
Serological Survey of Retrovirus and Coronavirus Infections, including SARS-CoV-2, in Rural Stray Cats in The Netherlands, 2020–2022

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
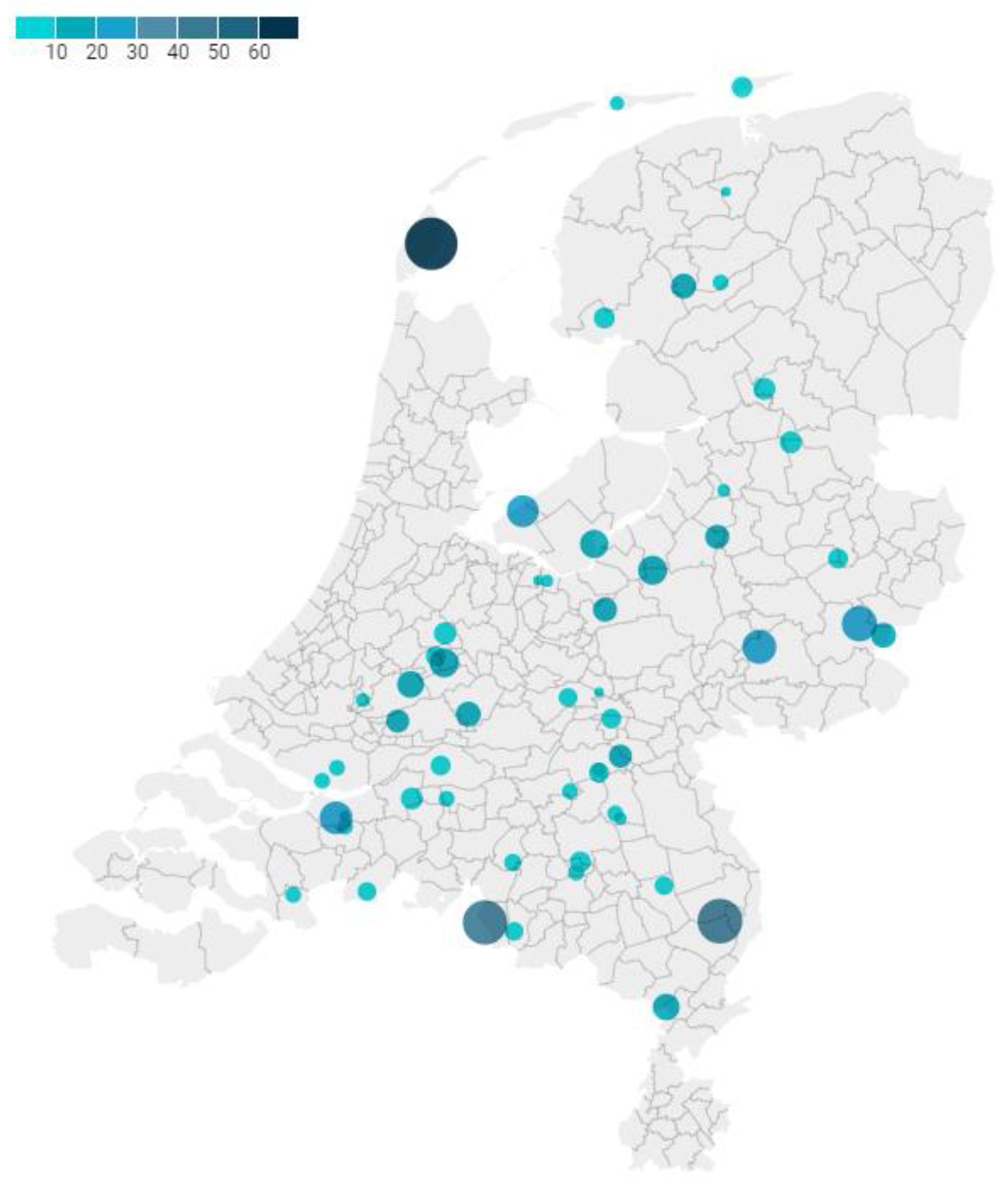
2.1. Study Population, Data Collection and Sampling
2.2. Ethics
2.3. Virus Detection
2.4. Antibody Detection
2.5. Data Analysis
3. Results
3.1. Characteristics of the Study Population and Samples
3.2. (Sero)prevalence Retroviruses and Coronaviruses in Stray Cats
3.3. FIV Seroprevalence and Associations with Age, Sex, Health Status, Location Type, Sample Size and Seropositivity for Other Viruses
3.4. FCoV Seroprevalence and Associations with Age, Sex, Health Status, Location Type, Sample Size and Seropositivity for Other Viruses
3.5. SARS-CoV-2 Suspected Seropositivity and Associations with Age, Sex, Health Status, Location Type, Sample Size and Seropositivity for Other Viruses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gerhold, R.W.; Jessup, D.A. Zoonotic Diseases Associated with Free-Roaming Cats: Zoonoses and Free-Roaming Cats. Zoonoses Public Health 2013, 60, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Neijenhuis, F.; van Niekerk, T. Als de kat van Huis Is…Zwerfkatten in Nederland, een Inventarisatie; WUR Wetenschapswinkel: Wageningen, The Netherlands, 2015. [Google Scholar]
- Trendpanel Gezelschapsdieren. Pet Monitor 2021 De Gezelschapsdierensector in 2030; Aeres Hogeschool Dronten/Has Hogeschool Den Bosch: Dronten/Den Bosch, The Netherlands, 2021. [Google Scholar]
- Sparkes, A.H.; Bessant, C.; Cope, K.; Ellis, S.L.H.; Finka, L.; Halls, V.; Hiestand, K.; Horsford, K.; Laurence, C.; MacFarlaine, I.; et al. ISFM Guidelines on Population Management and Welfare of Unowned Domestic Cats (Felis catus). J. Feline Med. Surg. 2013, 15, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Vitale, K.R. The Social Lives of Free-Ranging Cats. Animals 2022, 12, 126. [Google Scholar] [CrossRef]
- Liberg, O.; Sandell, M.; Pontier, D.; Natoli, E. Density, Spatial Organisation and Reproductive Tactics in the Domestic Cat and Other Felids. In The Domestic Cat: The biology of Its Behavior, 2nd ed.; Cambridge University Press: Cambridge, UK, 2000; pp. 119–148. ISBN 978-0-521-63648-3. [Google Scholar]
- Natoli, E.; Say, L.; Cafazzo, S.; Bonanni, R.; Schmid, M.; Pontier, D. Bold Attitude Makes Male Urban Feral Domestic Cats More Vulnerable to Feline Immunodeficiency Virus. Neurosci. Biobehav. Rev. 2005, 29, 151–157. [Google Scholar] [CrossRef]
- Candela, M.G.; Fanelli, A.; Carvalho, J.; Serrano, E.; Domenech, G.; Alonso, F.; Martínez-Carrasco, C. Urban Landscape and Infection Risk in Free-Roaming Cats. Zoonoses Public Health 2022, 69, 295–311. [Google Scholar] [CrossRef] [PubMed]
- Dorny, P.; Speybroeck, N.; Verstraete, S.; Baeke, M.; De Becker, A.; Berkvens, D.; Vercruysse, J. Serological Survey Toxoplasma Gondii of a on Feline Immunodeficiency Virus and Feine Leukaemia Virus in Urban Stray Cats in Belgium. Vet. Rec. 2002, 151, 626–629. [Google Scholar] [CrossRef] [PubMed]
- Montoya, A.; García, M.; Gálvez, R.; Checa, R.; Marino, V.; Sarquis, J.; Barrera, J.P.; Rupérez, C.; Caballero, L.; Chicharro, C.; et al. Implications of Zoonotic and Vector-Borne Parasites to Free-Roaming Cats in Central Spain. Vet. Parasitol. 2018, 251, 125–130. [Google Scholar] [CrossRef]
- Garigliany, M.; Jolly, S.; Dive, M.; Bayrou, C.; Berthemin, S.; Robin, P.; Godenir, R.; Petry, J.; Dahout, S.; Cassart, D.; et al. Risk Factors and Effect of Selective Removal on Retroviral Infections Prevalence in Belgian Stray Cats. Vet. Rec. 2016, 178, 45. [Google Scholar] [CrossRef]
- Spada, E.; Canzi, I.; Baggiani, L.; Perego, R.; Vitale, F.; Migliazzo, A.; Proverbio, D. Prevalence of Leishmania Infantum and Co-Infections in Stray Cats in Northern Italy. Comp. Immunol. Microbiol. Infect. Dis. 2016, 45, 53–58. [Google Scholar] [CrossRef]
