

Phenothiazines Inhibit SARS-CoV-2 Entry through Targeting Spike Protein

Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Compounds
2.2. Plasmids and Pseudovirus Production
2.3. Luciferase Assay on Pseudovirus Infection
2.4. Cytotoxicity Assay
2.5. Time-of-Addition Assay
2.6. Drug or Virus Pretreatment Assay
2.7. Surface Plasmon Resonance (SPR) Binding Analysis
2.8. CTSL-Mediated the Cleavage of SARS-CoV-2 Spike Protein In Vitro
2.9. The Cleavage of SARS-CoV-2 Spike Protein
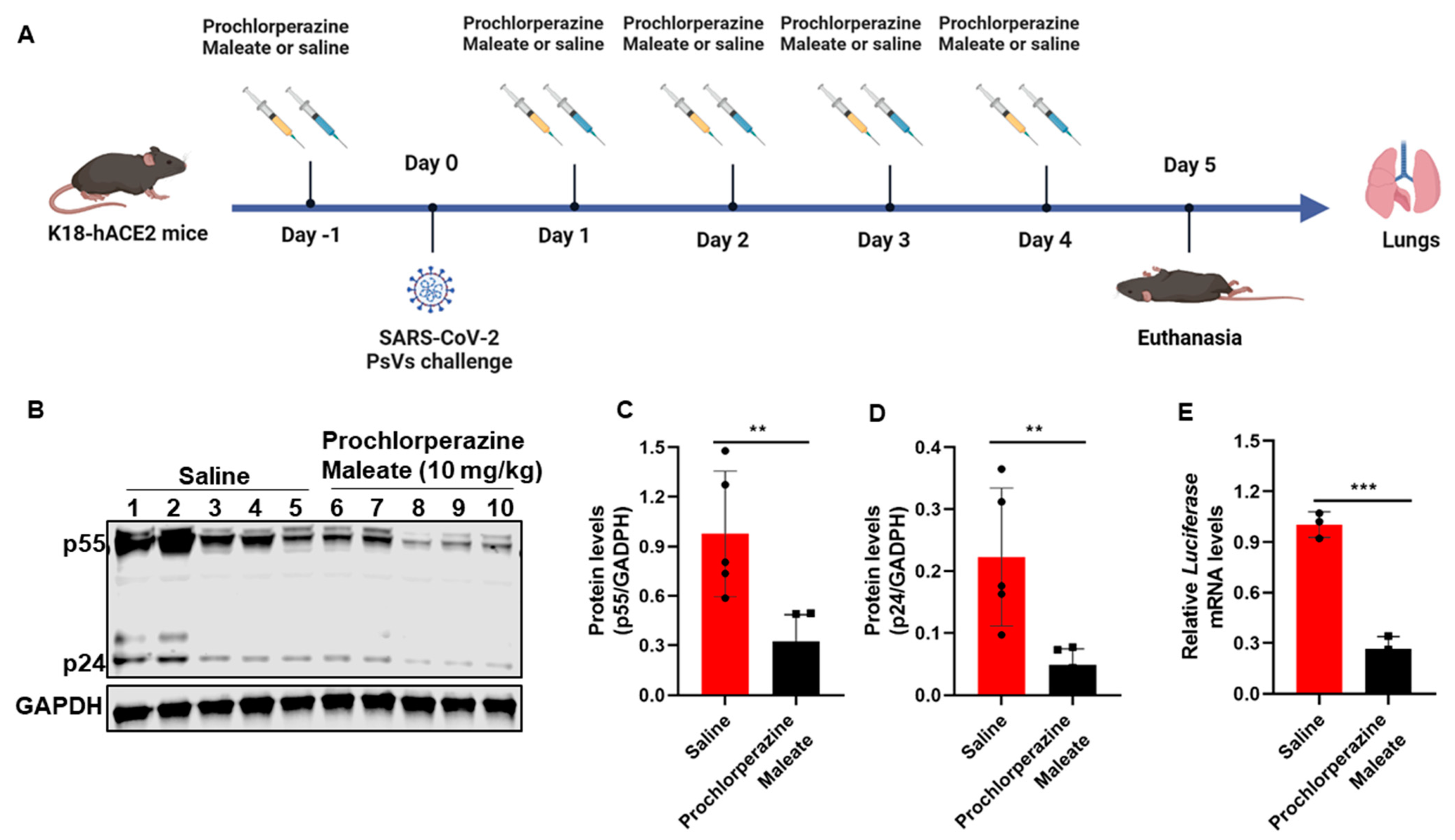
2.10. Antiviral Experiments In Vivo
2.11. Real-Time Quantitative PCR (RT-qPCR)
2.12. Statistical Analysis
3. Results
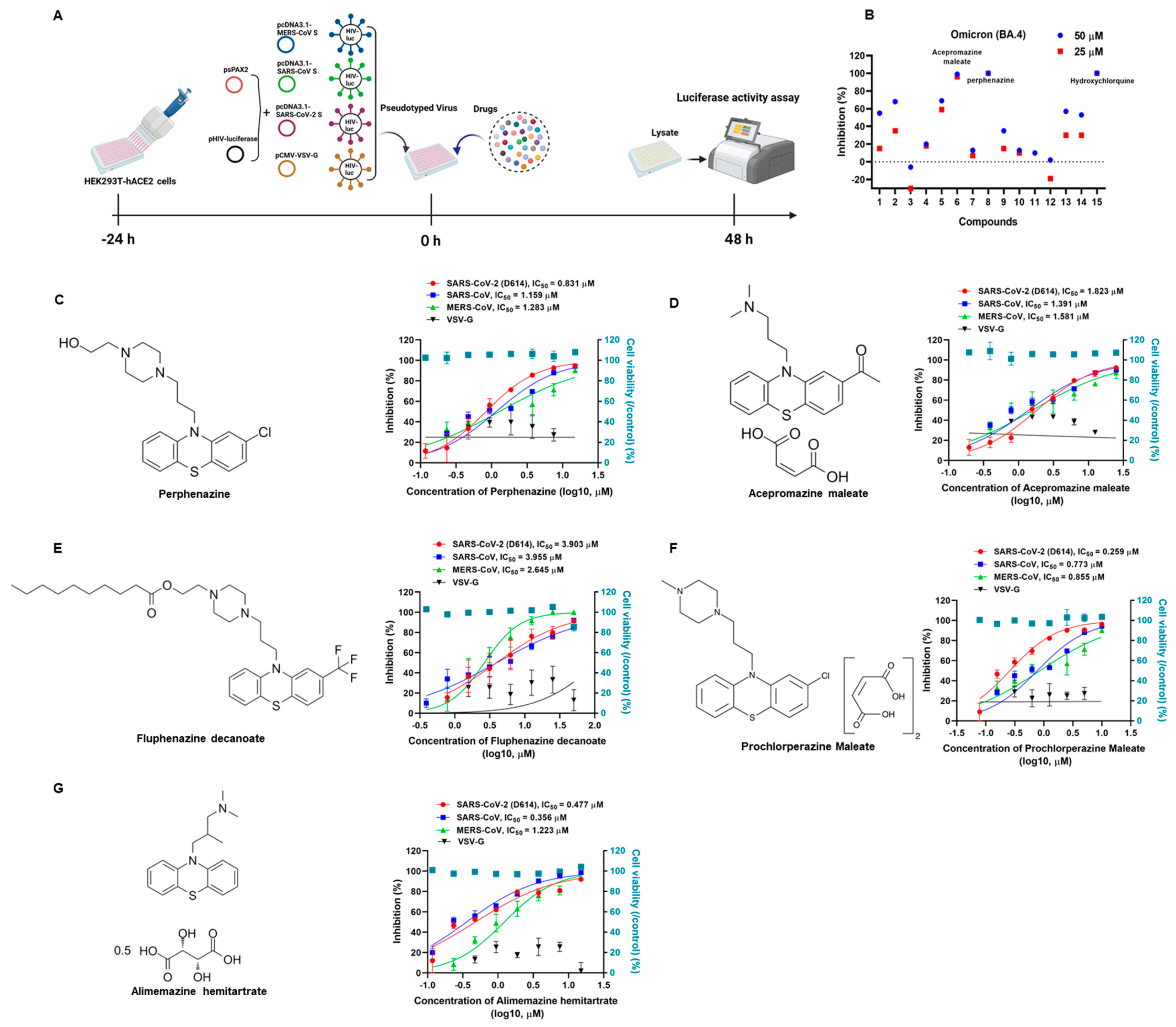
3.1. Phenothiazines Show Potent Broad-Spectrum Antiviral Activities
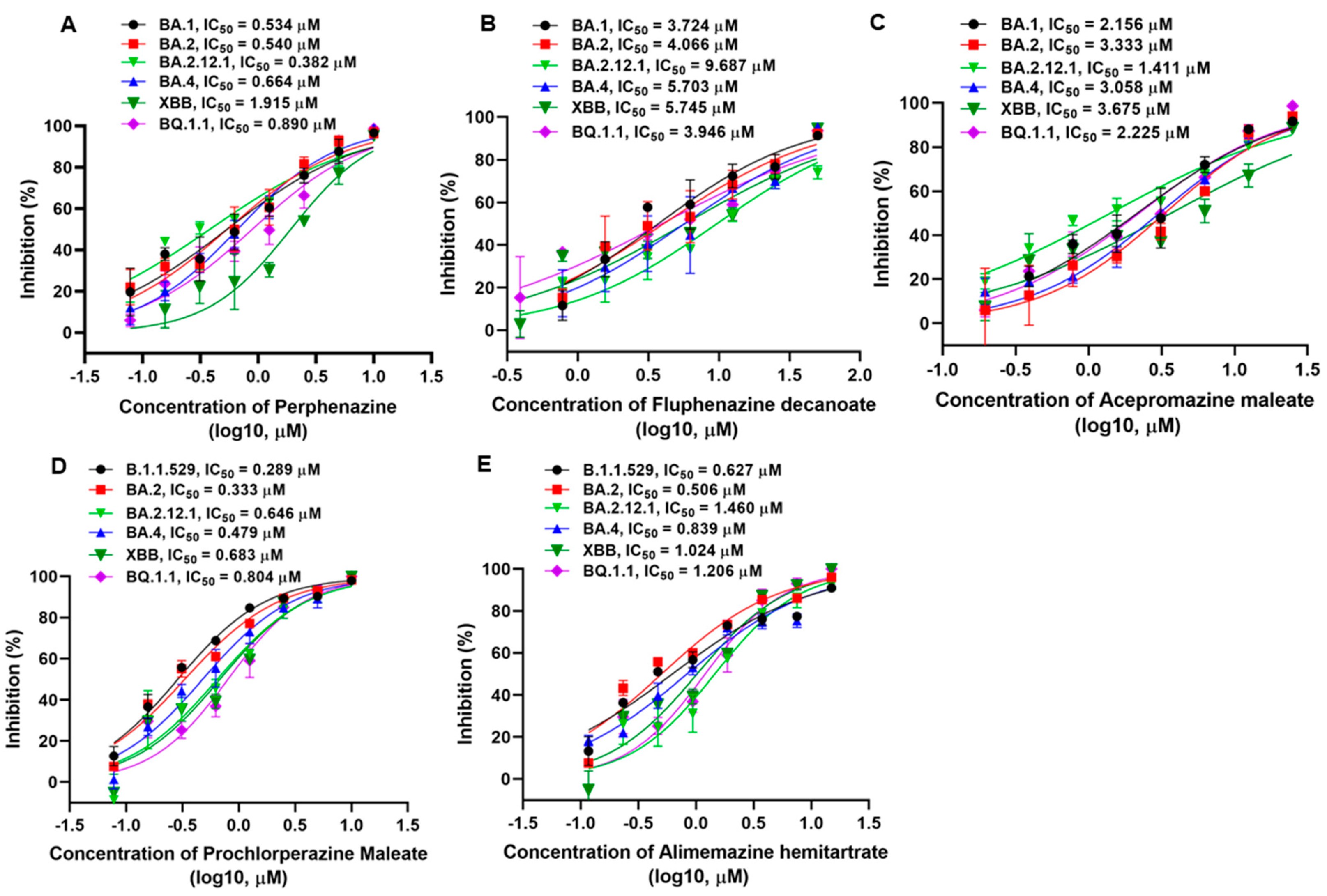
3.2. Phenothiazines Inhibit the Entry of Multiple SARS-CoV-2 Variants
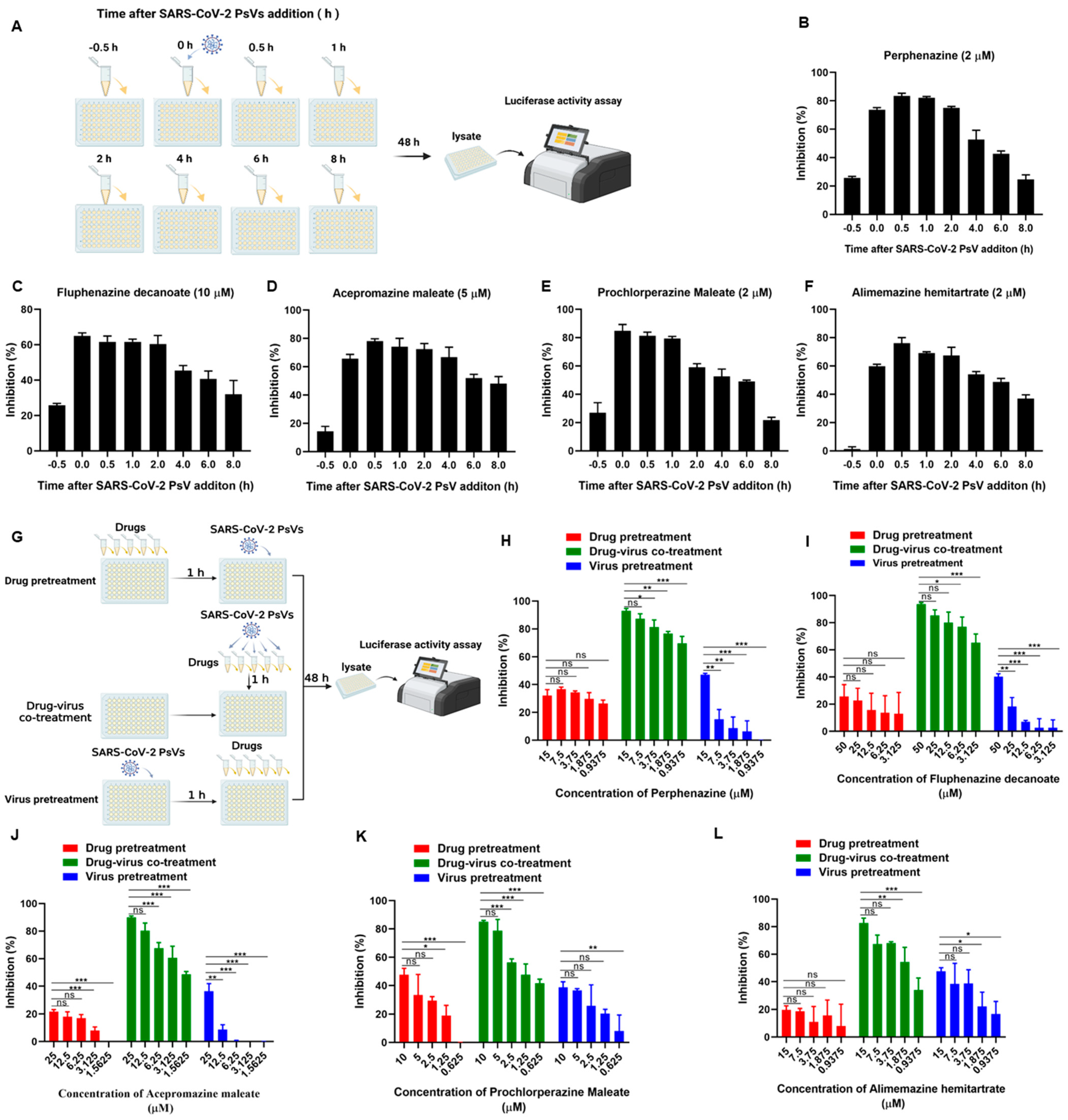
3.3. Phenothiazines Predominantly Inhibit SARS-CoV-2 PsV Infection at the Early Stage
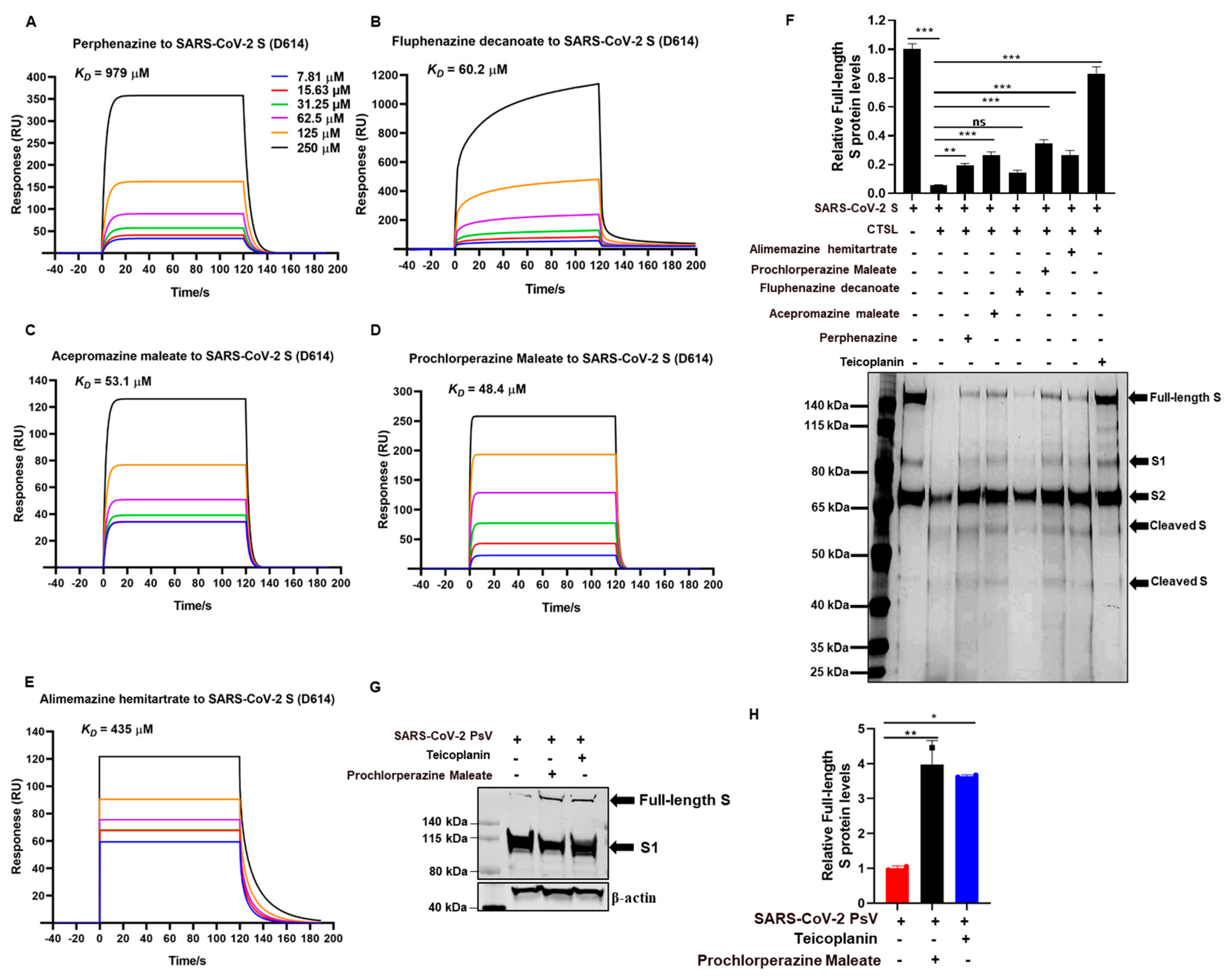
3.4. Phenothiazines Bind to S Protein and Interfere S Protein Cleavage
3.5. Prochlorperazine Maleate Inhibits SARS-CoV-2 PsV Infection in K18-hACE2 Mice

4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Li, X.; Li, T.; Zhang, S.; Wang, L.; Wu, X.; Liu, J. The genetic sequence, origin, and diagnosis of SARS-CoV-2. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1629–1635. [Google Scholar] [CrossRef]
- Telenti, A.; Arvin, A.; Corey, L.; Corti, D.; Diamond, M.S.; García-Sastre, A.; Garry, R.F.; Holmes, E.C.; Pang, P.S.; Virgin, H.W. After the pandemic: Perspectives on the future trajectory of COVID-19. Nature 2021, 596, 495–504. [Google Scholar] [CrossRef]
- Brady, D.K.; Gurijala, A.R.; Huang, L.; Hussain, A.A.; Lingan, A.L.; Pembridge, O.G.; Ratangee, B.A.; Sealy, T.T.; Vallone, K.T.; Clements, T.P. A guide to COVID-19 antiviral therapeutics: A summary and perspective of the antiviral weapons against SARS-CoV-2 infection. Febs. J. 2022. [Google Scholar] [CrossRef] [PubMed]
