
Characterizing the Pathogenicity and Immunogenicity of Simian Retrovirus Subtype 8 (SRV-8) Using SRV-8-Infected Cynomolgus Monkeys

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Verification of SRV Subtype via Polymerase Chain Reaction (PCR) in Euthanized Monkeys
2.3. Real-Time Quantitative PCR for the Detection of SRV-8 cDNA Copies in Euthanized Monkeys
2.4. Detection of SRV-8 Antibodies in Euthanized Monkeys
2.5. Haematoxylin and Eosin (HE) Staining
2.6. Collection of PBMCs for CPE Formation Analysis and Immunofluorescent Assay (IFA)
2.7. Detection of Viremia, Proviral DNA, and Antibodies of SRV-8 in 28 Live Monkeys
2.8. Routine Blood Examination
2.9. Flow Cytometry Analysis
2.10. Transcriptome Sequencing Analysis
2.11. Identification of the Differentially Expressed Genes (DEGs)
2.12. DEG-Based KEGG Enrichment Analysis
2.13. Statistical Analysis
3. Results
3.1. Identification and Stratification of SRV-8-Infected Cynomolgus Monkeys
3.2. Tissue Specificity and Pathology of SRV-8 Infection
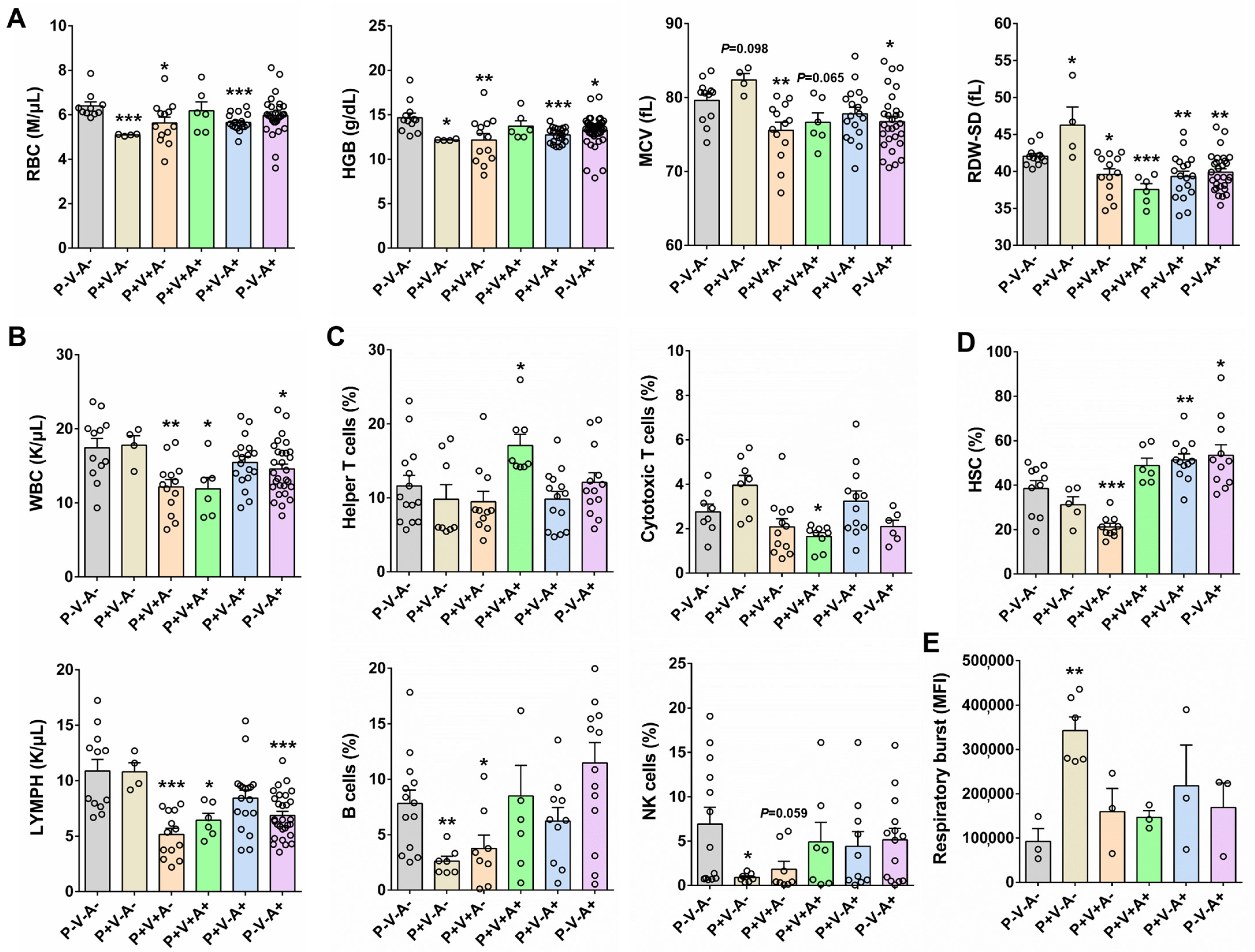
3.3. The Health Status and Cellular Immune Responses in SRV-8-Infected Monkeys
3.4. Gene Expression Profile of PBMCs in SRV-8-Infected Monkeys
3.5. KEGG Enrichment Analyses of DEGs in SRV-8-Infected Monkeys
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chopra, H.C.; Mason, M.M. A new virus in a spontaneous mammary tumor of a rhesus monkey. Cancer Res. 1970, 30, 2081–2086. [Google Scholar]
- Grant, R.; Keele, B.; Kuller, L.R.; Watanabe, R.; Perret, A.; Smedley, J. Identification of novel simian endogenous retroviruses that are indistinguishable from simian retrovirus (SRV) on current SRV diagnostic assays. J. Med. Primatol. 2017, 46, 158–161. [Google Scholar] [CrossRef]
- Nicholas, W.L. Simian retroviruses: Infection and disease—Implications for immunotoxicology research in primates. J. Immunotoxicol. 2010, 7, 93–101. [Google Scholar]
- Osborn, K.G.; Prahalada, S.; Lowenstine, L.J.; Gardner, M.B.; Maul, D.H.; Henrickson, R.V. The pathology of an epizootic of acquired immunodeficiency in rhesus macaques. Am. J. Pathol. 1984, 114, 94–103. [Google Scholar]
- Maul, D.H.; Lerche, N.W.; Osborn, K.G.; Marx, P.A.; Zaiss, C.; Spinner, A.; Kluge, J.D.; MacKenzie, M.R.; Lowenstine, L.J.; Bryant, M.L. Pathogenesis of simian AIDS in rhesus macaques inoculated with the SRV-1 strain of type D retrovirus. Am. J. Vet. Res. 1986, 47, 863–868. [Google Scholar]