- Muirden, A. Prevalence of Feline Leukaemia Virus and Antibodies to Feline Immunodeficiency Virus and Feline Coronavirus in Stray Cats Sent to an RSPCA Hospital. Vet. Rec. 2002, 150, 621–625. [Google Scholar] [CrossRef]
- Cave, T.A.; Golder, M.C.; Simpson, J.; Addie, D.D. Risk Factors for Feline Coronavirus Seropositivity in Cats Relinquished to a UK Rescue Charity. J. Feline Med. Surg. 2004, 6, 53–58. [Google Scholar] [CrossRef]
- Komiya, T.; Sadamasu, K.; Kang, M.-I.; Tsuboshima, S.; Fukushi, H.; Hirai, K. Seroprevalence of Coxiella Burnetii Infections among Cats in Different Living Environments. J. Vet. Med. Sci. 2003, 65, 1047–1048. [Google Scholar] [CrossRef] [PubMed]
- van Aart, A.E.; Velkers, F.C.; Fischer, E.A.J.; Broens, E.M.; Egberink, H.; Zhao, S.; Engelsma, M.; Hakze-van der Honing, R.W.; Harders, F.; de Rooij, M.M.T.; et al. SARS-CoV-2 Infection in Cats and Dogs in Infected Mink Farms. Transbound Emerg. Dis. 2022, 69, 3001–3007. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmeier, E.; Chan, T.; Klaus, J.; Pineroli, B.; Geisser, E.; Hofmann-Lehmann, R.; Meli, M.L. A Pre- and Within-Pandemic Survey of SARS-CoV-2 RNA in Saliva Swabs from Stray Cats in Switzerland. Viruses 2022, 14, 681. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Saz, S.; Martínez, M.; Giner, J.; González, A.; Tobajas, A.P.; Pérez, M.D.; Lira-Navarrete, E.; González-Ramírez, A.M.; Macías-León, J.; Verde, M.; et al. A Cross-Sectional Serosurvey of SARS-CoV-2 and Co-Infections in Stray Cats from the Second Wave to the Sixth Wave of COVID-19 Outbreaks in Spain. Vet. Res. Commun. 2023, 47, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Spada, E.; Vitale, F.; Bruno, F.; Castelli, G.; Reale, S.; Perego, R.; Baggiani, L.; Proverbio, D. A Pre- and during Pandemic Survey of Sars-Cov-2 Infection in Stray Colony and Shelter Cats from a High Endemic Area of Northern Italy. Viruses 2021, 13, 618. [Google Scholar] [CrossRef]
- Stranieri, A.; Lauzi, S.; Giordano, A.; Galimberti, L.; Ratti, G.; Decaro, N.; Brioschi, F.; Lelli, D.; Gabba, S.; Amarachi, N.L.; et al. Absence of SARS-CoV-2 RNA and Anti-SARS-CoV-2 Antibodies in Stray Cats. Transbound Emerg. Dis. 2022, 69, 2089–2095. [Google Scholar] [CrossRef]
- Duarte, A.; Castro, I.; Pereira da Fonseca, I.M.; Almeida, V.; Madeira de Carvalho, L.M.; Meireles, J.; Fazendeiro, M.I.; Tavares, L.; Vaz, Y. Survey of Infectious and Parasitic Diseases in Stray Cats at the Lisbon Metropolitan Area, Portugal. J. Feline Med. Surg. 2010, 12, 441–446. [Google Scholar] [CrossRef]
- Spada, E.; Proverbio, D.; Galluzzo, P.; Pepa, A.D.; Giorgi, G.B.D.; Perego, R.; Ferro, E. Prevalence of Haemoplasma Infections in Stray Cats in Northern Italy. ISRN Microbiol. 2014, 2014, 1–8. [Google Scholar] [CrossRef]
- Villanueva-Saz, S.; Giner, J.; Tobajas, A.P.; Pérez, M.D.; González-Ramírez, A.M.; Macías-León, J.; González, A.; Verde, M.; Yzuel, A.; Hurtado-Guerrero, R.; et al. Serological Evidence of SARS-CoV-2 and Co-Infections in Stray Cats in Spain. Transbound Emerg. Dis. 2021, 69, 1056–1064. [Google Scholar] [CrossRef]
- Taetzsch, S.J.; Bertke, A.S.; Gruszynski, K.R. Zoonotic Disease Transmission Associated with Feral Cats in a Metropolitan Area: A Geospatial Analysis. Zoonoses Public Health 2018, 65, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Kreisler, R.E.; Cornell, H.N.; Levy, J.K. Decrease in Population and Increase in Welfare of Community Cats in a Twenty-Three Year Trap-Neuter-Return Program in Key Largo, FL: The ORCAT Program. Front. Vet. Sci. 2019, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Hosie, M.J.; Addie, D.; Belák, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Lloret, A.; Lutz, H.; et al. Feline Immunodeficiency. ABCD Guidelines on Prevention and Management. J. Feline Med. Surg. 2009, 11, 575–584. [Google Scholar] [CrossRef]
