
Transcriptome Analysis of LLC-PK Cells Single or Coinfected with Porcine Epidemic Diarrhea Virus and Porcine Deltacoronavirus



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Antibodies and Reagents
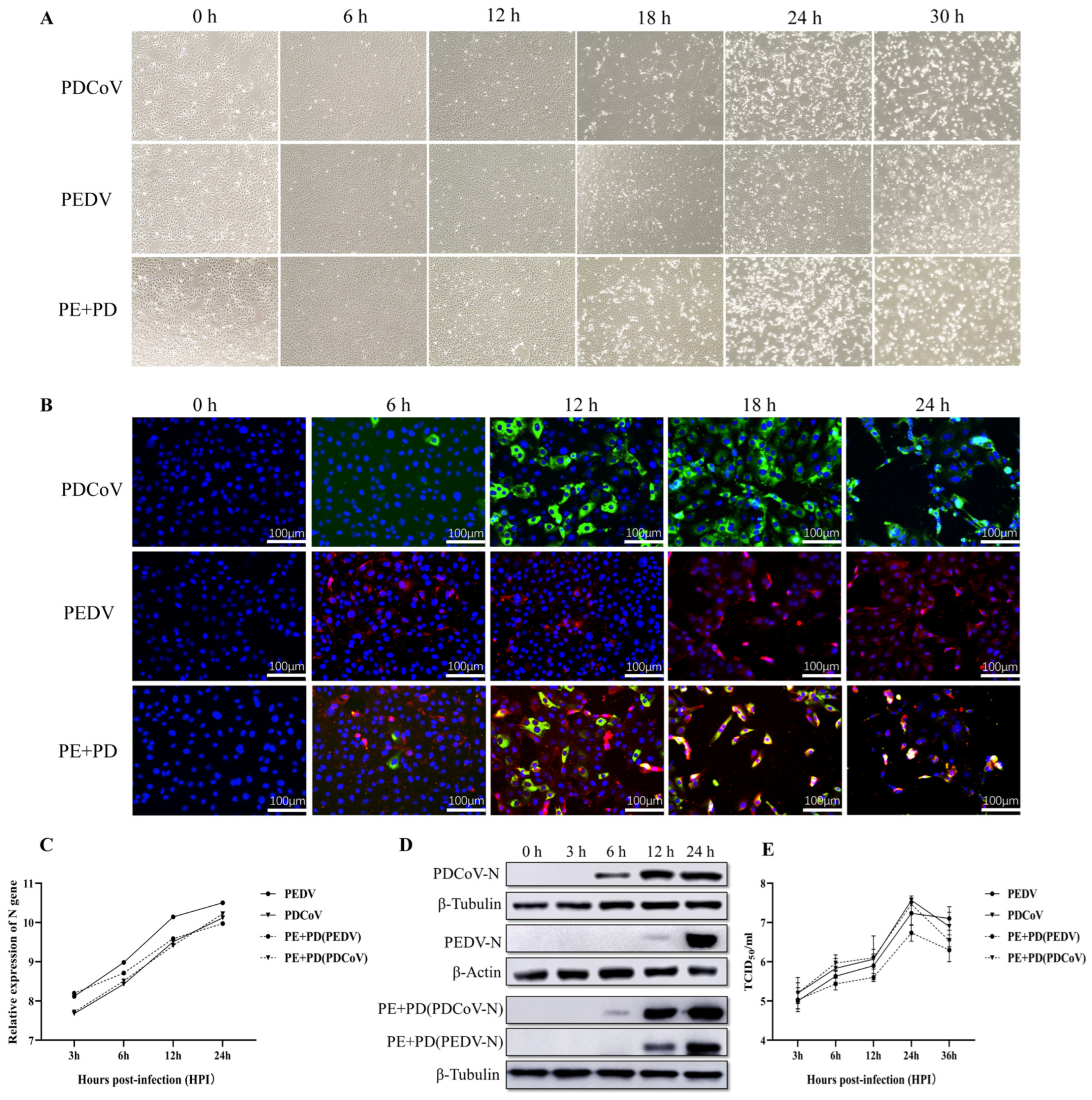
2.3. One-Step Growth Curve
2.4. Immunofluorescence Assay (IFA)
2.5. Viral Infection
2.6. RNA Extraction
2.7. Real-Time Quantitative PCR
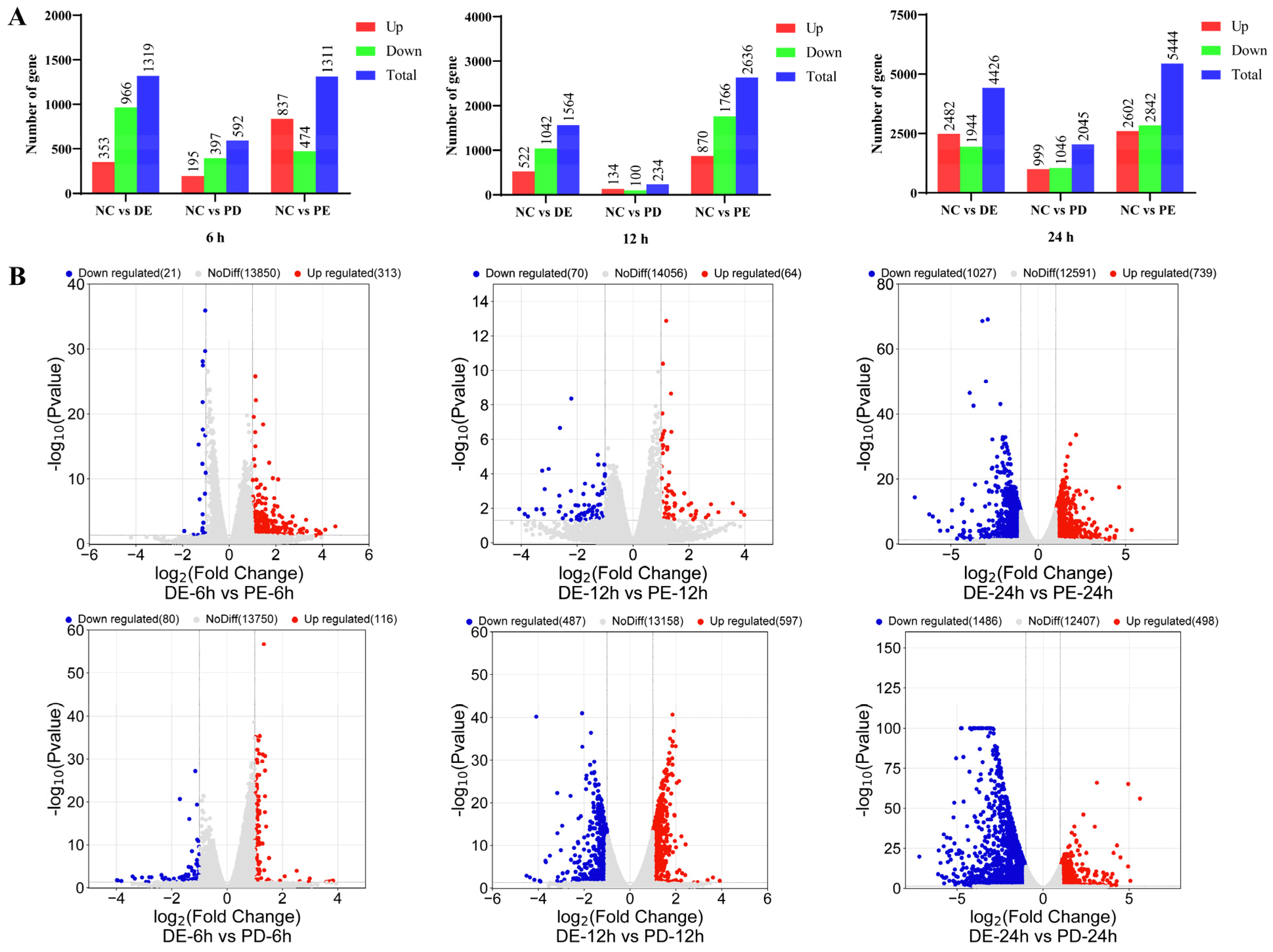
2.8. RNA-Seq Analysis
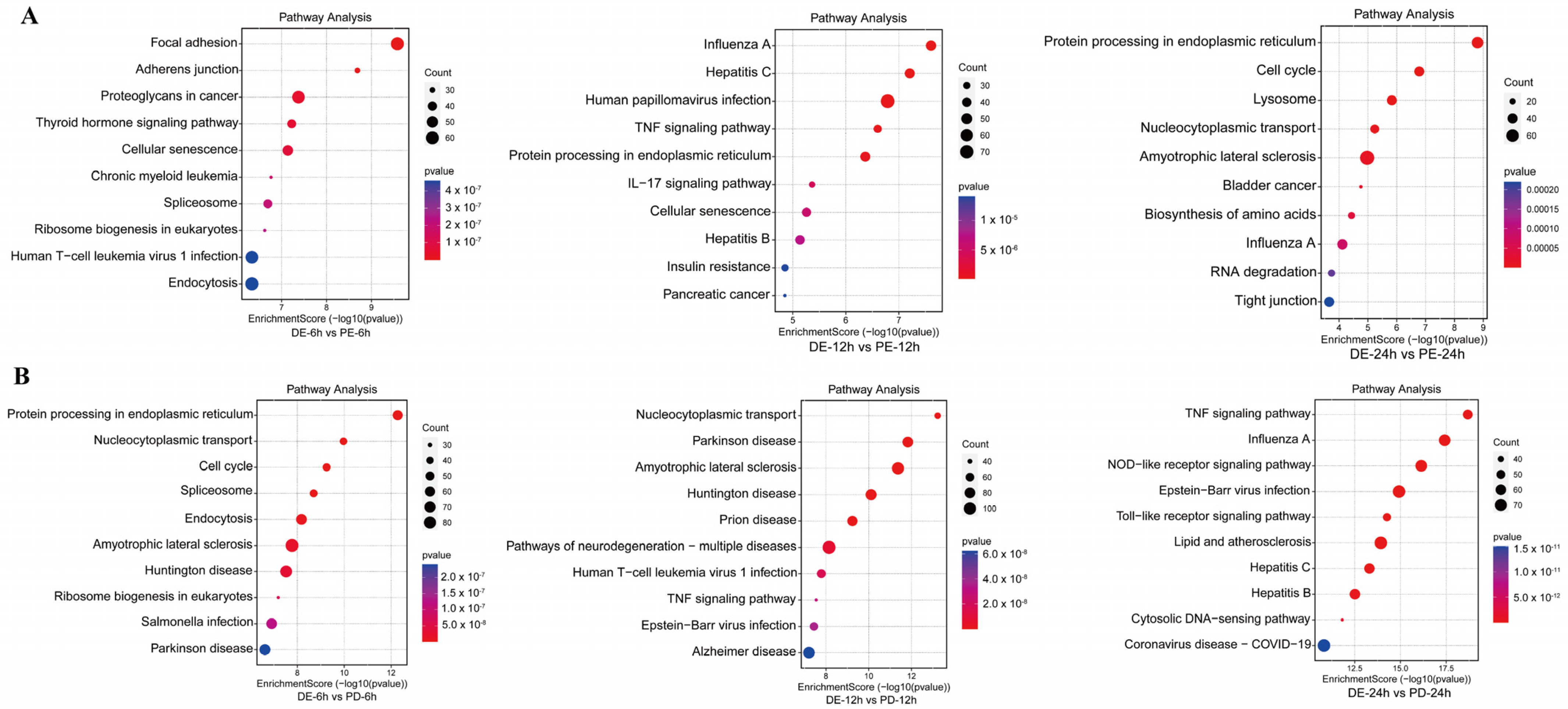
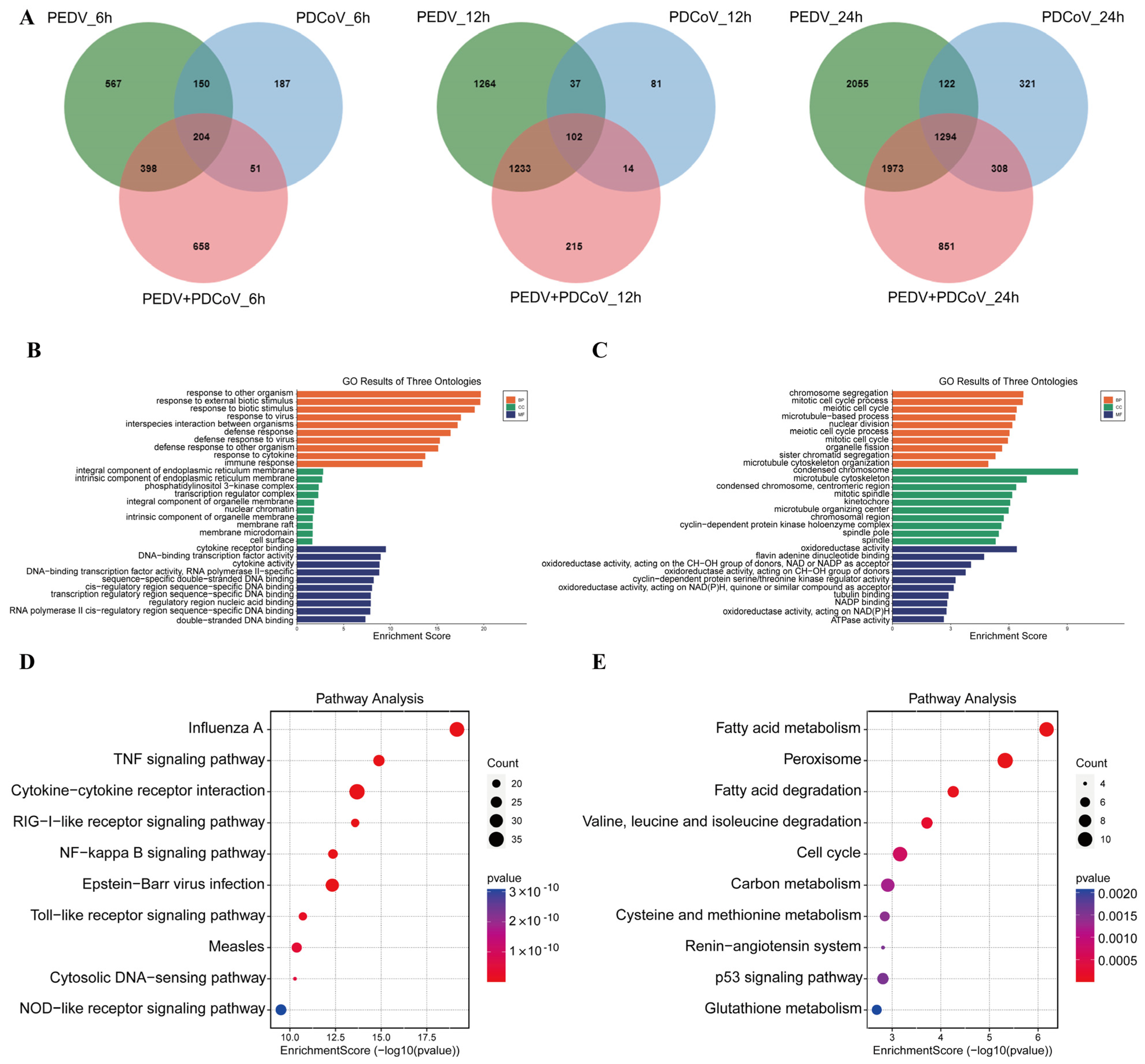
2.9. GO and KEGG Enrichment Analysis
2.10. Overexpression or Knockdown of ISG20
2.11. Western Blotting
2.12. Statistical Analysis
3. Results
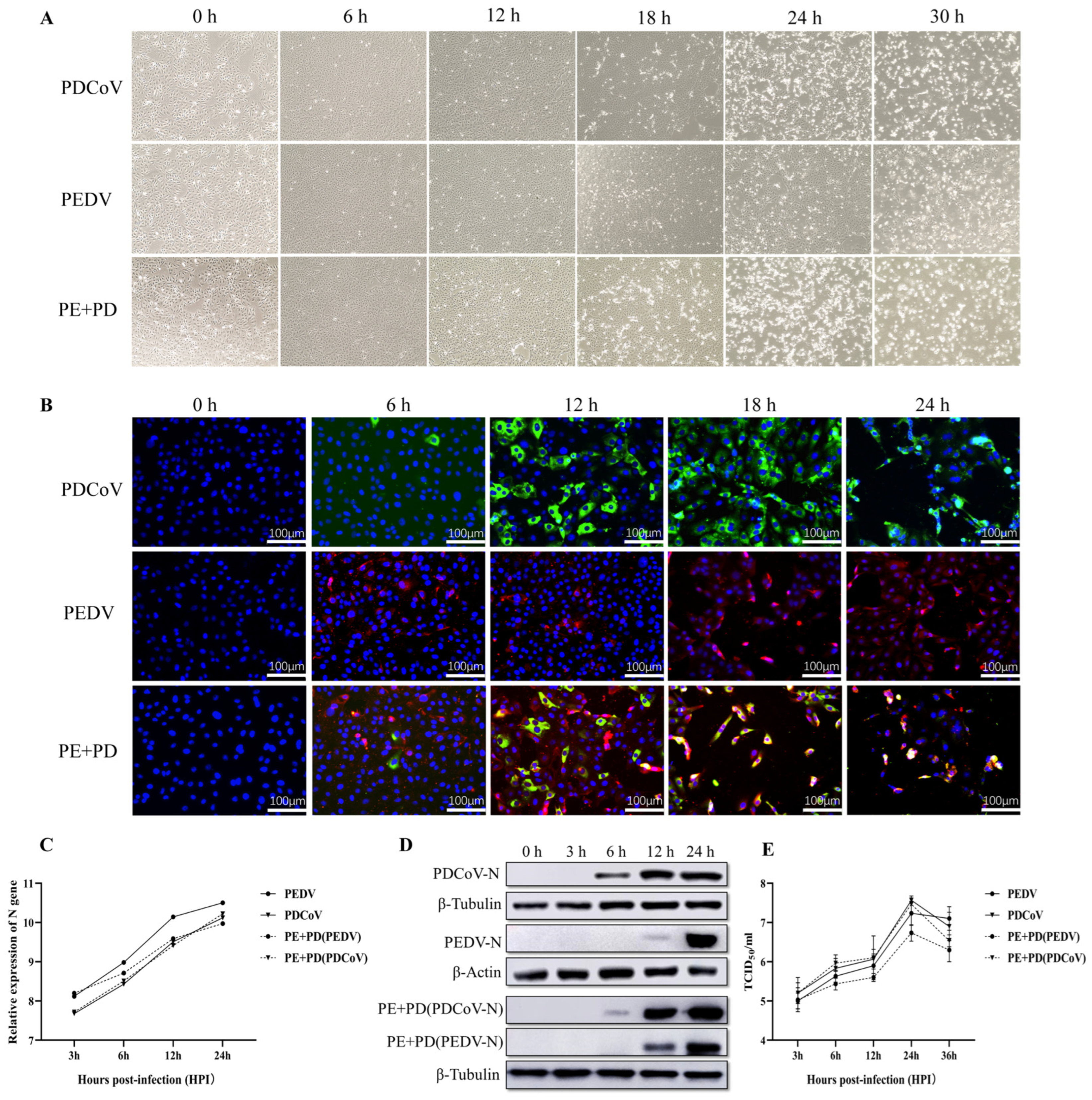
3.1. Proliferation Kinetics of LLC-PK Cells Infected with PEDV, PDCoV, or PEDV and PDCoV
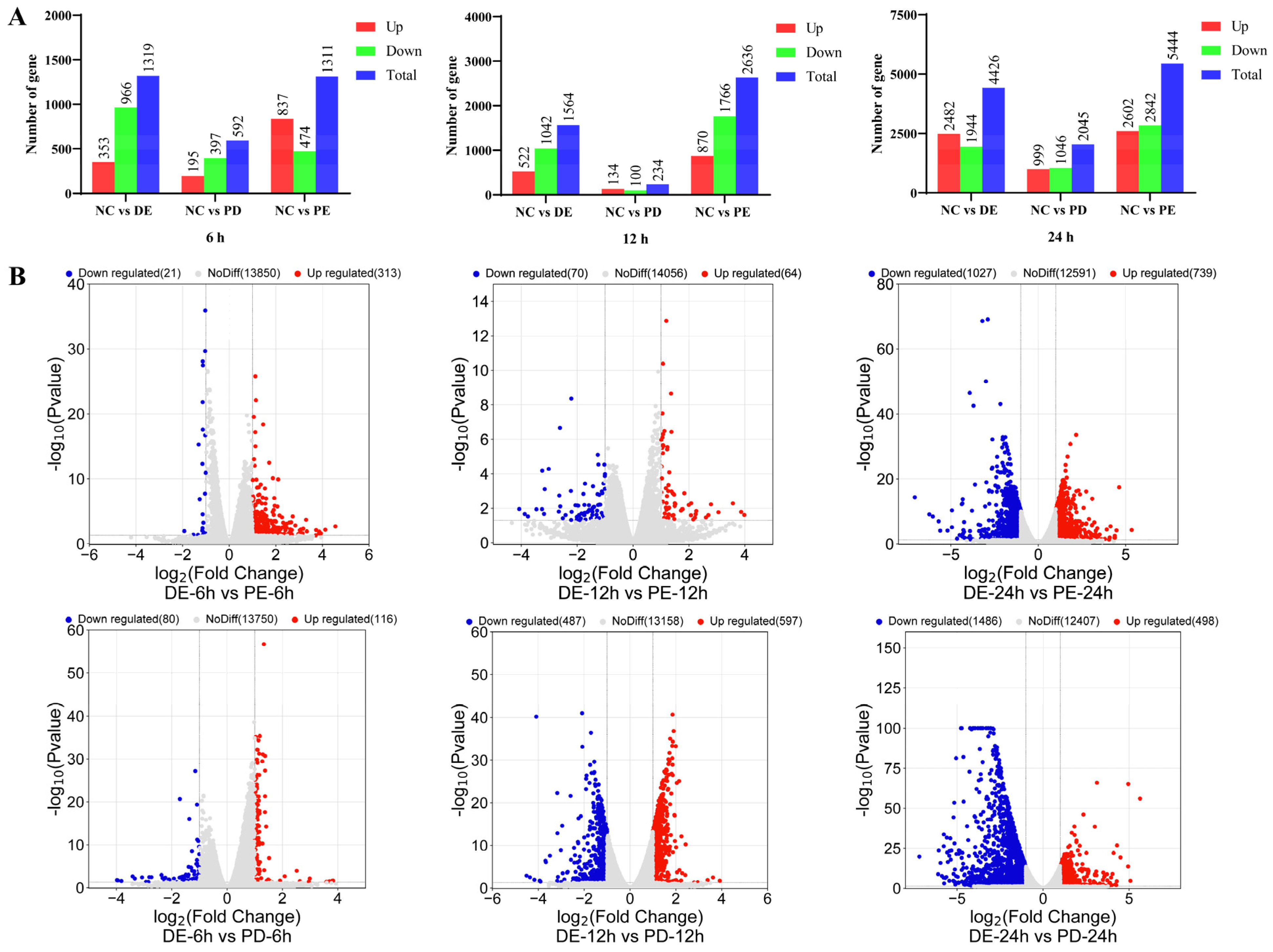
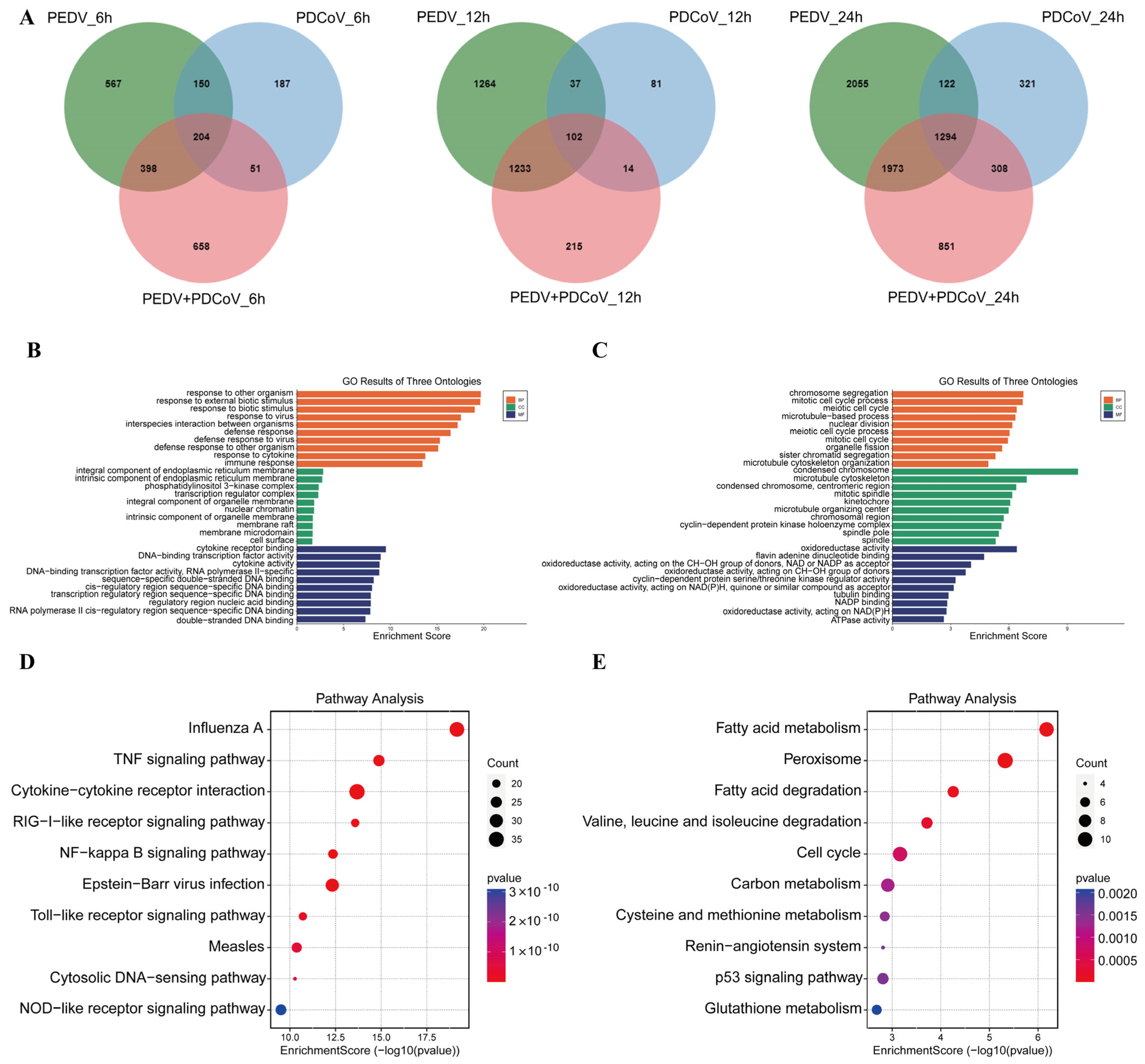
3.2. Transcriptional Profiles of PEDV-Infected, PDCoV-Infected, and PEDV and PDCoV-Coinfected LLC-PK Cells
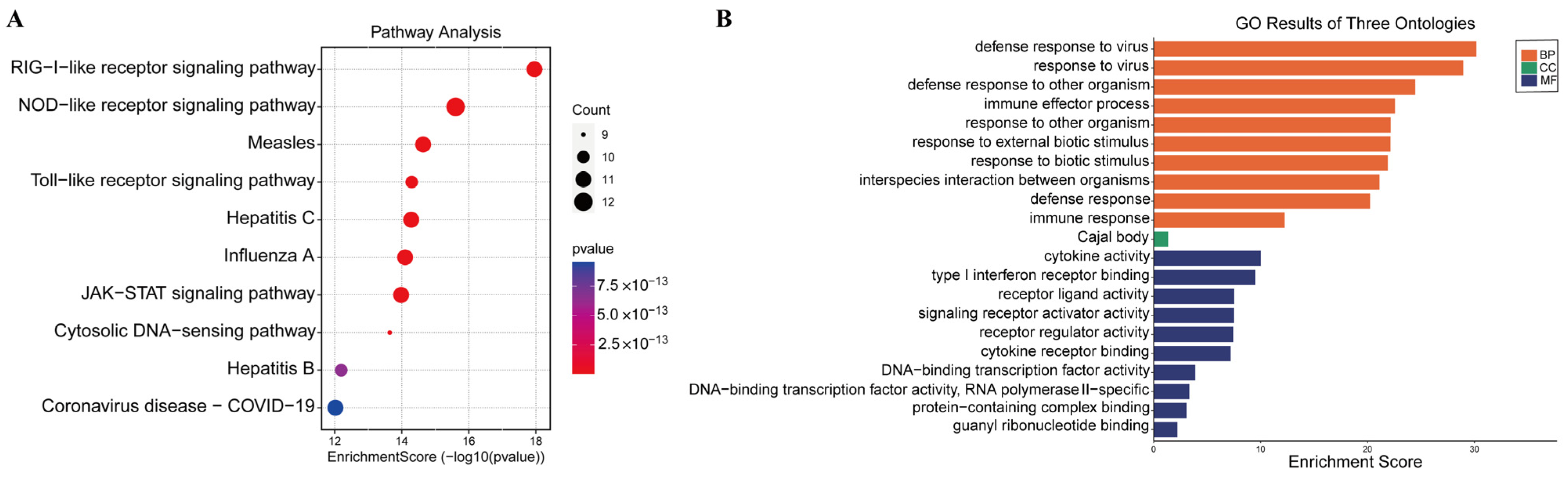
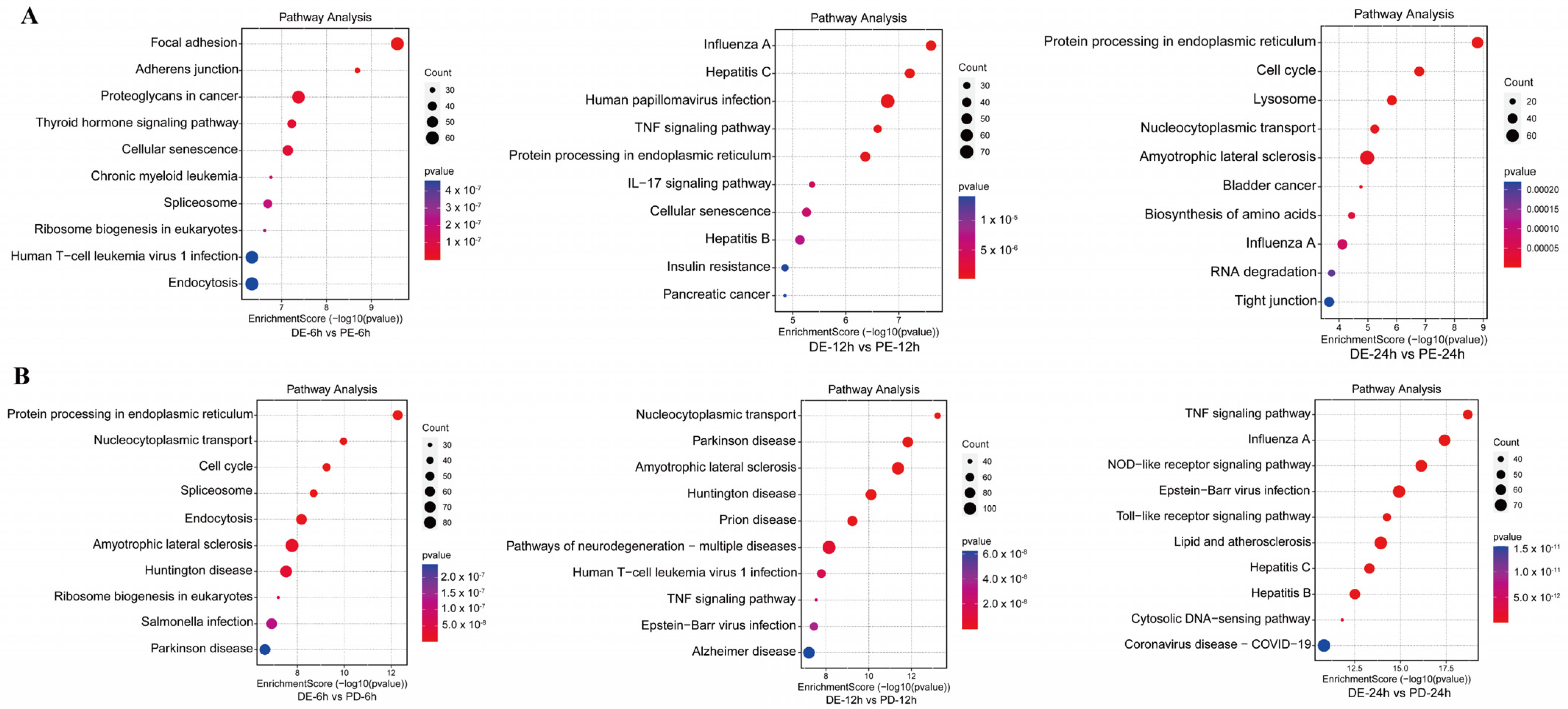
3.3. KEGG Pathway Enrichment Analysis of DEPs between the Coinfection Group and Groups Infected with Individual Viruses
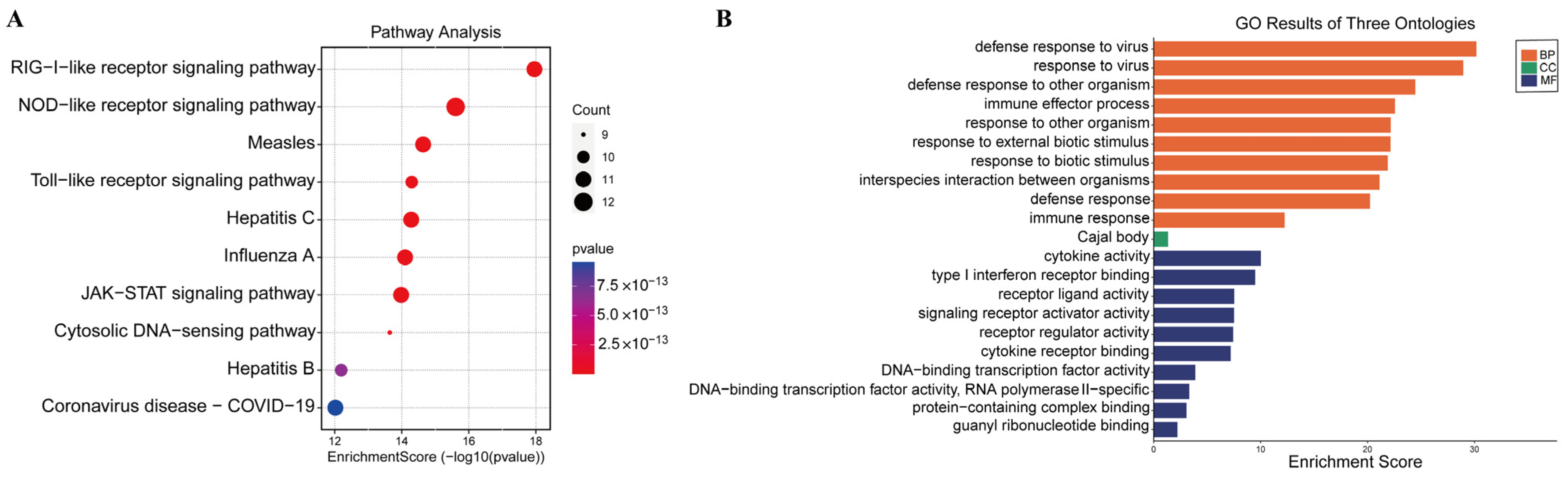
3.4. GO and KEGG Pathway Enrichment Analysis of Shared DEGs
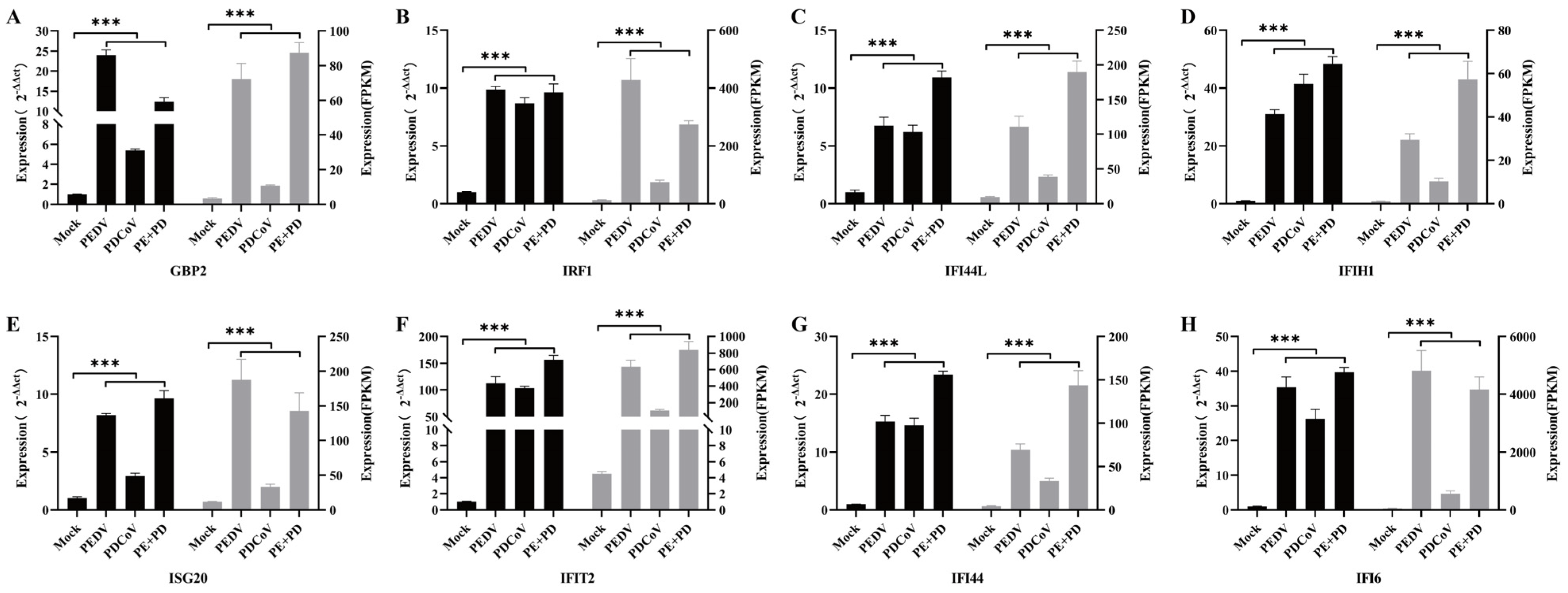
3.5. Assessment of the Expression of Genes Stimulated by Interferons
3.6. Real-Time qPCR Verification of the Differential Expression of ISGs
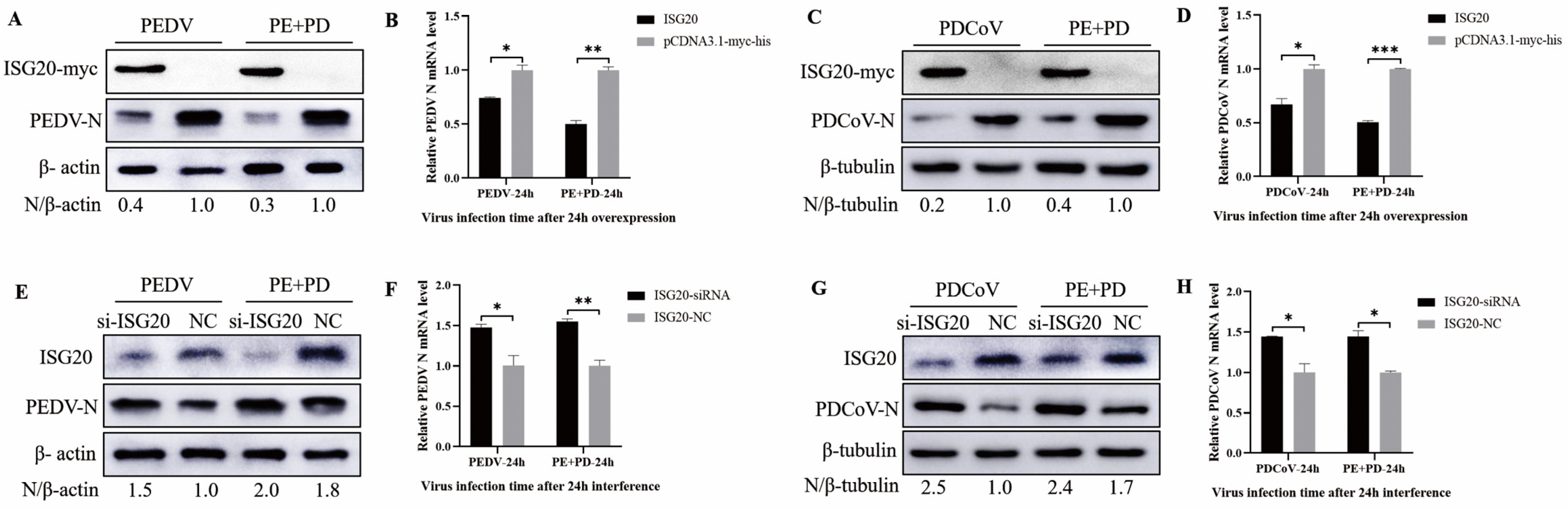
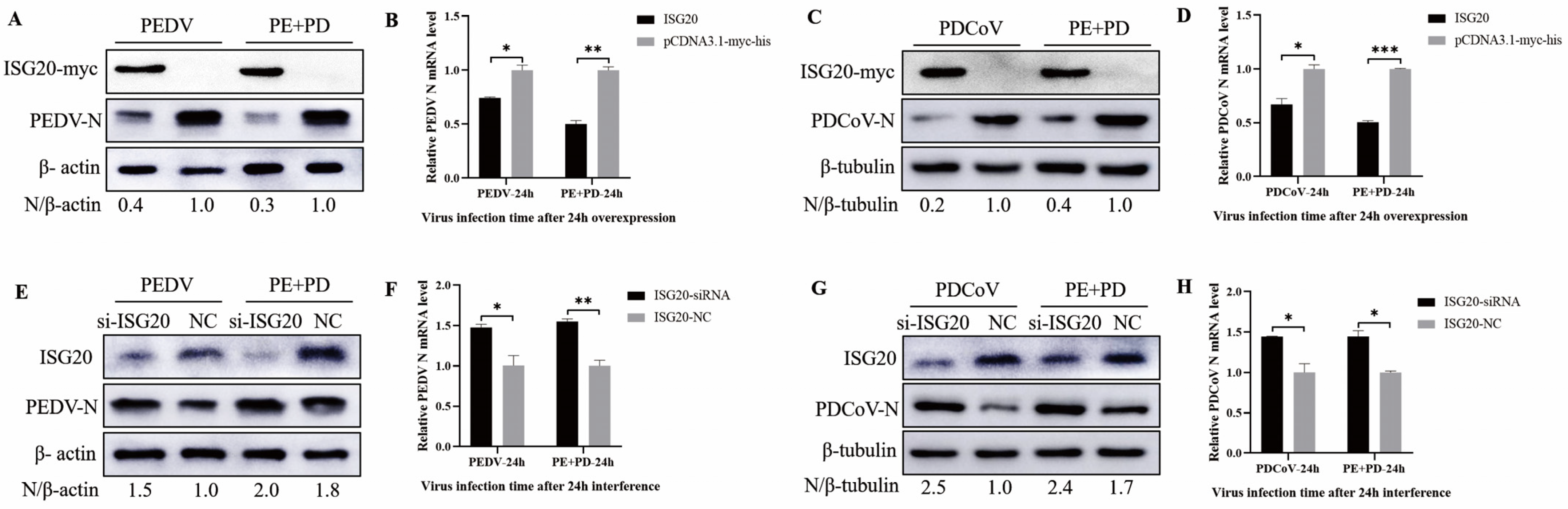
3.7. Overexpression of ISG20 Inhibits Viral Infection, While Knocking down ISG20 Enhanced Viral Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woo, P.C.Y.; Huang, Y.; Lau, S.K.P.; Yuen, K.Y. Coronavirus genomics and bioinformatics analysis. Viruses 2010, 2, 1804–1820. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Jung, K.; Vlasova, A.N.; Chepngeno, J.; Lu, Z.; Wang, Q.; Saif, L.J. Isolation and characterization of porcine deltacoronavirus from pigs with diarrhea in the United States. J. Clin. Microbiol. 2015, 53, 1537–1548. [Google Scholar] [CrossRef] [PubMed]
- Kocherhans, R.; Bridgen, A.; Ackermann, M.; Tobler, K. Completion of the porcine epidemic diarrhoea coronavirus (PEDV) genome sequence. Virus Genes 2001, 23, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Lam, C.S.; Lau, C.C.; Tsang, A.K.; Lau, J.H.; Bai, R.; Teng, J.L.; Tsang, C.C.; Wang, M.; et al. Discovery of seven novel Mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [PubMed]
- Chang, S.H.; Bae, J.L.; Kang, T.J.; Kim, J.; Chung, G.H.; Lim, C.W.; Laude, H.; Yang, M.S.; Jang, Y.S. Identification of the epitope region capable of inducing neutralizing antibodies against the porcine epidemic diarrhea virus. Mol. Cells 2002, 14, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Zheng, Y.; Yang, Y.; Liu, C.; Geng, Q.; Tai, W.; Du, L.; Zhou, Y.; Zhang, W.; Li, F. Cryo-Electron Microscopy Structure of Porcine Deltacoronavirus Spike Protein in the Prefusion State. J. Virol. 2018, 92, e01556-17. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Q.; Zhao, R.; Liu, J.; Zhao, Q.; Zhu, L.; Zhang, B.; Bi, J.; Yang, G.; Liu, J.; Yin, G. Epidemiology and phylogeny of spike gene of porcine epidemic diarrhea virus from Yunnan, China. Virus Res. 2018, 249, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Pensaert, M.B.; de Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef]
