
Development of Respiratory Syncytial Virus Vaccine Candidates for the Elderly

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells, Virus, Plasmids
2.2. Preparation, Characterization, and Validation of VLP Stocks
2.3. Preparation of Soluble F and G Proteins
2.4. Quantification of Soluble and VLP-Associated F and G Proteins
2.5. Preparation of RSV, RSV Plaque Assays, and Antibody Neutralization
2.6. Antibodies
2.7. Animals
2.8. Lung Histopathology
2.9. RSV Gene Expression by Real-Time PCR
2.10. ELISA
2.11. Experimental Design
2.12. Statistical Analysis
3. Results
3.1. Impact of Age of Immunization on Immune Responses
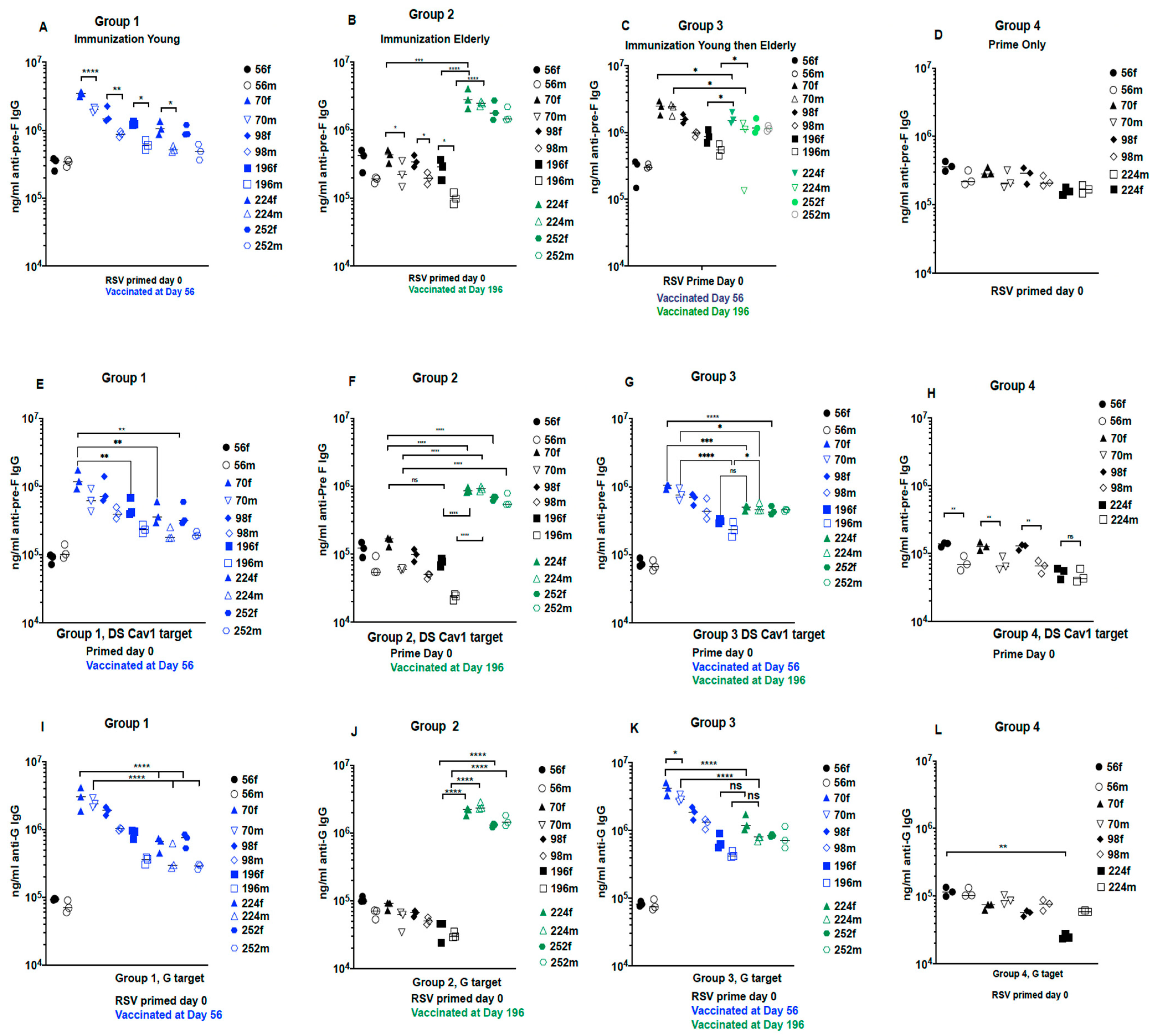
3.1.1. Total Anti-Pre-F and Anti-G Protein IgG Titers 8 Weeks after VLP Immunization
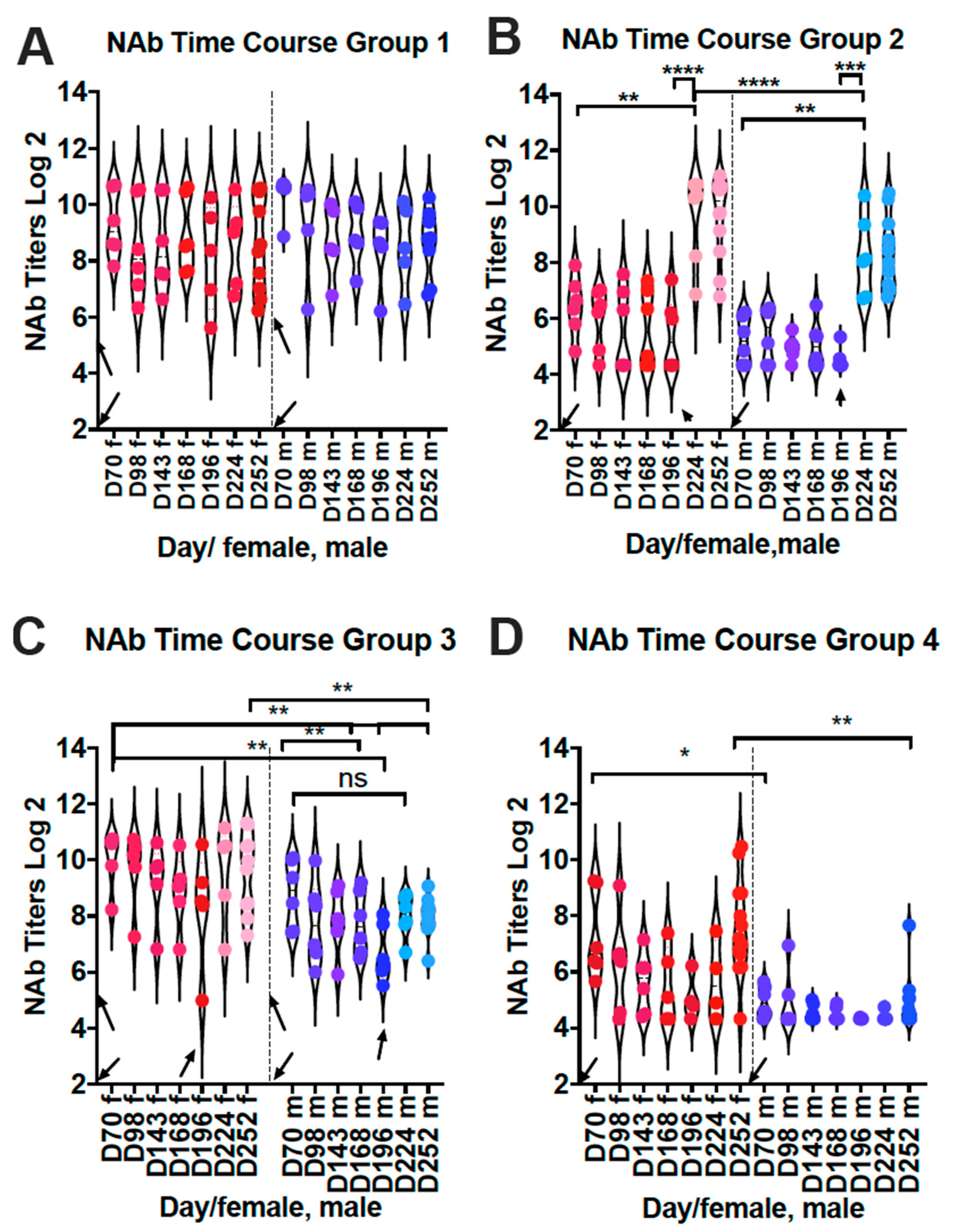
3.1.2. Total Neutralizing Antibody Titers 8 Weeks after VLP Immunization
3.2. Impact of One vs. Two Immunizations at Different Ages
3.2.1. Protocol 2
3.2.2. Total Anti-Pre-F and Anti-G Antibodies in Sera with Time after Immunization
3.2.3. Impact of Times of Immunization on Neutralization Titers in Sera
3.2.4. Impact of Age on Protection from RSV Replication by VLP Immunization
3.2.5. Assessment of Lung Pathology after RSV Challenge of CR Immunized Once or Twice with VLPs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, C.B.; Walsh, E.E.; Long, C.E.; Schnabel, K.C. Immunity to and Frequency of Reinfection with Respirtory Syncytial Virus. J. Infect. Dis. 1991, 163, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.; Shay, D.K.; Weintraub, E.; Brammer, L.; Cox, N.; Anderson, L.J.; Fukuda, K. Mortality associated with influenza and respiratory syncytial virus in the United