- Studer, N.; Lutz, H.; Saegerman, C.; Gönczi, E.; Meli, M.L.; Boo, G.; Hartmann, K.; Hosie, M.J.; Moestl, K.; Tasker, S.; et al. Pan-European Study on the Prevalence of the Feline Leukaemia Virus Infection—Reported by the European Advisory Board on Cat Diseases (ABCD Europe). Viruses 2019, 11, 993. [Google Scholar] [CrossRef]
- Courchamp, F.; Suppo, C.; Fromont, E.; Bouloux, C. Dynamics of Two Feline Retroviruses (FIV and FeLV) within One Population of Cats. Proc. Biol. Sci. 1997, 264, 785–794. [Google Scholar] [CrossRef]
- Lutz, H.; Addie, D. Belak Feline Leukaemia Abcd Guidelines on Prevention and Management. J. Feline Med. Surg. 2009, 11, 529. [Google Scholar] [CrossRef]
- Hartmann, K.; Griessmayr, P.; Schulz, B.; Greene, C.E.; Vidyashankar, A.N.; Jarrett, O.; Egberink, H.F. Quality of Different In-Clinic Test Systems for Feline Immunodeficiency Virus and Feline Leukaemia Virus Infection. J. Feline Med. Surg. 2007, 9, 439–445. [Google Scholar] [CrossRef]
- Nichols, J.; Weng, H.Y.; Litster, A.; Leutenegger, C.; Guptill, L. Commercially Available Enzyme-Linked Immunosorbent Assay and Polymerase Chain Reaction Tests for Detection of Feline Immunodeficiency Virus Infection. J. Vet. Intern. Med. 2017, 31, 55–59. [Google Scholar] [CrossRef]
- Hartmann, K. Feline Immunodeficiency Virus Infection: An Overview. Vet. J. 1998, 155, 123–137. [Google Scholar] [CrossRef]
- Novacco, M.; Kohan, N.R.; Stirn, M.; Meli, M.L. Prevalence, Geographic Distribution, Risk Factors and Co-Infections of Feline Gammaherpesvirus Infections in Domestic Cats in Switzerland. Viruses 2019, 11, 721. [Google Scholar] [CrossRef]
- Kokkinaki, K.G.; Saridomichelakis, M.N.; Leontides, L.; Mylonakis, M.E.; Konstantinidis, A.O.; Steiner, J.M.; Suchodolski, J.S.; Xenoulis, P.G. A Prospective Epidemiological, Clinical, and Clinicopathologic Study of Feline Leukemia Virus and Feline Immunodeficiency Virus Infection in 435 Cats from Greece. Comp. Immunol. Microbiol. Infect. Dis. 2021, 78, 101687. [Google Scholar] [CrossRef] [PubMed]
- Niels, P.; Claire, A. Leslie Lyons Pathogenesis of Feline Enteric Coronavirus Infection. J. Feline Med. Surg. 2008, 10, 529–541. [Google Scholar] [CrossRef]
- Felten, S.; Klein-Richers, U.; Hofmann-Lehmann, R.; Bergmann, M.; Unterer, S.; Leutenegger, C.M.; Hartmann, K. Correlation of Feline Coronavirus Shedding in Feces with Coronavirus Antibody Titer. Pathogens 2020, 9, 598. [Google Scholar] [CrossRef] [PubMed]
- Poland, A.M.; Vennema, H.; Foley, J.E.; Pedersen, N.C. Two Related Strains of Feline Infectious Peritonitis Virus Isolated from Immunocompromised Cats Infected with a Feline Enteric Coronavirus. J. Clin. Microbiol. 1996, 34, 5. [Google Scholar] [CrossRef] [PubMed]
- Klein-Richers, U.; Hartmann, K.; Hofmann-Lehmann, R.; Unterer, S.; Bergmann, M.; Rieger, A.; Leutenegger, C.; Pantchev, N.; Balzer, J.; Felten, S. Prevalence of Feline Coronavirus Shedding in German Catteries and Associated Risk Factors. Viruses 2020, 12, 1000. [Google Scholar] [CrossRef] [PubMed]
- Addie, D.; Belák, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A.; Lutz, H.; et al. Feline Infectious Peritonitis. ABCD Guidelines on Prevention and Management. J. Feline Med. Surg. 2009, 11, 594–604. [Google Scholar] [CrossRef]
- Grieco, V.; Crepaldi, P.; Giudice, C.; Roccabianca, P.; Sironi, G.; Brambilla, E.; Magistrelli, S.; Ravasio, G.; Granatiero, F.; Invernizzi, A.; et al. Causes of Death in Stray Cat Colonies of Milan: A Five-Year Report. Animals 2021, 11, 3308. [Google Scholar] [CrossRef]