- Li, W.; Li, H.; Liu, Y.; Pan, Y.; Deng, F.; Song, Y.; Tang, X.; He, Q. New variants of porcine epidemic diarrhea virus, China, 2011. Emerg. Infect. Dis. 2012, 18, 1350–1353. [Google Scholar] [CrossRef]
- Jung, K.; Hu, H.; Eyerly, B.; Lu, Z.; Chepngeno, J.; Saif, L.J. Pathogenicity of 2 porcine deltacoronavirus strains in gnotobiotic pigs. Emerg. Infect. Dis. 2015, 21, 650–654. [Google Scholar] [CrossRef]
- Ferrara, G.; D’Anza, E.; Rossi, A.; Improda, E.; Iovane, V.; Pagnini, U.; Iovane, G.; Montagnaro, S. A Serological Investigation of Porcine Reproductive and Respiratory Syndrome and Three Coronaviruses in the Campania Region, Southern Italy. Viruses 2023, 15, 300. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, G.; Nocera, F.P.; Longobardi, C.; Ciarcia, R.; Fioretti, A.; Damiano, S.; Iovane, G.; Pagnini, U.; Montagnaro, S. Retrospective Serosurvey of Three Porcine Coronaviruses among the Wild Boar (Sus scrofa) Population in the Campania Region of Italy. J. Wildl. Dis. 2022, 58, 887–891. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Jung, K.; Vlasova, A.N.; Saif, L.J. Experimental infection of gnotobiotic pigs with the cell-culture-adapted porcine deltacoronavirus strain OH-FD22. Arch. Virol. 2016, 161, 3421–3434. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Byrum, B.; Zhang, Y. Detection and genetic characterization of deltacoronavirus in pigs, Ohio, USA, 2014. Emerg. Infect. Dis. 2014, 20, 1227–1230. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Byrum, B.; Zhang, Y. Porcine coronavirus HKU15 detected in 9 US states, 2014. Emerg. Infect. Dis. 2014, 20, 1594–1595. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Zhou, X.; Peng, Q.; Chen, Y.; Zhang, F.; Huang, T.; Zhang, T.; Li, A.; Huang, D.; Wu, Q.; et al. Newly Emerged Porcine Deltacoronavirus Associated with Diarrhoea in Swine in China: Identification, Prevalence and Full-Length Genome Sequence Analysis. Transbound. Emerg. Dis. 2015, 62, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.; Lee, K.K.; Kim, S.H.; Lee, C. Prevalence, complete genome sequencing and phylogenetic analysis of porcine deltacoronavirus in South Korea, 2014–2016. Transbound. Emerg. Dis. 2017, 64, 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Lorsirigool, A.; Saeng-Chuto, K.; Madapong, A.; Temeeyasen, G.; Tripipat, T.; Kaewprommal, P.; Tantituvanont, A.; Piriyapongsa, J.; Nilubol, D. The genetic diversity and complete genome analysis of two novel porcine deltacoronavirus isolates in Thailand in 2015. Virus Genes 2017, 53, 240–248. [Google Scholar] [CrossRef]
