, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Wolter, N.; Jassat, W.; Walaza, S.; Welch, R.; Moultrie, H.; Groome, M.; Amoako, D.G.; Everatt, J.; Bhiman, J.N.; Scheepers, C.; et al. Early assessment of the clinical severity of the SARS-CoV-2 omicron variant in South Africa: A data linkage study. Lancet 2022, 399, 437–446. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort-1: SARS-CoV-2 infected; convalescent not vaccinated. | |
| Date of Collection | 26 April 2020 to 5 June 2020 |
| Number of specimens | 85 (31 sera and 54 plasma) |
| Time after infection (gap elapsed between the date of the confirmatory RT-qPCR test and the sampling date) | |
| Range | 0 to 139 days |
| Median | 35.5 days |
| Number specimens with 0–30 days after infection. | 26 |
| Median | 22 days |
| Number specimens with 31–60 days after infection | 32 |
| Median | 37.5 days |
| Number specimens with >60 days after infection. | 11 |
| Median | 84 days |
| Unknown | 16 |
| Cohort-5: Pre-Pandemic Samples | |
| Subjects with unknown health status | |
| Collection date | 2012 |
| Number included | 78 (Sera) |
| Other respiratory/viral infections | |
| Collection date | 2018 to 2019 |
| Number included | 47 (Sera) |
| Respiratory allergies | 13 |
| Zika virus | 5 |
| Dengue virus | 5 |
| Influenza A/B | 12 |
| Respiratory Syncytial Virus | 6 |
| Mycoplasma | 6 |
| Cohort-2: Serial samples from previously SARS-CoV-2 infected subjects that received two doses of mRNA vaccine (Pfizer-BioNTech or Moderna-1273). | |
| Number of subjects | 12 |
| Sex | 7 female and 5 males |
| Date of collection | 27 October 2020 to 20 September 2021 |
| Number of specimens | 36 |
| Sample 1 (Baseline) | Days after infection |
| Range | |
| Median | |
| Sample 2 | Days after the 2nd dose |
| Range | 15 to 32 days |
| Median | 21.5 |
| Sample 3 | Days after the 2nd dose |
| Range | 74 to 169 days |
| Median | 96 |
| Cohort-3a: Serial samples from no previous SARS-CoV-2 infected subjects that received multiple Pfizer-BioNTech vaccinations. | |
| Number of subjects | 4 |
| Sex | 3 female and 1 male |
| Date of collection | 10 August 2020 to10 August 2023 |
| Number of specimens | 26 |
| Sample 1 (baseline) | Previous 1st dose |
| Sample 2 | Days after the 2nd dose |
| Range | 19 to 35 days |
| Median | 20 |
| Sample 3 | Days after the 3rd dose |
| Range | 31 to 43 days |
| Median | 31 |
| Sample 4 | Days after the 3rd dose |
| Range | 180 to 420 days |
| Median | 255 |
| Sample 5 | Days after bivalent vaccine |
| Median | 30 days |
| Sample 6 | Days after bivalent vaccine |
| Range | 90–180 days |
| Median | 180 |
| Cohort-3b: Single sample from no previous SARS-CoV-2 infection multiple Pfizer-BioNTech vaccinations, collected ~2 years after the last dose. | |
| Number of subjects | 8 |
| Sex | 4 female and 4 males |
| Number of specimens | 8 |
| Date of sample collection | 3 August 2023 to 23 October 2023 |
| Date of last vaccination (3rd dose or bivalent) | 30 September 2021 to 28 December 2021 |
| Sample 1 | Days after the 3rd dose |
| Range | 351 to 723 days |
| Median | 634 |
| Cohort-4: Serial samples from no-previous SARS-CoV-2 infected subjects with inflammatory bowel disease (IBD) that received three doses of mRNA vaccine (Pfizer). | |
| Number of subjects | 6 |