- Möstl, K.; Addie, D.D.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A.; Lutz, H.; et al. Something Old, Something New: Update of the 2009 and 2013 ABCD Guidelines on Prevention and Management of Feline Infectious Diseases. J. Feline Med. Surg. 2015, 17, 570–582. [Google Scholar] [CrossRef]
- Barrs, V.R.; Peiris, M.; Tam, K.W.S.; Law, P.Y.T.; Brackman, C.J.; To, E.M.W.; Yu, V.Y.T.; Chu, D.K.W.; Perera, R.A.P.M.; Sit, T.H.C. SARS-CoV-2 in Quarantined Domestic Cats from COVID-19 Households or Close Contacts, Hong Kong, China. Emerg. Infect. Dis. 2020, 26, 3071–3074. [Google Scholar] [CrossRef]
- Kannekens-Jager, M.M.; de Rooij, M.M.T.; de Groot, Y.; Biesbroeck, E.; de Jong, M.K.; Pijnacker, T.; Smit, L.A.M.; Schuurman, N.; Broekhuizen-Stins, M.J.; Zhao, S.; et al. SARS-CoV-2 Infection in Dogs and Cats Is Associated with Contact to COVID-19-Positive Household Members. Transbound Emerg. Dis. 2022, 69, 4034–4040. [Google Scholar] [CrossRef]
- Alberto-Orlando, S.; Calderon, J.L.; Leon-Sosa, A.; Patiño, L.; Zambrano-Alvarado, M.N.; Pasquel-Villa, L.D.; Rugel-Gonzalez, D.O.; Flores, D.; Mera, M.D.; Valencia, P.; et al. SARS-CoV-2 Transmission from Infected Owner to Household Dogs and Cats Is Associated with Food Sharing. Int. J. Infect. Dis. 2022, 122, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Neira, V.; Brito, B.; Agüero, B.; Berrios, F.; Valdés, V.; Gutierrez, A.; Ariyama, N.; Espinoza, P.; Retamal, P.; Holmes, E.C.; et al. A Household Case Evidences Shorter Shedding of SARS-CoV-2 in Naturally Infected Cats Compared to Their Human Owners. Emerg. Microbes Infect. 2021, 10, 376–383. [Google Scholar] [CrossRef]
- Hamer, S.A.; Pauvolid-Corrêa, A.; Zecca, I.B.; Davila, E.; Auckland, L.D.; Roundy, C.M.; Tang, W.; Torchetti, M.K.; Killian, M.L.; Jenkins-Moore, M.; et al. SARS-CoV-2 Infections and Viral Isolations among Serially Tested Cats and Dogs in Households with Infected Owners in Texas, USA. Viruses 2021, 13, 938. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.I.; Elia, G.; Grassi, A.; Giordano, A.; Desario, C.; Medardo, M.; Smith, S.L.; Anderson, E.R.; Prince, T.; Patterson, G.T.; et al. Evidence of Exposure to SARS-CoV-2 in Cats and Dogs from Households in Italy. Nat. Commun. 2020, 11, 6231. [Google Scholar] [CrossRef] [PubMed]
- Bienzle, D.; Rousseau, J.; Marom, D.; MacNicol, J.; Jacobson, L.; Sparling, S.; Prystajecky, N.; Fraser, E.; Weese, J.S. Risk Factors for SARS-CoV-2 Infection and Illness in Cats and Dogs. Emerg. Infect. Dis. 2022, 28, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Schuurman, N.; Li, W.; Wang, C.; Smit, L.A.M.; Broens, E.M.; Wagenaar, J.A.; van Kuppeveld, F.J.M.; Bosch, B.-J.; Egberink, H. Serologic Screening of Severe Acute Respiratory Syndrome Coronavirus 2 Infection in Cats and Dogs during First Coronavirus Disease Wave, The Netherlands. Emerg. Infect. Dis. 2021, 27, 1362–1370. [Google Scholar] [CrossRef] [PubMed]
- Adler, J.M.; Weber, C.; Wernike, K.; Michelitsch, A.; Friedrich, K.; Trimpert, J.; Beer, M.; Kohn, B.; Osterrieder, K.; Müller, E. Prevalence of Anti-Severe Acute Respiratory Syndrome Coronavirus 2 Antibodies in Cats in Germany and Other European Countries in the Early Phase of the Coronavirus Disease-19 Pandemic. Zoonoses Public Health 2022, 69, 439–450. [Google Scholar] [CrossRef]
- Bosco-Lauth, A.M.; Hartwig, A.E.; Porter, S.M.; Gordy, P.W.; Nehring, M.; Byas, A.D.; VandeWoude, S.; Ragan, I.K.; Maison, R.M.; Bowen, R.A. Experimental Infection of Domestic Dogs and Cats with SARS-CoV-2: Pathogenesis, Transmission, and Response to Reexposure in Cats. Proc. Natl. Acad. Sci. USA 2020, 117, 26382–26388. [Google Scholar] [CrossRef]