- Saeng-Chuto, K.; Lorsirigool, A.; Temeeyasen, G.; Vui, D.T.; Stott, C.J.; Madapong, A.; Tripipat, T.; Wegner, M.; Intrakamhaeng, M.; Chongcharoen, W.; et al. Different Lineage of Porcine Deltacoronavirus in Thailand, Vietnam and Lao PDR in 2015. Transbound. Emerg. Dis. 2017, 64, 3–10. [Google Scholar] [CrossRef]
- Suzuki, T.; Hayakawa, J.; Ohashi, S. Complete Genome Characterization of the Porcine Deltacoronavirus HKD/JPN/2016, Isolated in Japan, 2016. Genome Announc. 2017, 5, e00795-17. [Google Scholar] [CrossRef]
- Ajayi, T.; Dara, R.; Misener, M.; Pasma, T.; Moser, L.; Poljak, Z. Herd-level prevalence and incidence of porcine epidemic diarrhoea virus (PEDV) and porcine deltacoronavirus (PDCoV) in swine herds in Ontario, Canada. Transbound. Emerg. Dis. 2018, 65, 1197–1207. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rivera, C.; Ramírez-Mendoza, H.; Mendoza-Elvira, S.; Segura-Velázquez, R.; Sánchez-Betancourt, J.I. First report and phylogenetic analysis of porcine deltacoronavirus in Mexico. Transbound. Emerg. Dis. 2019, 66, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Saeng-Chuto, K.; Jermsutjarit, P.; Stott, C.J.; Vui, D.T.; Tantituvanont, A.; Nilubol, D. Retrospective study, full-length genome characterization and evaluation of viral infectivity and pathogenicity of chimeric porcine deltacoronavirus detected in Vietnam. Transbound. Emerg. Dis. 2020, 67, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Hu, H.; Saif, L.J. Porcine deltacoronavirus infection: Etiology, cell culture for virus isolation and propagation, molecular epidemiology and pathogenesis. Virus Res. 2016, 226, 50–59. [Google Scholar] [CrossRef]
- Zhang, H.; Liang, Q.; Li, B.; Cui, X.; Wei, X.; Ding, Q.; Wang, Y.; Hu, H. Prevalence, phylogenetic and evolutionary analysis of porcine deltacoronavirus in Henan province, China. Prev. Vet. Med. 2019, 166, 8–15. [Google Scholar] [CrossRef]
- Feng, Y.; Xu, Z.; Zhu, L. Prevalence and phylogenetic analysis of porcine deltacoronavirus in Sichuan province, China. Arch. Virol. 2020, 165, 2883–2889. [Google Scholar] [CrossRef]
- Saeng-Chuto, K.; Madapong, A.; Kaeoket, K.; Piñeyro, P.E.; Tantituvanont, A.; Nilubol, D. Coinfection of porcine deltacoronavirus and porcine epidemic diarrhea virus increases disease severity, cell trophism and earlier upregulation of IFN-α and IL12. Sci. Rep. 2021, 11, 3040. [Google Scholar] [CrossRef]
- Liang, J.; Li, Y.; Yan, Z.; Jiao, Z.; Peng, D.; Zhang, W. Study of the effect of intestinal immunity in neonatal piglets coinfected with porcine deltacoronavirus and porcine epidemic diarrhea virus. Arch. Virol. 2022, 167, 1649–1657. [Google Scholar] [CrossRef]
- Zhang, H.; Han, F.; Shu, X.; Li, Q.; Ding, Q.; Hao, C.; Yan, X.; Xu, M.; Hu, H. Co-infection of porcine epidemic diarrhoea virus and porcine deltacoronavirus enhances the disease severity in piglets. Transbound. Emerg. Dis. 2022, 69, 1715–1726. [Google Scholar] [CrossRef]