- Reina, J.; Iglesias, C. Nirmatrelvir plus ritonavir (Paxlovid) a potent SARS-CoV-2 3CLpro protease inhibitor combination. Rev. Esp. Quim. 2022, 35, 236–240. [Google Scholar] [CrossRef]
- Wen, W.; Chen, C.; Tang, J.; Wang, C.; Zhou, M.; Cheng, Y.; Zhou, X.; Wu, Q.; Zhang, X.; Feng, Z.; et al. Efficacy and safety of three new oral antiviral treatment (molnupiravir, fluvoxamine and Paxlovid) for COVID-19: A meta-analysis. Ann. Med. 2022, 54, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Vitiello, A.; Troiano, V.; La Porta, R. What will be the role of molnupiravir in the treatment of COVID-19 infection? Drugs Ther. Perspect. 2021, 37, 579–580. [Google Scholar] [CrossRef] [PubMed]
- WHO Solidarity Trial Consortium. Remdesivir and three other drugs for hospitalised patients with COVID-19: Final results of the WHO Solidarity randomised trial and updated meta-analyses. Lancet 2022, 399, 1941–1953. [Google Scholar] [CrossRef]
- Kabinger, F.; Stiller, C.; Schmitzová, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Höbartner, C.; Cramer, P. Mechanism of molnupiravir-induced SARS-CoV-2 mutagenesis. Nat. Struct. Mol. Biol. 2021, 28, 740–746. [Google Scholar] [CrossRef]
- Gandhi, S.; Klein, J.; Robertson, A.J.; Peña-Hernández, M.A.; Lin, M.J.; Roychoudhury, P.; Lu, P.; Fournier, J.; Ferguson, D.; Mohamed Bakhash, S.A.K.; et al. De novo emergence of a remdesivir resistance mutation during treatment of persistent SARS-CoV-2 infection in an immunocompromised patient: A case report. Nat. Commun. 2022, 13, 1547. [Google Scholar] [CrossRef]
- Najjar-Debbiny, R.; Gronich, N.; Weber, G.; Khoury, J.; Amar, M.; Stein, N.; Goldstein, L.H.; Saliba, W. Effectiveness of Paxlovid in Reducing Severe Coronavirus Disease 2019 and Mortality in High-Risk Patients. Clin. Infect. Dis. 2023, 76, e342–e349. [Google Scholar] [CrossRef]
- Hammond, J.; Leister-Tebbe, H.; Gardner, A.; Abreu, P.; Bao, W.; Wisemandle, W.; Baniecki, M.; Hendrick, V.M.; Damle, B.; Simón-Campos, A.; et al. Oral Nirmatrelvir for High-Risk, Nonhospitalized Adults with COVID-19. N. Engl. J. Med. 2022, 386, 1397–1408. [Google Scholar] [CrossRef]