- Gerhards, N.M.; Gonzales, J.L.; Vreman, S.; Ravesloot, L.; van den Brand, J.M.A.; Doekes, H.P.; Egberink, H.F.; Stegeman, A.; Oreshkova, N.; van der Poel, W.H.M.; et al. Efficient Direct and Limited Environmental Transmission of SARS-CoV-2 Lineage B.1.22 in Domestic Cats. Microbiol. Spectr. 2023, 11, e02553-22. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of Ferrets, Cats, Dogs, and Other Domesticated Animals to SARS–Coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef]
- Martins, M.; do Nascimento, G.M.; Nooruzzaman, M.; Yuan, F.; Chen, C.; Caserta, L.C.; Miller, A.D.; Whittaker, G.R.; Fang, Y.; Diel, D.G. The Omicron Variant BA.1.1 Presents a Lower Pathogenicity than B.1 D614G and Delta Variants in a Feline Model of SARS-CoV-2 Infection. J. Virol. 2022, 96, e00961-22. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, J.L.; de Jong, M.C.M.; Gerhards, N.M.; Van der Poel, W.H.M. The SARS-CoV-2 Reproduction Number R0 in Cats. Viruses 2021, 13, 2480. [Google Scholar] [CrossRef] [PubMed]
- Cardillo, L.; de Martinis, C.; Brandi, S.; Levante, M.; Cozzolino, L.; Spadari, L.; Boccia, F.; Carbone, C.; Pompameo, M.; Fusco, G. SARS-CoV-2 Serological and Biomolecular Analyses among Companion Animals in Campania Region (2020–2021). Microorganisms 2022, 10, 263. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Bastit, L.; Marfil, S.; Pradenas, E.; Valle, R.; Roca, N.; Rodon, J.; Pailler-García, L.; Trinité, B.; Parera, M.; Noguera-Julian, M.; et al. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection and Humoral Responses against Different Variants of Concern in Domestic Pet Animals and Stray Cats from North-Eastern Spain. Transbound Emerg. Dis. 2022, 69, 3518–3529. [Google Scholar] [CrossRef]
- Statistics Netherlands. Available online: https://www.cbs.nl/en-gb/our-services/methods/definitions/rural-area (accessed on 1 May 2023).
- Diehl, K.-H.; Hull, R.; Morton, D.; Pfister, R.; Rabemampianina, Y.; Smith, D.; Vidal, J.-M.; Vorstenbosch, C.V.D. A Good Practice Guide to the Administration of Substances and Removal of Blood, Including Routes and Volumes. J. Appl. Toxicol. 2001, 21, 15–23. [Google Scholar] [CrossRef]
- Egberink, H.F.; Lutz, H.; Horzinek, M.C. Use of Western Blot and Radioimmunoprecipitation for Diagnosis of Feline Leukemia and Feline Immunodeficiency Virus Infections. J. Am. Vet. Med. Assoc. 1991, 199, 1339–1342. [Google Scholar]
- Zhao, S.; Li, W.; Schuurman, N.; van Kuppeveld, F.; Bosch, B.-J.; Egberink, H. Serological Screening for Coronavirus Infections in Cats. Viruses 2019, 11, 743. [Google Scholar] [CrossRef]
- Wang, C.; Li, W.; Drabek, D.; Okba, N.M.A.; van Haperen, R.; Osterhaus, A.D.M.E.; van Kuppeveld, F.J.M.; Haagmans, B.L.; Grosveld, F.; Bosch, B.-J. A Human Monoclonal Antibody Blocking SARS-CoV-2 Infection. Nat. Commun. 2020, 11, 2251. [Google Scholar] [CrossRef]
- Riepler, L.; Rössler, A.; Falch, A.; Volland, A.; Borena, W.; von Laer, D.; Kimpel, J. Comparison of Four SARS-CoV-2 Neutralization Assays. Vaccines 2021, 9, 13. [Google Scholar] [CrossRef]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.-C.; Tiu, C.; Hu, Z.; Chen, V.C.-W.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 Surrogate Virus Neutralization Test Based on Antibody-Mediated Blockage of ACE2–Spike Protein–Protein Interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef]
- Perera, R.A.; Ko, R.; Tsang, O.T.; Hui, D.; Ym, M.; Brackman, C.J.; To, E.M.W.; Yen, H.; Leung, K.; Cheng, S.; et al. Evaluation of a SARS-CoV-2 Surrogate Virus Neutralization Test for Detection of Antibody in Human, Canine, Cat and Hamster Sera. J. Clin. Microbiol. 2021, 59, e02504-20. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M.; Aisch, G.; Kokkelink, D. Datawrapper. Available online: https://www.datawrapper.de/ (accessed on 10 January 2022).
- Epitools—Sample Size to Estimate a Proportion or Apparent Prevalence with Specified Precision. Available online: https://epitools.ausvet.com.au/oneproportion (accessed on 20 June 2021).