- Yee, J.L.; Vanderford, T.H.; Didier, E.S.; Gray, S.; Lewis, A.; Roberts, J.; Taylor, K.; Bohm, R.P. Specific pathogen free macaque colonies: A review of principles and recent advances for viral testing and colony management. J. Med. Primatol. 2016, 45, 55–78. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.F.; Malinak, C.J.; Wu, H.; Sabo, A.; Tsai, C.-C. PCR amplification and DNA sequencing of SRV-2 from archived tumor tissues. Virus Res. 1995, 36, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Klatzmann, D.; Barre-Sinoussi, F.; Nugeyre, M.T.; Danquet, C.; Vilmer, E.; Griscelli, C.; Brun-Veziret, F.; Rouzioux, C.; Gluckman, J.C.; Chermann, J.C. Selective tropism of lymphadenopathy associated virus (LAV) for helper-inducer T lymphocytes. Science 1984, 225, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Zao, C.; Tomanek, L.; Cooke, A.; Berger, R.; Estep, J.S.; Armstrong, K.; Yee, J.L.; Lerche, N.W. An Unusual SRV/D-T, SRV-4-Like Outbreak in Cynomolgus Monkeys in the United States. J. Am. Assoc. Lab. Anim. 2009, 48, 559. [Google Scholar]
- Koide, R.; Yoshikawa, R.; Okamoto, M.; Sakaguchi, S.; Suzuki, J.; Isa, T.; Nakagawa, S.; Sakawaki, H.; Miura, T.; Miyazawa, T. Experimental infection of Japanese macaques with simian retrovirus 5. J. Gen. Virol. 2019, 100, 266–277. [Google Scholar] [CrossRef]
- Zao, C.L.; Tomanek, L.; Cooke, A.; Berger, R.; Yang, L.Y.; Xie, C.; Chen, S.; Shi, C.J.; Rong, R. A novel simian retrovirus subtype discovered in cynomolgus monkeys (Macaca fascicularis). J. Gen. Virol. 2016, 97, 3017–3023. [Google Scholar] [CrossRef]
- Irigoyen, N.; Firth, A.E.; Jones, J.D.; Chung, B.Y.W.; Siddell, S.G.; Brierley, I. High-Resolution Analysis of Coronavirus Gene Expression by RNA Sequencing and Ribosome Profiling. PLoS Patho. 2016, 12, e1005473. [Google Scholar] [CrossRef]
- Stern-Ginossar, N. Decoding Viral Infection by Ribosome Profiling. J. Virol. 2015, 89, 6164–6166. [Google Scholar] [CrossRef]
- Tokuyama, M.; Kong, Y.; Song, E.; Jayewickreme, T.; Kang, I.; Iwasaki, A. ERVmap analysis reveals genome-wide transcription of human endogenous retroviruses. Proc. Natl. Acad. Sci. USA 2018, 115, 12565–12572. [Google Scholar] [CrossRef]
- Mautino, M.R.; Ramsey, W.J.; Reiser, J.; Morgan, R.A. Modified human immunodeficiency virus-based lentiviral vectors display decreased sensitivity to trans-dominant rev. Hum. Gene Ther. 2000, 11, 895–908. [Google Scholar] [CrossRef]