| Sex | 3 female and 3 males. |
| Date of collection | 14 April 2021 to 22 July 2022 |
| Number of specimens | 24 |
| Sample 1 (baseline) | Days after 2nd dose |
| Range | 15 to 28 days |
| Median | 17 |
| Sample 2 | Days after 3rd dose |
| Median | 60 |
| Sample 3 | Days after 3rd dose |
| Median | 180 |
| Antibody Class/Subclass | C-PASS Neutralization Test | |||
|---|---|---|---|---|
| Total IgG | WT | Alpha | Delta | Omicron |
| 98.87 | 75.29 | 69.41 | 3.529 |
| 0.661 | 0.056 | 0.049 | 0.00057 |
| Substantial agreement | Slight agreement | Slight agreement | No agreement |
| IgG1 | ||||
| 76.47 | 72.94 | 57.64 | 24.70 |
| 0.050 | 0.268 | −0.038 | −0.016 |
| Slight agreement | Fair agreement | No agreement | No agreement |
| IgG2 | ||||
| 9.41 | 31.76 | 34.88 | -- |
| 0.002 | 0.032 | −0.014 | No computed |
| No agreement | Slight agreement | No agreement | -- |
| IgG3 | ||||
| 30.58 | 48.23 | 50.58 | 69.23 |
| 0.018 | 0.151 | 0.147 | 0.006 |
| Slight agreement | Slight agreement | Slight agreement | No agreement |
| IgG4 | ||||
| 14.11 | 30.588 | 35.29 | -- |
| 0.0064 | 0.011 | −0.026 | No computed |
| No agreement | Slight agreement | No agreement | -- |
| IgM | ||||
| 88.23 | 77.64 | 72.94 | 16.47 |
| 0.255 | 0.298 | 0.266 | 0.0078 |
| Fair agreement | Fair agreement | Fair agreement | No agreement |
| IgA | ||||
| 40.00 | 55.29 | 48.23 | 64.19 |
| 0.028 | 0.205 | 0.050 | −0.002 |
| Slight agreement | Fair agreement | Slight agreement | No agreement |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espino, A.M.; Armina-Rodriguez, A.; Alvarez, L.; Ocasio-Malavé, C.; Ramos-Nieves, R.; Rodriguez Martinó, E.I.; López-Marte, P.; Torres, E.A.; Sariol, C.A. The Anti-SARS-CoV-2 IgG1 and IgG3 Antibody Isotypes with Limited Neutralizing Capacity against Omicron Elicited in a Latin Population a Switch toward IgG4 after Multiple Doses with the mRNA Pfizer–BioNTech Vaccine. Viruses 2024, 16, 187. https://doi.org/10.3390/v16020187
Espino AM, Armina-Rodriguez A, Alvarez L, Ocasio-Malavé C, Ramos-Nieves R, Rodriguez Martinó EI, López-Marte P, Torres EA, Sariol CA. The Anti-SARS-CoV-2 IgG1 and IgG3 Antibody Isotypes with Limited Neutralizing Capacity against Omicron Elicited in a Latin Population a Switch toward IgG4 after Multiple Doses with the mRNA Pfizer–BioNTech Vaccine. Viruses. 2024; 16(2):187. https://doi.org/10.3390/v16020187
Chicago/Turabian StyleEspino, Ana M., Albersy Armina-Rodriguez, Laura Alvarez, Carlimar Ocasio-Malavé, Riseilly Ramos-Nieves, Esteban I. Rodriguez Martinó, Paola López-Marte, Esther A. Torres, and Carlos A. Sariol. 2024. "The Anti-SARS-CoV-2 IgG1 and IgG3 Antibody Isotypes with Limited Neutralizing Capacity against Omicron Elicited in a Latin Population a Switch toward IgG4 after Multiple Doses with the mRNA Pfizer–BioNTech Vaccine" Viruses 16, no. 2: 187. https://doi.org/10.3390/v16020187
APA StyleEspino, A. M., Armina-Rodriguez, A., Alvarez, L., Ocasio-Malavé, C., Ramos-Nieves, R., Rodriguez Martinó, E. I., López-Marte, P., Torres, E. A., & Sariol, C. A. (2024). The Anti-SARS-CoV-2 IgG1 and IgG3 Antibody Isotypes with Limited Neutralizing Capacity against Omicron Elicited in a Latin Population a Switch toward IgG4 after Multiple Doses with the mRNA Pfizer–BioNTech Vaccine. Viruses, 16(2), 187. https://doi.org/10.3390/v16020187