- Ghebremariam, M.K.; Michel, A.L.; Nielen, M.; Vernooij, J.C.M.; Rutten, V.P.M.G. Farm-Level Risk Factors Associated with Bovine Tuberculosis in the Dairy Sector in Eritrea. Transbound. Emerg. Dis. 2018, 65, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Gleich, S.E.; Krieger, S.; Hartmann, K. Prevalence of Feline Immunodeficiency Virus and Feline Leukaemia Virus among Client-Owned Cats and Risk Factors for Infection in Germany. J. Feline Med. Surg. 2009, 11, 985–992. [Google Scholar] [CrossRef]
- Hendriks, P. Onderzoek naar Het Voorkomen van Het Feline Immunodeficientie Virus (Katte-Aids Virus) Bij de Nederlandse Asielkat; Rijksuniversiteit Utrecht: Utrecht, The Netherlands, 1993; unpublished manuscript. [Google Scholar]
- Egberink, H.F.; Hendriks, P.; Slappendel, R.J.; Horzinek, M.C. The Diagnosis and Epidemiology of Feline Immunodeficiency Virus (FIV) in The Netherlands. Tijdschrift voor Diergeneeskunde 1993, 118 (Suppl. S1), 2. [Google Scholar]
- Polak, K.C.; Levy, J.K.; Crawford, P.C.; Leutenegger, C.M.; Moriello, K.A. Infectious Diseases in Large-Scale Cat Hoarding Investigations. Vet. J. 2014, 201, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Mostl, K.; Egberink, H.; Addie, D.; Frymus, T.; Boucraut-Baralon, C.; Truyen, U.; Hartmann, K.; Lutz, H.; Gruffydd-Jones, T.; Radford, A.D.; et al. Prevention of Infectious Diseases in Cat Shelters: ABCD Guidelines. J. Feline Med. Surg. 2013, 15, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Martina, B.; Mirolo, M.; Müller, E.; Klein, R.; Volk, H.; Egberink, H.; Gonzalez-Hernandez, M.; Kaiser, F.; von Köckritz-Blickwede, M.; et al. SARS-CoV-2–Specific Antibodies in Domestic Cats during First COVID-19 Wave, Europe. Emerg. Infect. Dis. 2021, 27, 3115–3118. [Google Scholar] [CrossRef]
- van der Leij, W.J.R.; Broens, E.M.; Hesselink, J.W.; Schuurman, N.; Vernooij, J.C.M.; Egberink, H.F. Serological Screening for Antibodies against SARS-CoV-2 in Dutch Shelter Cats. Viruses 2021, 13, 1634. [Google Scholar] [CrossRef]
- Decaro, N.; Grassi, A.; Lorusso, E.; Patterson, E.I.; Lorusso, A.; Desario, C.; Anderson, E.R.; Vasinioti, V.; Wastika, C.E.; Hughes, G.L.; et al. Long-Term Persistence of Neutralizing SARS-CoV-2 Antibodies in Pets. Transbound Emerg. Dis. 2022, 69, 3073–3076. [Google Scholar] [CrossRef]
- WHO 2022 Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants (accessed on 6 January 2023).
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; COVID-19 Genomics UK (COG-UK) Consortium; et al. SARS-CoV-2 Variants, Spike Mutations and Immune Escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Sánchez-Morales, L.; Sánchez-Vizcaíno, J.M.; Pérez-Sancho, M.; Barroso-Arévalo, S. 1 The Omicron (B.1.1.529) SARS-CoV-2 Variant of Concern Also Affects 2 Companion Animals. Front. Vet. Sci. 2022, 9, 940710. [Google Scholar] [CrossRef]
- Pecora, A.; Malacari, D.A.; Mozgovoj, M.V.; de los Díaz, M.Á.; Peralta, A.V.; Cacciabue, M.; Puebla, A.F.; Carusso, C.; Mundo, S.L.; Gonzalez Lopez Ledesma, M.M.; et al. Anthropogenic Infection of Domestic Cats With SARS-CoV-2 Alpha Variant B.1.1.7 Lineage in Buenos Aires. Front. Vet. Sci. 2022, 9, 790058. [Google Scholar] [CrossRef] [PubMed]
- Barroso-Arévalo, S.; Sánchez-Morales, L.; Pérez-Sancho, M.; Domínguez, L.; Sánchez-Vizcaíno, J.M. First Detection of SARS-CoV-2 B.1.617.2 (Delta) Variant of Concern in a Symptomatic Cat in Spain. Front. Vet. Sci. 2022, 9, 841430. [Google Scholar] [CrossRef] [PubMed]
- Jairak, W.; Chamsai, E.; Udom, K.; Charoenkul, K.; Chaiyawong, S.; Techakriengkrai, N.; Tangwangvivat, R.; Suwannakarn, K.; Amonsin, A. SARS-CoV-2 Delta Variant Infection in Domestic Dogs and Cats, Thailand. Sci. Rep. 2022, 12, 8403. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Michelitsch, A.; Allendorf, V.; Conraths, F.J.; Beer, M.; Denzin, N.; Wernike, K. Dogs and Cats Are Less Susceptible to the Omicron Variant of Concern of SARS-CoV-2—A Field Study. bioRxiv 2023. [Google Scholar] [CrossRef]