- Sacco, M.D.; Hu, Y.; Gongora, M.V.; Meilleur, F.; Kemp, M.T.; Zhang, X.; Wang, J.; Chen, Y. The P132H mutation in the main protease of Omicron SARS-CoV-2 decreases thermal stability without compromising catalysis or small-molecule drug inhibition. Cell Res. 2022, 32, 498–500. [Google Scholar] [CrossRef]
- Mótyán, J.A.; Mahdi, M.; Hoffka, G.; Tőzsér, J. Potential Resistance of SARS-CoV-2 Main Protease (Mpro) against Protease Inhibitors: Lessons Learned from HIV-1 Protease. Int. J. Mol. Sci. 2022, 23, 3507. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef]
- Lan, Q.; Wang, L.; Jiao, F.; Lu, L.; Xia, S.; Jiang, S. Pan-coronavirus fusion inhibitors to combat COVID-19 and other emerging coronavirus infectious diseases. J. Med. Virol. 2023, 95, e28143. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Yan, L.; Xu, W.; Agrawal, A.S.; Algaissi, A.; Tseng, C.K.; Wang, Q.; Du, L.; Tan, W.; Wilson, I.A.; et al. A pan-coronavirus fusion inhibitor targeting the HR1 domain of human coronavirus spike. Sci. Adv. 2019, 5, eaav4580. [Google Scholar] [CrossRef]
- Yu, F.; Pan, T.; Huang, F.; Ying, R.; Liu, J.; Fan, H.; Zhang, J.; Liu, W.; Lin, Y.; Yuan, Y.; et al. Glycopeptide Antibiotic Teicoplanin Inhibits Cell Entry of SARS-CoV-2 by Suppressing the Proteolytic Activity of Cathepsin L. Front. Microbiol. 2022, 13, 884034. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Pan, T.; Zhang, J.; Li, Q.; Zhang, X.; Bai, C.; Huang, F.; Peng, T.; Zhang, J.; Liu, C.; et al. Glycopeptide Antibiotics Potently Inhibit Cathepsin L in the Late Endosome/Lysosome and Block the Entry of Ebola Virus, Middle East Respiratory Syndrome Coronavirus (MERS-CoV), and Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV). J. Biol. Chem. 2016, 291, 9218–9232. [Google Scholar] [CrossRef]
- Zhao, M.M.; Zhu, Y.; Zhang, L.; Zhong, G.; Tai, L.; Liu, S.; Yin, G.; Lu, J.; He, Q.; Li, M.J.; et al. Novel cleavage sites identified in SARS-CoV-2 spike protein reveal mechanism for cathepsin L-facilitated viral infection and treatment strategies. Cell Discov. 2022, 8, 53. [Google Scholar] [CrossRef]