States. JAMA 2003, 289, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Ann Falsey, R.; Hennessey, P.A.; Formica, M.A.; Cox, C.; Walsh, E.E. Respiratory Syncytial Virus Infection in Elderly and High-Risk Adults. N. Engl. J. Med. 2005, 352, 1749–1759. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; McAllister, D.A.; O’Brien, K.L.; Simoes, E.A.F.; Madhi, S.A.; Gessner, B.D.; Polack, F.P.; Balsells, E.; Acacio, S.; Aguayo, C.; et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: A systematic review and modelling study. Lancet 2017, 390, 946–958. [Google Scholar] [CrossRef] [PubMed]
- Han, L.L.; Alexander, J.P.; Anderson, L.J. Respiratory Syncytial Virus Pneumonia among the Elderly: An Assessment of Disease Burden. J. Infect. Dis. 1999, 179, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Falsey, A.R.; Walsh, E.E. Respiratory Syncytial Virus Infection in Adults. Clin. Microbiol. Rev. 2000, 13, 371–384. [Google Scholar] [CrossRef]
- Schneider, J.L.; Rowe, J.H.; Garcia-de-Alba, C.; Kim, C.F.; Sharpe, A.H.; Haigis, M.C. The aging lung: Physiology, disease, and immunity. Cell 2021, 184, 1990–2019. [Google Scholar] [CrossRef]
- Chen, W.H.; Kozlovsky, B.F.; Effros, R.B.; Grubeck-Loebenstein, B.; Edelman, R.; Sztein, M.B. Vaccination in the elderly: An immunological perspective. Trends Immunol. 2009, 30, 351–359. [Google Scholar] [CrossRef]
- Frasca, D.; Blomberg, B.B. B cell function and influenza vaccine responses in healthy aging and disease. Curr. Opin. Immunol. 2014, 29, 112–118. [Google Scholar] [CrossRef]
- Janse, A.G.S.C.; Sanders, E.A.M.; Hoes, A.W.; van Loon, A.M.; Hak, E. Influenza- and Respiratory Syncytial Virus-Associated Mortality and Hospitalisations. Eur. Respir. J. 2007, 30, 1158–1166. [Google Scholar] [CrossRef]