- Indraccolo, S.; Feroli, F.; Minuzzo, S.; Mion, M.; Rosato, A.; Zamarchi, R.; Titti, F.; Verani, P.; Amadori, A.; Chieco-Bianchi, L. DNA immunization of mice against SIVmac239 Gag and Env using Rev-independent expression plasmids. Aids Res. Hum. Retrov. 1998, 14, 83–90. [Google Scholar] [CrossRef]
- Yasutomi, Y. Establishment of specific pathogen-free macaque colonies in Tsukuba Primate Research Center of Japan for AIDS research. Vaccine 2010, 28, B75–B77. [Google Scholar] [CrossRef] [PubMed]
- He, M.G.; Zheng, K.; Tan, D.; Wang, Z.X. Association between ERCC1 and ERCC2 gene polymorphisms and susceptibility to pancreatic cancer. Genet. Mol. Res. 2016, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.D.; Vega-Rodriguez, J.; Diabate, A.; Liu, J.N.; Cui, C.L.; Nignan, C.; Dong, L.; Li, F.; Ouedrago, C.O.; Bandaogo, A.M.; et al. Clock genes and environmental cues coordinate Anopheles pheromone synthesis, swarming, and mating. Science 2021, 371, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.Q.; Yang, X.W.; Hou, F.J.; Yu, X.F.; Wang, Q.Y.; Oh, H.S.; Raja, P.; Pesola, J.M.; Vanni, E.A.H.; McCarron, S.; et al. Regulation of host and virus genes by neuronal miR-138 favours herpes simplex virus 1 latency. Nat. Microbiol. 2021, 6, 682–696. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Feldman, A.T.; Wolfe, D. Tissue Processing and Hematoxylin and Eosin Staining. Histopathol. Methods Protoc. 2014, 1180, 31–43. [Google Scholar]
- Chen, H.; Schurch, C.M.; Noble, K.; Kim, K.; Krutzik, P.O.; O’Donnell, E.; Vander Tuig, J.; Nolan, G.P.; McIlwain, D.R. Functional comparison of PBMCs isolated by Cell Preparation Tubes (CPT) vs. Lymphoprep Tubes. BMC Immunol. 2020, 21, 15. [Google Scholar] [CrossRef]
- Wang, T.E.; Chao, T.L.; Tsai, H.T.; Line, P.H.; Tsai, Y.L.; Chang, S.Y. Differentiation of Cytopathic Effects (CPE) induced by influenza virus infection using deep Convolutional Neural Networks (CNN). PLoS Comput. Biol. 2020, 16, e1007883. [Google Scholar] [CrossRef]
- Zao, C.L.; Armstrong, K.; Tomanek, L.; Cooke, A.; Berger, R.; Estep, J.S.; Marx, P.A.; Trask, J.S.; Smith, D.G.; Yee, J.L.; et al. The complete genome and genetic characteristics of SRV-4 isolated from cynomolgus monkeys (Macaca fascicularis). Virology 2010, 405, 390–396. [Google Scholar] [CrossRef]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstrale, M.; Laurila, E.; et al. PGC-1 alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Chen, Y.G.; Lu, S.S.; Zhang, Y.; Yu, J.M.; Deng, L.Q.; Chen, H.; Zhang, Y.J.; Zhou, N.J.; Yuan, K.; Yu, L.Z.; et al. TLR2 agonist Pam3CSK4 enhances the antibacterial functions of GM-CSF induced neutrophils to methicillin-resistant Staphylococcus aureus. Microb. Pathog. 2019, 130, 204–212. [Google Scholar] [CrossRef]