- Song, W.; Gui, M.; Wang, X.; Xiang, Y. Cryo-EM structure of the SARS coronavirus spike glycoprotein in complex with its host cell receptor ACE2. PLoS Pathog. 2018, 14, e1007236. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.M.; Yang, W.L.; Yang, F.Y.; Zhang, L.; Huang, W.J.; Hou, W.; Fan, C.F.; Jin, R.H.; Feng, Y.M.; Wang, Y.C.; et al. Cathepsin L plays a key role in SARS-CoV-2 infection in humans and humanized mice and is a promising target for new drug development. Signal Transduct. Target. Ther. 2021, 6, 134. [Google Scholar] [CrossRef]
- Simmons, G.; Zmora, P.; Gierer, S.; Heurich, A.; Pöhlmann, S. Proteolytic activation of the SARS-coronavirus spike protein: Cutting enzymes at the cutting edge of antiviral research. Antivir. Res. 2013, 100, 605–614. [Google Scholar] [CrossRef]
- McCray, P.B., Jr.; Pewe, L.; Wohlford-Lenane, C.; Hickey, M.; Manzel, L.; Shi, L.; Netland, J.; Jia, H.P.; Halabi, C.; Sigmund, C.D.; et al. Lethal infection of K18-hACE2 mice infected with severe acute respiratory syndrome coronavirus. J. Virol. 2007, 81, 813–821. [Google Scholar] [CrossRef]
- Muralidar, S.; Gopal, G.; Visaga Ambi, S. Targeting the viral-entry facilitators of SARS-CoV-2 as a therapeutic strategy in COVID-19. J. Med. Virol. 2021, 93, 5260–5276. [Google Scholar] [CrossRef]
- Pu, J.; Zhou, J.T.; Liu, P.; Yu, F.; He, X.; Lu, L.; Jiang, S. Viral Entry Inhibitors Targeting Six-Helical Bundle Core against Highly Pathogenic Enveloped Viruses with Class I Fusion Proteins. Curr. Med. Chem. 2022, 29, 700–718. [Google Scholar] [CrossRef]
- Li, L.; He, L.; Tan, S.; Guo, X.; Lu, H.; Qi, Z.; Pan, C.; An, X.; Jiang, S.; Liu, S. 3-hydroxyphthalic anhydride-modified chicken ovalbumin exhibits potent and broad anti-HIV-1 activity: A potential microbicide for preventing sexual transmission of HIV-1. Antimicrob. Agents Chemother. 2010, 54, 1700–1711. [Google Scholar] [CrossRef]
- Li, M.; Duan, J.; Qiu, J.; Yu, F.; Che, X.; Jiang, S.; Li, L. 3-hydroxyphthalic anhydride-modified human serum albumin as a microbicide candidate against HIV type 1 entry by targeting both viral envelope glycoprotein gp120 and cellular receptor CD4. AIDS Res. Hum. Retroviruses 2013, 29, 1455–1464. [Google Scholar] [CrossRef]