- Jiao, Z.; Liang, J.; Yang, Y.; Li, Y.; Yan, Z.; Hu, G.; Gu, C.; Hu, X.; Cheng, G.; Peng, G.; et al. Coinfection of porcine deltacoronavirus and porcine epidemic diarrhea virus altered viral tropism in gastrointestinal tract in a piglet model. Virology 2021, 558, 119–125. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, D.; Zhou, P.; Zhang, L.; Li, M.; Li, W.; Zhang, Y.; Wang, Y.; Liu, X. Characterization, pathogenicity and protective efficacy of a cell culture-derived porcine deltacoronavirus. Virus Res. 2020, 282, 197955. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, X.; Zhang, Q.; Zhou, P.; Fang, Y.; Dong, Z.; Zhao, D.; Li, W.; Feng, J.; Zhang, Y.; et al. Biological characterization and pathogenicity of a newly isolated Chinese highly virulent genotype GIIa porcine epidemic diarrhea virus strain. Arch. Virol. 2019, 164, 1287–1295. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wang, D.; Zhou, J.; Pan, T.; Chen, J.; Yang, Y.; Lv, M.; Ye, X.; Peng, G.; Fang, L.; et al. Porcine Deltacoronavirus nsp5 Antagonizes Type I Interferon Signaling by Cleaving STAT2. J. Virol. 2017, 91, e00003-17. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.; Wen, Y.; Hu, J.; Xiao, D.; Li, S.; Zhang, L.; Liao, Y.; Chen, R.; Zhao, Y.; Wen, Y.; et al. Study of the inhibitory effect of STAT1 on PDCoV infection. Vet. Microbiol. 2022, 266, 109333. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, F.; Ye, S.; Guo, X.; Muhanmmad Memon, A.; Wu, M.; He, Q. Comparative Proteome Analysis of Porcine Jejunum Tissues in Response to a Virulent Strain of Porcine Epidemic Diarrhea Virus and Its Attenuated Strain. Viruses 2016, 8, 323. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Li, B.; Chen, L.; Ma, Z.; He, K.; Fan, H. Differential Protein Analysis of IPEC-J2 Cells Infected with Porcine Epidemic Diarrhea Virus Pandemic and Classical Strains Elucidates the Pathogenesis of Infection. J. Proteome Res. 2017, 16, 2113–2120. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Chen, J.; Li, X.; Ren, W.; Li, F.; Wang, T.; Li, C.; Dong, Z.; Tian, X.; Zhang, L.; et al. Identification and integrated analysis of lncRNAs and miRNAs in IPEC-J2 cells provide novel insight into the regulation of the innate immune response by PDCoV infection. BMC Genom. 2022, 23, 486. [Google Scholar] [CrossRef]
- Mascia, T.; Gallitelli, D. Synergies and antagonisms in virus interactions. Plant Sci. An. Int. J. Exp. Plant Biol. 2016, 252, 176–192. [Google Scholar] [CrossRef]
- Eclercy, J.; Larcher, T.; Andraud, M.; Renson, P.; Bernard, C.; Bigault, L.; Ledevin, M.; Paboeuf, F.; Grasland, B.; Rose, N.; et al. PCV2 co-infection does not impact PRRSV MLV1 safety but enhances virulence of a PRRSV MLV1-like strain in infected SPF pigs. Vet. Microbiol. 2020, 244, 108656. [Google Scholar] [CrossRef]
- Lai, C.C.; Wang, C.Y.; Hsueh, P.R. Co-infections among patients with COVID-19: The need for combination therapy with non-anti-SARS-CoV-2 agents? J. Microbiol. Immunol. Infect. Wei Mian Yu Gan Ran Za Zhi 2020, 53, 505–512. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Z.M.; Chen, L.D.; Zhan, Y.Q.; Li, S.Q.; Cheng, J.; Zhu, A.R.; Chen, L.Y.; Zhong, N.S.; Li, S.Y.; et al. Coinfection with SARS-CoV-2 and other respiratory pathogens in patients with COVID-19 in Guangzhou, China. J. Med. Virol. 2020, 92, 2381–2383. [Google Scholar] [CrossRef]