- Kumar, A.; Kankainen, M.; Parsons, A.; Kallioniemi, O.; Mattila, P.; Heckman, C.A. The impact of RNA sequence library construction protocols on transcriptomic profiling of leukemia. BMC Genom. 2017, 18, 629. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef]
- Tang, Y.; Sun, Y.J.; Zhao, L.M.; Xu, X.J.; Huang, L.X.; Qin, Y.X.; Su, Y.Q.; Yi, G.F.; Yan, Q.P. Mechanistic insight into the roles of Pseudomonas plecoglossicida clpV gene in host-pathogen interactions with Larimichthys crocea by dual RNA-seq. FIish Shellfish Immun. 2019, 93, 344–353. [Google Scholar] [CrossRef]
- McRae, M. HIV and viral protein effects on the blood brain barrier. Tissue Barriers 2016, 4, e1143543. [Google Scholar] [CrossRef] [PubMed]
- Guzman, R.E.; Kerlin, R.L.; Zimmerman, T.E. Histologic lesions in cynomolgus monkeys (Macaca fascicularis) naturally infected with simian retrovirus type D: Comparison of seropositive, virus-positive, and uninfected animals. Toxicol. Pathol. 1999, 27, 672–677. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, R.; Okamoto, M.; Sakaguchi, S.; Nakagawa, S.; Miura, T.; Hirai, H.; Miyazawa, T. Simian Retrovirus 4 Induces Lethal Acute Thrombocytopenia in Japanese Macaques. J. Virol. 2015, 89, 3965–3975. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Kobayashi, T.; Misawa, N.; Yoshikawa, R.; Takeuchi, J.S.; Miura, T.; Okamoto, M.; Yasunaga, J.I.; Matsuoka, M.; Ito, M.; et al. Experimental evaluation of the zoonotic infection potency of simian retrovirus type 4 using humanized mouse model. Sci. Rep. 2015, 5, 14040. [Google Scholar] [CrossRef]
- Banerjee, P.; Crawford, L.; Samuelson, E.; Feuer, G. Hematopoietic stem cells and retroviral infection. Retrovirology 2010, 7, 8. [Google Scholar] [CrossRef]
- Zhu, J.T.; Yang, L.Y.; Zhang, Q.B.; Meng, J.; Lu, Z.L.; Rong, R. Autophagy Induced by Simian Retrovirus Infection Controls Viral Replication and Apoptosis of Jurkat T Lymphocytes. Viruses 2020, 12, 381. [Google Scholar] [CrossRef]
- Sun, L.N.; Middleton, D.R.; Wantuch, P.L.; Ozdilek, A.; Avci, F.Y. Carbohydrates as T-cell antigens with implications in health and disease. Glycobiology 2016, 26, 1029–1040. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.; Pearce, E.L. Amino Assests: How Amino Acids Support Immunity. Cell Metab. 2020, 32, 154–175. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotides Name | Sequence (5′-3′) | Description |