- Fu, Y.; Li, P.; Xu, W.; Liu, Z.; Wang, C.; Wang, Q.; Tang, J.; Li, W.; Lu, L.; Jiang, S. Chemically Modified Bovine β-Lactoglobulin as a Broad-Spectrum Influenza Virus Entry Inhibitor with the Potential to Combat Influenza Outbreaks. Viruses 2022, 14, 55. [Google Scholar] [CrossRef]
- Lu, L.; Yang, X.; Li, Y.; Jiang, S. Chemically modified bovine beta-lactoglobulin inhibits human papillomavirus infection. Microbes Infect. 2013, 15, 506–510. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, Q.; Jia, R.; Xia, S.; Li, Y.; Liu, Q.; Xu, W.; Xu, J.; Du, L.; Lu, L.; et al. Intranasal administration of maleic anhydride-modified human serum albumin for pre-exposure prophylaxis of respiratory syncytial virus infection. Viruses 2015, 7, 798–819. [Google Scholar] [CrossRef]
- Zhao, G.; Du, L.; Ma, C.; Li, Y.; Li, L.; Poon, V.K.; Wang, L.; Yu, F.; Zheng, B.J.; Jiang, S.; et al. A safe and convenient pseudovirus-based inhibition assay to detect neutralizing antibodies and screen for viral entry inhibitors against the novel human coronavirus MERS-CoV. Virol. J. 2013, 10, 266. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Qiu, J.; Niu, X.; Ma, Q.; Zhou, C.; Chen, P.; Zhang, Q.; Chen, M.; Yang, Z.; Liu, S.; et al. 3-Hydroxyphthalic Anhydride-Modified Chicken Ovalbumin as a Potential Candidate Inhibits SARS-CoV-2 Infection by Disrupting the Interaction of Spike Protein With Host ACE2 Receptor. Front. Pharmacol. 2020, 11, 603830. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, M.; Takashima, A.; Ono, C.; Okamoto, T.; Iwasaki, M. Phenothiazines inhibit SARS-CoV-2 cell entry via a blockade of spike protein binding to neuropilin-1. Antivir. Res. 2023, 209, 105481. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yu, D.; Yan, H.; Chong, H.; He, Y. Design of Potent Membrane Fusion Inhibitors against SARS-CoV-2, an Emerging Coronavirus with High Fusogenic Activity. J. Virol. 2020, 94, 10–1128. [Google Scholar] [CrossRef]
- Candurra, N.A.; Maskin, L.; Damonte, E.B. Inhibition of arenavirus multiplication in vitro by phenotiazines. Antivir. Res. 1996, 31, 149–158. [Google Scholar] [CrossRef]