- Dzau, V.J.; Inouye, S.K.; Rowe, J.W.; Finkelman, E.; Yamada, T. Enabling Healthful Aging for All—The National Academy of Medicine Grand Challenge in Healthy Longevity. N. Engl. J. Med. 2019, 381, 1699–1701. [Google Scholar] [CrossRef]
- Guichelaar, T.; Hoeboer, J.; Widjojoatmodjo, M.N.; Reemers, S.S.N.; van Els, C.A.C.M.; Otten, R.; van Remmerden, Y.; Boes, J.; Luytjes, W. Impaired Immune Response to Vaccination against Infection with Human Respiratory Syncytial Virus at Advanced Age. J. Virol. 2014, 88, 9744–9750. [Google Scholar] [CrossRef]
- Henry, C.; Zheng, N.-Y.; Huang, M.; Cabanov, A.; Rojas, K.T.; Kaur, K.; Andrews, S.F.; Palm, A.-K.E.; Chen, Y.-Q.; Li, Y.; et al. Influenza Virus Vaccination Elicits Poorly Adapted B Cell Responses in Elderly Individuals. Cell Host Microbe 2019, 25, 357–366.e6. [Google Scholar] [CrossRef]
- Pangesti, K.N.A.; Abd El Ghany, M.; Walsh, M.G.; Kesson, A.M.; Hill-Cawthorne, G.A. Molecular epidemiology of respiratory syncytial virus. Rev. Med. Virol. 2018, 28, e1968. [Google Scholar] [CrossRef]
- Cullen, L.M.; Schmidt, M.R.; Morrison, T.G. Effect of Previous Respiratory Syncytial Virus Infection on Murine Immune Responses to F and G Protein-Containing Virus-Like Particles. J. Virol. 2019, 93, e00087-19. [Google Scholar] [CrossRef]
- Blanco, J.C.G.; Pletneva, L.M.; Cullen, L.; Otoa, R.O.; Patel, M.C.; Fernando, L.R.; Boukhvalova, M.S.; Morrison, T.G. Efficacy of a respiratory syncytial virus vaccine candidate in a maternal immunization model. Nat. Comm. 2018, 9, 1904–1914. [Google Scholar] [CrossRef]