|---|---|---|
| SRV identification | ||
| SRV-1-F | GAGACAAATCTCCCTCCAGTGGTGACG | Forward primer for SRV-1 provirus |
| SRV-1-R | CAGCAATCTTCGGCTAATTGGCGTTGG | Reverse primer for SRV-1 provirus |
| SRV-2-F | GCGAGGCTATTACTATGATACACCAGC | Forward primer for SRV-2 provirus |
| SRV-2-R | CTTTAGGATTCCAGCAAATGGGCTGACC | Reverse primer for SRV-2 provirus |
| SRV-4-F | GTTAGTAATGTTCCTACAGTCATAGGATCAGG | Forward primer for SRV-4 provirus |
| SRV-4-R | GGTAAGGTTTTCATAGGTGTAATTACTGGGTAAGG | Reverse primer for SRV-4 provirus |
| SRV-5-F | GCAATGGTACAACTTATAATACAGCTAAATTGC | Forward primer for SRV-5 provirus |
| SRV-5-R | GTCAGATGCATTGGCCAAAGATAAATTTTGG | Reverse primer for SRV-5 provirus |
| SRV-8-F | CAGCTTACTCCCAAGTAGGTTCC | Forward primer for SRV-8 provirus |
| SRV-8-R | GAGATAGATATGTTATAAGTGTTAGTACAATTGG | Reverse primer for SRV-8 provirus |
| GAPDH-F | GGATATTGTTGCCATCAATGACC | Forward primer for GAPDH |
| GAPDH-R | CCTTCTCCGTGGTGGTGAAGAC | Reverse primer for GAPDH |
| qPCR | ||
| qRT-mk-GAPDH-F | GAGTCCACTGGCGTCTTCA | Forward qRT-PCR primer for GAPDH |
| qRT-mk-GAPDH-R | TCTTGAGGCTGTTGTCATACTTC | Reverse qRT-PCR primer for GAPDH |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Yang, Y.; Li, Y.; Lu, Y.; Gao, C.; Bian, X.; Liu, Z.; Sun, Q. Characterizing the Pathogenicity and Immunogenicity of Simian Retrovirus Subtype 8 (SRV-8) Using SRV-8-Infected Cynomolgus Monkeys. Viruses 2023, 15, 1538. https://doi.org/10.3390/v15071538
Xu L, Yang Y, Li Y, Lu Y, Gao C, Bian X, Liu Z, Sun Q. Characterizing the Pathogenicity and Immunogenicity of Simian Retrovirus Subtype 8 (SRV-8) Using SRV-8-Infected Cynomolgus Monkeys. Viruses. 2023; 15(7):1538. https://doi.org/10.3390/v15071538
Chicago/Turabian StyleXu, Libing, Yunpeng Yang, Yandong Li, Yong Lu, Changshan Gao, Xinyan Bian, Zongping Liu, and Qiang Sun. 2023. "Characterizing the Pathogenicity and Immunogenicity of Simian Retrovirus Subtype 8 (SRV-8) Using SRV-8-Infected Cynomolgus Monkeys" Viruses 15, no. 7: 1538. https://doi.org/10.3390/v15071538
APA StyleXu, L., Yang, Y., Li, Y., Lu, Y., Gao, C., Bian, X., Liu, Z., & Sun, Q. (2023). Characterizing the Pathogenicity and Immunogenicity of Simian Retrovirus Subtype 8 (SRV-8) Using SRV-8-Infected Cynomolgus Monkeys. Viruses, 15(7), 1538. https://doi.org/10.3390/v15071538