- Hewlett, I.; Lee, S.; Molnar, J.; Foldeak, S.; Pine, P.S.; Weaver, J.L.; Aszalos, A. Inhibition of HIV infection of H9 cells by chlorpromazine derivatives. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol 1997, 15, 16–20. [Google Scholar] [CrossRef]
- Ochiai, H.; Kurokawa, M.; Niwayama, S. Influence of trifluoperazine on the late stage of influenza virus infection in MDCK cells. Antivir. Res. 1991, 15, 149–160. [Google Scholar] [CrossRef]
- Piccini, L.E.; Castilla, V.; Damonte, E.B. Inhibition of dengue virus infection by trifluoperazine. Arch. Virol. 2022, 167, 2203–2212. [Google Scholar] [CrossRef]
- Ruiz de Pellón Santamaría, Á. Psychosis Treatment During COVID-19 Pandemic and the Potential Role of Phenothiazines: A Call for Research Studies. J. Clin. Psychopharmacol. 2020, 40, 641–642. [Google Scholar] [CrossRef]
- Amaral, L.; Viveiros, M.; Molnar, J. Antimicrobial activity of phenothiazines. In Vivo 2004, 18, 725–731. [Google Scholar]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef]
- Koch, J.; Uckeley, Z.M.; Doldan, P.; Stanifer, M.; Boulant, S.; Lozach, P.Y. TMPRSS2 expression dictates the entry route used by SARS-CoV-2 to infect host cells. Embo J. 2021, 40, e107821. [Google Scholar] [CrossRef]
- Forrestall, K.L.; Burley, D.E.; Cash, M.K.; Pottie, I.R.; Darvesh, S. Phenothiazines as dual inhibitors of SARS-CoV-2 main protease and COVID-19 inflammation. Can. J. Chem. 2021, 99, 801–811. [Google Scholar] [CrossRef]
- Machado-Vieira, R.; Quevedo, J.; Shahani, L.; Soares, J.C. Convergent evidence for the antiviral effects of several FDA-approved phenothiazine antipsychotics against SARS-CoV-2 and other coronaviruses. Braz. J. Psychiatry 2021, 43, 462–464. [Google Scholar] [CrossRef]
- Plaze, M.; Attali, D.; Petit, A.C.; Blatzer, M.; Simon-Loriere, E.; Vinckier, F.; Cachia, A.; Chrétien, F.; Gaillard, R. Repurposing chlorpromazine to treat COVID-19: The reCoVery study. Encephale 2020, 46, 169–172. [Google Scholar] [CrossRef]
- Lu, J.; Hou, Y.; Ge, S.; Wang, X.; Wang, J.; Hu, T.; Lv, Y.; He, H.; Wang, C. Screened antipsychotic drugs inhibit SARS-CoV-2 binding with ACE2 in vitro. Life Sci. 2021, 266, 118889. [Google Scholar] [CrossRef]