- Hu, Z.; Li, Y.; Du, H.; Ren, J.; Zheng, X.; Wei, K.; Liu, J. Transcriptome analysis reveals modulation of the STAT family in PEDV-infected IPEC-J2 cells. BMC Genom. 2020, 21, 891. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, T.; Yang, Z.; Wan, J.; Zhang, Y.; Wang, X.; Kong, L.; Wang, T.; Li, Y. Transcriptome analysis of host response to porcine epidemic diarrhea virus nsp15 in IPEC-J2 cells. Microb. Pathog. 2022, 162, 105195. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Li, F.; Li, X.; Wang, L.; Zhang, L.; Lu, C.; Zheng, L.; Yan, M. Transcriptome analysis of PK-15 cells in innate immune response to porcine deltacoronavirus infection. PLoS ONE 2019, 14, e0223177. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Pulido, D.; Boley, P.A.; Ouma, W.Z.; Alhamo, M.A.; Saif, L.J.; Kenney, S.P. Comparative Transcriptome Profiling of Human and Pig Intestinal Epithelial Cells after Porcine Deltacoronavirus Infection. Viruses 2021, 13, 292. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, R.; Xiao, D.; Zhang, L.; Song, D.; Wen, Y.; Wu, R.; Zhao, Q.; Du, S.; Wen, X.; et al. A Comparative Transcriptomic Analysis Reveals That HSP90AB1 Is Involved in the Immune and Inflammatory Responses to Porcine Deltacoronavirus Infection. Int. J. Mol. Sci. 2022, 23, 3280. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-stimulated genes: A complex web of host defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef]
- Zhang, Q.; Yoo, D. Immune evasion of porcine enteric coronaviruses and viral modulation of antiviral innate signaling. Virus Res. 2016, 226, 128–141. [Google Scholar] [CrossRef]
- Li, L.; Fu, F.; Xue, M.; Chen, W.; Liu, J.; Shi, H.; Chen, J.; Bu, Z.; Feng, L.; Liu, P. IFN-lambda preferably inhibits PEDV infection of porcine intestinal epithelial cells compared with IFN-alpha. Antivir. Res. 2017, 140, 76–82. [Google Scholar] [CrossRef]
- Zhang, Q.; Ke, H.; Blikslager, A.; Fujita, T.; Yoo, D. Type III Interferon Restriction by Porcine Epidemic Diarrhea Virus and the Role of Viral Protein nsp1 in IRF1 Signaling. J. Virol. 2018, 92, e01677-17. [Google Scholar] [CrossRef]
- Liu, S.; Fang, P.; Ke, W.; Wang, J.; Wang, X.; Xiao, S.; Fang, L. Porcine deltacoronavirus (PDCoV) infection antagonizes interferon-λ1 production. Vet. Microbiol. 2020, 247, 108785. [Google Scholar] [CrossRef] [PubMed]
- Gongora, C.; David, G.; Pintard, L.; Tissot, C.; Hua, T.D.; Dejean, A.; Mechti, N. Molecular cloning of a new interferon-induced PML nuclear body-associated protein. J. Biol. Chem. 1997, 272, 19457–19463. [Google Scholar] [CrossRef] [PubMed]
- Gongora, C.; Degols, G.; Espert, L.; Hua, T.D.; Mechti, N. A unique ISRE, in the TATA-less human Isg20 promoter, confers IRF-1-mediated responsiveness to both interferon type I and type II. Nucleic Acids Res. 2000, 28, 2333–2341. [Google Scholar] [CrossRef] [PubMed]