- Hancock, T.J.; Hickman, P.; Kazerooni, N.; Kennedy, M.; Kania, S.A.; Dennis, M.; Szafranski, N.; Gerhold, R.; Su, C.; Masi, T.; et al. Possible Cross-Reactivity of Feline and White-Tailed Deer Antibodies against the SARS-CoV-2 Receptor Binding Domain. J. Virol. 2022, 96, e00250-22. [Google Scholar] [CrossRef]
- Yamamoto, J.K.; Edison, L.K.; Rowe-Haas, D.K.; Takano, T.; Gilor, C.; Crews, C.D.; Tuanyok, A.; Arukha, A.P.; Shiomitsu, S.; Walden, H.D.S.; et al. Feline Coronavirus Infection of Domestic Cats Causes Development of Cross-Reactive Antibodies to SARS-CoV-2 Receptor Binding Domain. bioRxiv 2022. [Google Scholar] [CrossRef]
- Michelitsch, A.; Hoffmann, D.; Wernike, K.; Beer, M. Occurrence of Antibodies against SARS-CoV-2 in the Domestic Cat Population of Germany. Vaccines 2020, 8, 772. [Google Scholar] [CrossRef] [PubMed]
- Frutos, R.; Serra-Cobo, J.; Pinault, L.; Lopez Roig, M.; Devaux, C.A. Emergence of Bat-Related Betacoronaviruses: Hazard and Risks. Front. Microbiol. 2021, 12, 591535. [Google Scholar] [CrossRef]
- Ancillotto, L.; Serangeli, M.T.; Russo, D. Curiosity Killed the Bat: Domestic Cats as Bat Predators. Mamm. Biol. 2013, 78, 369–373. [Google Scholar] [CrossRef]
- Sikkema, R.S.; Koopmans, M.P.G. Preparing for Emerging Zoonotic Viruses. Encycl. Virol. 2021, 256–266. [Google Scholar] [CrossRef]


{kind=link}
| Total # | FIV Positive | FIV Negative | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | Nr. | % | Nr. | % | Nr. | % | Relative Risk | 95% CI * | p-Value |
| Total | 580 | 100 | 29 | 5.0 | 551 | 95.0 | 3.4–7.1 | ||
| Estimated age | <0.001 | ||||||||
| <3 years | 440 | 78.3 | 12 | 2.7 | 428 | 97.3 | Ref | ||
| ≥3 years | 122 | 21.7 | 14 | 11.5 | 108 | 88.5 | 4.2 | 2.0–8.9 | |
| Sex | <0.001 | ||||||||
| Female | 320 | 55.8 | 5 | 1.6 | 315 | 98.4 | Ref | ||
| Male | 253 | 44.2 | 24 | 9.5 | 229 | 90.5 | 6.1 | 2.4–15.6 | |
| Health status | <0.001 | ||||||||
| Apparently | 392 | 76.0 | 9 | 2.3 | 383 | 97.7 | Ref | ||
| healthy | |||||||||
| Unhealthy | 124 | 24.0 | 14 | 11.3 | 110 | 88.7 | 4.9 | 2.2–11.1 | |
| Location type | 0.27 | ||||||||
| Dairy Farm | 294 | 55.3 | 17 | 5.8 | 277 | 94.2 | Ref | ||
| Industrial area | 32 | 6.0 | 2 | 6.3 | 30 | 93.8 | 1.1 | 0.26–4.5 | |
| Countryside | 78 | 14.7 | 6 | 7.7 | 72 | 92.3 | 1.2 | 0.50–3.0 | |
| residence | |||||||||
| Holidayparc/ campsite | 111 | 20.9 | 2 | 1.8 | 109 | 98.2 | 0.31 | 0.01–1.3 | |
| Nature reserve | 17 | 3.2 | 0 | 0.0 | 17 | 100 | 0.47 | 0.20–1.1 | |
| Sample size | 0.43 | ||||||||
| ≤10 | 202 | 34.8 | 8 | 4.0 | 194 | 96.0 | Ref | ||
| >10 | 378 | 65.2 | 21 | 5.6 | 357 | 94.4 | 1.4 | 0.6–3.1 | |
| Co-seropositivity | |||||||||
| FeLV neg | 578 | 100.0 | 29 | 5.0 | 549 | 95.0 | |||
| FeLV pos | 0 | 0.0 | 0 | 0.0 | 0 | ||||
| FCoV neg | 270 | 66.3 | 13 | 4.9 | 257 | 95.1 | Ref | 0.31 | |
| FCoV pos | 137 | 33.7 | 10 | 7.2 | 127 | 92.8 | 1.5 | 0.68–3.4 | |
| SARS-CoV-2 neg | 396 | 97.3 | 23 | 5.8 | 373 | 94.2 | Ref | 0.43 | |
| SARS-CoV-2 | 11 | 2.7 | 0 | 0.0 | 11 | 100.0 | 0.70 | 0.30–1.7 | |
| suspected | |||||||||
| Total # | FCoV Positive | FCoV Negative | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | Nr. | % | Nr. | % | Nr. | % | Relative Risk | 95% CI * | p-Value |
| Total | 407 | 100 | 137 | 33.7 | 270 | 66.3 | 29.1–38.5 | ||
| Estimated age | 0.002 | ||||||||
| <3 years | 304 | 75.4 | 90 | 29.6 | 214 | 70.4 | Ref | Ref | |
| ≥3 years | 99 | 24.6 | 46 | 46.5 | 53 | 53.5 | 1.6 | 1.2–2.1 | |
| Sex | 0.093 | ||||||||
| Female | 228 | 56.0 | 69 | 30.3 | 159 | 69.7 | Ref | Ref | |
| Male | 178 | 43.7 | 68 | 38.2 | 110 | 61.8 | 1.3 | 0.96–1.7 | |
| Health status | <0.001 | ||||||||
| Apparently healthy | 283 | 69.5 | 83 | 29.3 | 200 | 70.7 | Ref | Ref | |
| Unhealthy | 94 | 23.1 | 49 | 52.1 | 45 | 47.9 | 1.8 | 1.4–2.3 | |
| Location type | 0.018 | ||||||||
| Dairy Farm | 202 | 54.2 | 65 | 32.2 | 137 | 67.8 | Ref | Ref | |
| Industrial area | 18 | 4.8 | 10 | 55.6 | 8 | 44.4 | 1.7 | 1.1–2.7 | |
| Countryside residence | 59 | 15.8 | 27 | 45.8 | 32 | 54.2 | 1.4 | 1.0–2.0 | |