- Varga, B.; Csonka, Á.; Csonka, A.; Molnár, J.; Amaral, L.; Spengler, G. Possible Biological and Clinical Applications of Phenothiazines. Anticancer. Res. 2017, 37, 5983–5993. [Google Scholar] [CrossRef]
- Jaszczyszyn, A.; Gąsiorowski, K.; Świątek, P.; Malinka, W.; Cieślik-Boczula, K.; Petrus, J.; Czarnik-Matusewicz, B. Chemical structure of phenothiazines and their biological activity. Pharmacol. Rep. 2012, 64, 16–23. [Google Scholar] [CrossRef]
- Galea, M.; Agius, M.; Vassallo, N. Neurological manifestations and pathogenic mechanisms of COVID-19. Neurol. Res. 2022, 44, 571–582. [Google Scholar] [CrossRef]
- Jaffry, M.; Faiz, I.; Jaffry, K.; Souayah, N. Neurological Manifestations of SARS-CoV-2 Infection and the Role of Complement Activation. US Neurol. 2022. [Google Scholar] [CrossRef]
- Chen, J.; Li, S.; Lei, Z.; Tang, Q.; Mo, L.; Zhao, X.; Xie, F.; Zi, D.; Tan, J. Inhibition of SARS-CoV-2 pseudovirus invasion by ACE2 protecting and Spike neutralizing peptides: An alternative approach to COVID19 prevention and therapy. Int. J. Biol. Sci. 2021, 17, 2957–2969. [Google Scholar] [CrossRef] [PubMed]
- Simanjuntak, Y.; Liang, J.-J.; Lee, Y.-L.; Lin, Y.-L. Repurposing of Prochlorperazine for Use Against Dengue Virus Infection. J. Infect. Dis. 2014, 211, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Scovino, A.M.; Dahab, E.C.; Vieira, G.F.; Freire-de-Lima, L.; Freire-de-Lima, C.G.; Morrot, A. SARS-CoV-2’s Variants of Concern: A Brief Characterization. Front. Immunol. 2022, 13, 834098. [Google Scholar] [CrossRef] [PubMed]
- Kaufer, A.M.; Theis, T.; Lau, K.A.; Gray, J.L.; Rawlinson, W.D. Laboratory biosafety measures involving SARS-CoV-2 and the classification as a Risk Group 3 biological agent. Pathology 2020, 52, 790–795. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 (μM) | CC50 (μM) | SI b |
|---|---|---|---|
| Perphenazine | 0.831 ± 0.120 | 18.120 ± 1.450 | 21.805 |
| Acepromazine maleate | 1.823 ± 0.230 | 93.300 ± 11.520 | 51.179 |