- Degols, G.; Eldin, P.; Mechti, N. ISG20, an actor of the innate immune response. Biochimie 2007, 89, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Wang, N.; Woodson, S.E.; Dong, Q.; Wang, J.; Liang, Y.; Rijnbrand, R.; Wei, L.; Nichols, J.E.; Guo, J.T.; et al. Antiviral activities of ISG20 in positive-strand RNA virus infections. Virology 2011, 409, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Nguyen, X.N.; Wang, L.; Appourchaux, R.; Zhang, C.; Panthu, B.; Gruffat, H.; Journo, C.; Alais, S.; Qin, J.; et al. The interferon stimulated gene 20 protein (ISG20) is an innate defense antiviral factor that discriminates self versus non-self translation. PLoS Pathog. 2019, 15, e1008093. [Google Scholar] [CrossRef]
- Kang, D.; Gao, S.; Tian, Z.; Zhang, G.; Guan, G.; Liu, G.; Luo, J.; Du, J.; Yin, H. ISG20 inhibits bluetongue virus replication. Virol. Sin. 2022, 37, 521–530. [Google Scholar] [CrossRef]
- Qu, H.; Li, J.; Yang, L.; Sun, L.; Liu, W.; He, H. Influenza A Virus-induced expression of ISG20 inhibits viral replication by interacting with nucleoprotein. Virus Genes. 2016, 52, 759–767. [Google Scholar] [CrossRef]
- Espert, L.; Degols, G.; Gongora, C.; Blondel, D.; Williams, B.R.; Silverman, R.H.; Mechti, N. ISG20, a new interferon-induced RNase specific for single-stranded RNA, defines an alternative antiviral pathway against RNA genomic viruses. J. Biol. Chem. 2003, 278, 16151–16158. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer | Reverse Primer | Amplicon Size (bp) |
|---|---|---|---|
| GBP2 | GCAGCTACCGGAAGAAGATG | TTGACAAAGGTCAGCACCAG | 153 |
| IRF1 | AAGTCCAGCCGAGATGCTAA | GGCCTGTTCAATGTCCAAGT | 162 |
| IFI44L | GGCCATAGTGGGCTCTGATA | CTGTGATTCAGCGATGGAGA | 231 |
| IFIH1 | CTCAAAGAGCATCCCCTGAG | GTTCGAACTCTTTGCGGAAG | 246 |
| ISG20 | AGATCCTGCAGCTCCTGAAA | CCAACACACTGTCCTGGATG | 216 |
| IFIT2 | CCTGGGAAGACTTGCAGAAG | GGCCAGTTATCCAGACGGTA | 249 |
| IFI44 | GAGGCCTGGTTCACCAAATA | GGCACAGTCTGCCTTCTTTC | 249 |
| IFI6 | CCAAAACGAAACGCAATACA | CTCCACCGCAGGTGTAGAGT | 163 |
| β-Actin | CTTCCTGGGCATGGAGTCC | GGCGCGATGATCTTGATCTTC | 201 |
| Gene ID | PEDV Infection | PDCoV Infection | PEDV + PDCoV Infection | Description | |||
|---|---|---|---|---|---|---|---|
| log2 Fold Change | p Values | log2 Fold Change | p Values | log2 Fold Change | p Values | ||
| ENSSSCG00000002002 | 6.310629225 | 6.20 × 10−11 | 5.494214783 | 1.66 × 10−7 | 6.767597361 | 1.27 × 10−15 | interferon regulatory factor 9 |
| ENSSSCG00000003091 | 1.234811026 | 7.39 × 10−12 | – a | – | 1.401705148 | 1.30 × 10−14 | interferon regulatory factor 2 binding protein 1 |
| ENSSSCG00000003178 | 1.685537515 | 1.08 × 10−21 | – | – | 1.342602373 | 7.28 × 10−19 | interferon regulatory factor 3 |
| ENSSSCG00000003763 | 4.008604395 | 3.69 × 10−100 | 2.809789843 | 2.44 × 10−84 | 4.972617711 | 9.57 × 10−107 | interferon induced protein 44 |
| ENSSSCG00000005136 | 1.089794081 | 7.66 × 10−3 | – | – | – | – | interferon epsilon |
| ENSSSCG00000005163 | 1.479543723 | 9.92 × 10−38 | 1.257973246 | 2.68 × 10−13 | 1.887533485 | 6.16 × 10−18 | interferon beta 1 |
| ENSSSCG00000006923 | 4.468281465 | 4.91 × 10−71 | 1.575633517 | 1.79 × 10−26 | 4.658103435 | 5.66 × 10−188 | guanylate binding protein 2, interferon-inducible |