| Holiday parc/ campsite | 94 | 25.2 | 25 | 26.6 | 69 | 73.4 | 0.83 | 0.56–1.2 | |
| Nature reserve | 0 | 0 | NA | NA | NA | NA | NA | NA | |
| Sample size | 0.252 | ||||||||
| ≤10 | 119 | 29.2 | 35 | 29.4 | 84 | 70.6 | Ref | ||
| >10 | 288 | 70.8 | 102 | 35.4 | 186 | 64.6 | 1.2 | 0.87–1.7 | |
| Co-seropositivity | |||||||||
| FeLV neg | 407 | 100.0 | 137 | 33.7 | 270 | 66.3 | |||
| FeLV pos | 0 | 0.0 | 0 | 0.0 | 0 | 0 | |||
| FIV neg | 384 | 94.3 | 127 | 33.1 | 257 | 66.9 | Ref | ||
| FIV pos | 23 | 5.7 | 10 | 43.5 | 13 | 56.5 | 1.3 | 0.81–2.1 | 0.27 |
| SARS-CoV-2neg | 396 | 97.3 | 131 | 33.1 | 265 | 66.9 | Ref | ||
| SARS-CoV-2 suspected | 11 | 2.7 | 6 | 54.5 | 5 | 45.5 | 1.7 | 0.94–2.9 | 0.079 |
| Total # | SARS-CoV-2 Suspected | SARS-CoV-2 Negative | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | Nr. | % | Nr. | % | Nr. | % | Relative Risk | 95% CI * | p-Value |
| Total | 407 | 100 | 11 | 2.7 | 396 | 97.3 | 1.4–4.8 | ||
| Estimated age | 0.62 | ||||||||
| <3 years | 304 | 75.4 | 9 | 3.0 | 295 | 97.0 | Ref | ||
| ≥3 years | 99 | 24.6 | 2 | 2.0 | 97 | 98.0 | 0.68 | 0.15–3.1 | |
| Sex | 0.762 | ||||||||
| Female | 228 | 56.0 | 7 | 3.1 | 221 | 96.9 | Ref | ||
| Male | 178 | 43.7 | 4 | 2.2 | 174 | 97.8 | 0.73 | 0.22–2.5 | |
| Health status | 0.31 | ||||||||
| Apparently healthy | 283 | 69.5 | 10 | 3.5 | 273 | 96.5 | Ref | ||
| Unhealthy | 94 | 23.1 | 1 | 1.1 | 93 | 98.9 | 0.30 | 0.039–2.3 | |
| Location type | 0.91 | ||||||||
| Dairy Farm | 202 | 54.2 | 7 | 3.5 | 195 | 96.5 | Ref | ||
| Industrial area | 18 | 4.8 | 0 | 0.0 | 18 | 100 | 0.71 | 0.29–1.7 | |
| Countryside residence | 59 | 15.8 | 2 | 3.4 | 57 | 96.6 | 0.98 | 0.21–4.6 | |
| Holiday parc/ campsite | 94 | 25.2 | 2 | 2.1 | 92 | 97.9 | 0.61 | 0.13–2.9 | |
| Nature reserve | 0 | 0 | NA | NA | NA | NA | NA | ||
| Sample size | 0.52 | ||||||||
| ≤10 | 119 | 29.2 | 2 | 1.7 | 117 | 98.3 | Ref | ||
| >10 | 288 | 70.8 | 9 | 3.1 | 279 | 96.9 | 1.8 | 0.40–8.4 | |
| Co-seropositivity | |||||||||
| FeLV neg | 407 | 100.0 | 0 | 0.0 | 396 | 100.0 | |||
| FeLV pos | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | |||
| FIV neg | 384 | 94.3 | 11 | 2.9 | 373 | 97.1 | Ref | ||
| FIV pos | 23 | 5.7 | 0 | 0.0 | 23 | 100.0 | 0.70 | 0.29–1.7 | 0.43 |
| FCoV neg | 384 | 94.3 | 5 | 1.9 | 265 | 98.1 | Ref | ||
| FCoV pos | 23 | 5.7 | 6 | 4.4 | 131 | 95.6 | 2.4 | 0.73–7.6 | 0.193 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duijvestijn, M.B.H.M.; Schuurman, N.N.M.P.; Vernooij, J.C.M.; van Leeuwen, M.A.J.M.; Bosch, B.-J.; van den Brand, J.M.A.; Wagenaar, J.A.; van Kuppeveld, F.J.M.; Egberink, H.F.; Verhagen, J.H. Serological Survey of Retrovirus and Coronavirus Infections, including SARS-CoV-2, in Rural Stray Cats in The Netherlands, 2020–2022. Viruses 2023, 15, 1531. https://doi.org/10.3390/v15071531
Duijvestijn MBHM, Schuurman NNMP, Vernooij JCM, van Leeuwen MAJM, Bosch B-J, van den Brand JMA, Wagenaar JA, van Kuppeveld FJM, Egberink HF, Verhagen JH. Serological Survey of Retrovirus and Coronavirus Infections, including SARS-CoV-2, in Rural Stray Cats in The Netherlands, 2020–2022. Viruses. 2023; 15(7):1531. https://doi.org/10.3390/v15071531
Chicago/Turabian StyleDuijvestijn, Mirjam B. H. M., Nancy N. M. P. Schuurman, Johannes C. M. Vernooij, Michelle A. J. M. van Leeuwen, Berend-Jan Bosch, Judith M. A. van den Brand, Jaap A. Wagenaar, Frank J. M. van Kuppeveld, Herman F. Egberink, and Josanne H. Verhagen. 2023. "Serological Survey of Retrovirus and Coronavirus Infections, including SARS-CoV-2, in Rural Stray Cats in The Netherlands, 2020–2022" Viruses 15, no. 7: 1531. https://doi.org/10.3390/v15071531
APA StyleDuijvestijn, M. B. H. M., Schuurman, N. N. M. P., Vernooij, J. C. M., van Leeuwen, M. A. J. M., Bosch, B.-J., van den Brand, J. M. A., Wagenaar, J. A., van Kuppeveld, F. J. M., Egberink, H. F., & Verhagen, J. H. (2023). Serological Survey of Retrovirus and Coronavirus Infections, including SARS-CoV-2, in Rural Stray Cats in The Netherlands, 2020–2022. Viruses, 15(7), 1531. https://doi.org/10.3390/v15071531