| Fluphenazine decanoate | 3.903 ± 1.090 | 61.570 ± 28.840 | 15.775 |
| Prochlorperazine maleate | 0.259 ± 0.048 | 85.160 ± 13.915 | 328.803 |
| Alimemazine hemitartrate | 0.477 ± 0.124 | 53.390 ± 10.945 | 111.929 |
| SARS-CoV-2 Variants | IC50 (μM, Mean ± SD) | ||||
|---|---|---|---|---|---|
| Perphenazine | Fluphenazine Decanoate | Acepromazine Maleate | Prochlorperazine Maleate | Alimemazine Hemitartrate | |
| B.1.1.7 (Alpha) | 1.315 ± 0.283 | 2.680 ± 0.370 | 2.580 ± 0.494 | 0.526 ± 0.103 | 0.580 ± 0.084 |
| B.1.351 (Beta) | 1.215 ± 0.225 | 1.736 ± 0.518 | 3.152 ± 0.602 | 0.293 ± 0.027 | 0.293 ± 0.059 |
| P.1 (Gamma) | 0.487 ± 0.169 | 6.214 ± 1.211 | 1.719 ± 0.348 | 0.762 ± 0.100 | 1.526 ± 0.339 |
| B.1.429 (Epsilon) | 0.802 ± 0.125 | 1.200 ± 0.425 | 4.605 ± 1.068 | 0.113 ± 0.034 | 0.342 ± 0.103 |
| B.1.526 (lota) | 0.315 ± 0.042 | 4.525 ± 2.615 | 1.298 ± 0.196 | 0.31 ± 0.045 | 1.040 ± 0.574 |
| B.1.617.1 (Kappa) | 0.343 ± 0.075 | 3.957 ± 0.762 | 0.661 ± 0.161 | 0.249 ± 0.057 | 0.609 ± 0.082 |
| B.1.617.2 (Delta) | 0.903 ± 0.211 | 9.318 ± 3.236 | 2.496 ± 0.583 | 0.609 ± 0.128 | 1.776 ± 0.660 |
| C.37 (Lambda) | 0.482 ± 0.059 | 2.270 ± 0.419 | 1.993 ± 0.576 | 0.423 ± 0.096 | 0.342 ± 0.097 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, T.; Xiao, S.; Wu, Z.; Lv, X.; Liu, S.; Hu, M.; Li, G.; Li, P.; Ma, X. Phenothiazines Inhibit SARS-CoV-2 Entry through Targeting Spike Protein. Viruses 2023, 15, 1666. https://doi.org/10.3390/v15081666
Liang T, Xiao S, Wu Z, Lv X, Liu S, Hu M, Li G, Li P, Ma X. Phenothiazines Inhibit SARS-CoV-2 Entry through Targeting Spike Protein. Viruses. 2023; 15(8):1666. https://doi.org/10.3390/v15081666
Chicago/Turabian StyleLiang, Taizhen, Shiqi Xiao, Ziyao Wu, Xi Lv, Sen Liu, Meilin Hu, Guojie Li, Peiwen Li, and Xiancai Ma. 2023. "Phenothiazines Inhibit SARS-CoV-2 Entry through Targeting Spike Protein" Viruses 15, no. 8: 1666. https://doi.org/10.3390/v15081666
APA StyleLiang, T., Xiao, S., Wu, Z., Lv, X., Liu, S., Hu, M., Li, G., Li, P., & Ma, X. (2023). Phenothiazines Inhibit SARS-CoV-2 Entry through Targeting Spike Protein. Viruses, 15(8), 1666. https://doi.org/10.3390/v15081666