| ENSSSCG00000010451 | 7.219000344 | 1.99 × 10−135 | 4.494882714 | 7.29 × 10−103 | 7.526928847 | 1.71 × 10−221 | interferon induced protein with tetratricopeptide repeats 2 |
| ENSSSCG00000010452 | 6.567469339 | 5.33 × 10−229 | 4.725627596 | 7.70 × 10−53 | 7.425033228 | 1.81 × 10−114 | interferon-induced protein with tetratricopeptide repeats 1 |
| ENSSSCG00000010454 | 1.739174062 | 2.76 × 10−24 | – | – | 2.829380505 | 8.35 × 10−82 | interferon-induced protein with tetratricopeptide repeats 5 |
| ENSSSCG00000011402 | 1.161609759 | 9.63 × 10−11 | – | – | – | – | interferon related developmental regulator 2 |
| ENSSSCG00000012076 | 7.31026276 | 7.83 × 10−51 | 4.816830348 | 6.30 × 10−128 | 7.525711196 | 6.16 × 10−38 | Interferon-induced GTP-binding protein Mx2 |
| ENSSSCG00000012853 | 6.744825817 | 5.60 × 10−37 | 3.394978684 | 2.39 × 10−55 | 6.294980934 | 1.30 × 10−76 | interferon regulatory factor 7 |
| ENSSSCG00000014277 | 5.1491548 | 9.84 × 10−50 | 2.465843809 | 8.59 × 10−74 | 4.412829187 | 3.74 × 10−181 | interferon regulatory factor 1 |
| ENSSSCG00000014349 | 3.849359797 | 1.34 × 10−16 | – | – | 2.028133067 | 3.05 × 10−28 | stimulator of interferon response cGAMP interactor 1 |
| ENSSSCG00000014565 | 3.847166684 | 1.17 × 10−36 | – | – | 2.813431636 | 2.94 × 10−90 | interferon-induced transmembrane protein 1 |
| ENSSSCG00000015612 | 1.341585174 | 5.57 × 10−12 | – | – | – | – | interferon regulatory factor 6 |
| ENSSSCG00000015782 | 3.485083398 | 1.51 × 10−35 | 1.173186109 | 2.19 × 10−14 | 3.497169917 | 5.54 × 10−109 | interferon regulatory factor 2 |
| ENSSSCG00000015897 | 4.757076534 | 7.59 × 10−125 | 3.087070238 | 2.77 × 10−49 | 5.62226484 | 1.45 × 10−99 | interferon induced with helicase C domain 1 |
| ENSSSCG00000016573 | 3.00658889 | 1.38 × 10−12 | 1.889624403 | 8.31 × 10−16 | 3.031780305 | 6.00 × 10−42 | interferon regulatory factor 5 |
| ENSSSCG00000021504 | 10.66478654 | 7.55 × 10−13 | 8.54769866 | 4.67 × 10−36 | 9.668453582 | 2.20 × 10−9 | interferon-alphaomega |
| ENSSSCG00000024867 | 4.121896082 | 3.24 × 10−43 | 1.47607923 | 1.39 × 10−26 | 3.640161424 | 1.43 × 10−27 | interferon stimulated exonuclease gene 20 |
| ENSSSCG00000024973 | 6.542282639 | 7.60 × 10−71 | 2.506733001 | 1.20 × 10−61 | 5.723463173 | 1.22 × 10−257 | guanylate binding protein 1, interferon-inducible |
| ENSSSCG00000027660 | 3.578893585 | 1.57 × 10−45 | 1.917801061 | 3.74 × 10−47 | 4.265499699 | 7.20 × 10−173 | interferon induced protein 44 like |
| ENSSSCG00000029763 | 2.447517449 | 9.82 × 10−44 | – | – | 1.746768492 | 2.81 × 10−31 | interferon induced protein 35 |
| ENSSSCG00000031149 | 2.654658654 | 4.72 × 10−19 | – | – | 2.754558433 | 1.63 × 10−5 | interferon-alpha-15 |
| ENSSSCG00000031529 | 9.039504874 | 8.45 × 10−92 | 2.829509587 | 2.93 × 10−3 | 8.194740952 | 3.70 × 10−35 | interferon omega 1 |
| ENSSSCG00000032356 | 1.356456477 | 1.41 × 10−25 | 1.256657445 | 7.59 × 10−4 | 1.634339027 | 3.09 × 10−19 | interferon lambda-4 |
| ENSSSCG00000032591 | 1.069077146 | 2.03 × 10−9 | – | – | – | – | interferon-induced transmembrane protein 1-like |
| ENSSSCG00000033089 | 9.881950086 | 6.94 × 10−73 | 5.947358031 | 1.25 × 10−32 | 9.656541338 | 1.04 × 10−199 | interferon-induced very large GTPase 1-like |
| ENSSSCG00000033115 | – | – | – | – | 1.643246776 | 3.95 × 10−28 | interferon alpha and beta receptor subunit 1 |
| ENSSSCG00000034570 | 6.605547683 | 9.58 × 10−93 | 3.338971302 | 2.86 × 10−29 | 6.307464556 | 1.72 × 10−134 | interferon alpha inducible protein 6 |
| ENSSSCG00000034980 | 7.781934489 | 1.95 × 10−18 | 3.806621518 | 3.53 × 10−4 | 6.767895161 | 5.66 × 10−29 | interferon regulatory factor 8 |
| ENSSSCG00000035185 | 1.388993434 | 3.10 × 10−25 | 1.236766902 | 2.84 × 10−6 | 1.323690765 | 1.45 × 10−10 | interferon lambda-3-like |
| ENSSSCG00000036439 | 10.59936502 | 2.23 × 10−53 | 7.451354106 | 1.02 × 10−57 | 10.6376492 | 4.52 × 10−123 | interleukin 29 (interferon, lambda 1) |
| ENSSSCG00000036556 | 2.721750164 | 1.37 × 10−48 | – | – | 2.818164244 | 1.79 × 10−71 | interferon alpha and beta receptor subunit 2 |
| ENSSSCG00000038132 | 1.559518738 | 3.75 × 10−20 | – | – | 2.022454176 | 2.29 × 10−46 | interferon regulatory factor 2 binding protein 2 |
| ENSSSCG00000038912 | 2.834263813 | 8.28 × 10−29 | – | – | 2.48353821 | 1.41 × 10−46 | interferon induced transmembrane protein 3 |
| ENSSSCG00000040814 | 1.599755345 | 3.76 × 10−21 | 1.387675563 | 3.19 × 10−4 | 1.597754332 | 2.26 × 10−6 | interleukin 28B (interferon, lambda 3) |
| ENSSSCG00000043301 | 2.108776423 | 3.21 × 10−4 | 2.386654345 | 4.87 × 10−2 | 2.386454745 | 5.77 × 10−6 | interferon-alpha-8 |
| ENSSSCG00000044440 | 1.389538790 | 5.86 × 10−13 | – | – | 1.385354889 | 3.18 × 10−8 | interferon-alpha-4 |
| ENSSSCG00000045163 | 1.298445331 | 2.83 × 10−21 | – | – | 1.534679975 | 1.43 × 10−9 | interferon-alpha-9 |
| ENSSSCG00000050619 | 3.678654549 | 6.64 × 10−46 | 2.66908943 | 1.03 × 10−2 | 3.598458533 | 8.29 × 10−34 | Sus scrofa interferon, alpha 1 (IFNA1), mRNA. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Yu, R.; Tong, L.; Zhang, L.; Zhang, Z.; Pan, L.; Wang, Y.; Guo, H.; Hu, Y.; Liu, X. Transcriptome Analysis of LLC-PK Cells Single or Coinfected with Porcine Epidemic Diarrhea Virus and Porcine Deltacoronavirus. Viruses 2024, 16, 74. https://doi.org/10.3390/v16010074
Lu Y, Yu R, Tong L, Zhang L, Zhang Z, Pan L, Wang Y, Guo H, Hu Y, Liu X. Transcriptome Analysis of LLC-PK Cells Single or Coinfected with Porcine Epidemic Diarrhea Virus and Porcine Deltacoronavirus. Viruses. 2024; 16(1):74. https://doi.org/10.3390/v16010074
Chicago/Turabian StyleLu, Yanzhen, Ruiming Yu, Lixin Tong, Liping Zhang, Zhongwang Zhang, Li Pan, Yonglu Wang, Huichen Guo, Yonghao Hu, and Xinsheng Liu. 2024. "Transcriptome Analysis of LLC-PK Cells Single or Coinfected with Porcine Epidemic Diarrhea Virus and Porcine Deltacoronavirus" Viruses 16, no. 1: 74. https://doi.org/10.3390/v16010074
APA StyleLu, Y., Yu, R., Tong, L., Zhang, L., Zhang, Z., Pan, L., Wang, Y., Guo, H., Hu, Y., & Liu, X. (2024). Transcriptome Analysis of LLC-PK Cells Single or Coinfected with Porcine Epidemic Diarrhea Virus and Porcine Deltacoronavirus. Viruses, 16(1), 74. https://doi.org/10.3390/v16010074