- Blanco, J.; Pletneva, L.; Oue, R.; Patel, M.; Boukhvalova, M.S. Maternal transfer of RSV immunity in cotton rats vaccinated during pregnancy. Vaccine 2015, 33, 5371–5379. [Google Scholar] [CrossRef]
- Murawski, M.R.; McGinnes, L.W.; Finberg, R.W.; Kurt-Jones, E.A.; Massare, M.; Smith, G.; Morrison, T.G. Newcastle disease virus-like particles containing respiratory syncytial virus G protein induced protection in BALB/c mice with no evidence of immunopathology. J. Virol. 2010, 84, 1110–1123. [Google Scholar] [CrossRef]
- McGinnes, L.W.; Gravel, K.A.; Finberg, R.W.; Kurt-Jones, E.A.; Massare, M.J.; Smith, G. Assembly and immunological properties of Newcastle disease virus-like particles containing the respiratory syncytial virus F and G proteins. J. Virol. 2011, 85, 366–377. [Google Scholar] [CrossRef]
- McGinnes, L.; Schmidt, M.R.; Kenward, S.A.; Woodland, R.T.; Morrison, T.G. Murine Immune Responses to Virus-Like Particle-Associated Pre- and Postfusion Forms of the Respiratory Syncytial Virus F Protein. J. Virol. 2015, 89, 6835–6847. [Google Scholar] [CrossRef]
- Cullen, M.L.; Schmidt, R.M.; Torres, M.G.; Capoferri, A.A.; Morrison, G.T. Comparison of Immune Responses to Different Versions of VLP Associated Stabilized RSV Pre-Fusion F Protein. Vaccines 2019, 7, 21–41. [Google Scholar] [CrossRef] [PubMed]
- Krarup, A.; Truan, D.; Furmanova-Hollenstein, P.; Bogaert, L.; Bouchier, P.; Bisschop, I.J.M.; Widjojoatmodjo, M.N.; Zahn, R.; Schuitemaker, H.; McLellan, J.S.; et al. A highly stable prefusion RSV F vaccine derived from structural analysis of the fusion mechanism. Nat. Commun. 2015, 6, 8143–8155. [Google Scholar] [CrossRef] [PubMed]
- McLellan, J.S.; Chen, M.; Joyce, M.G.; Sastry, M.; Stewart-Jones, G.B.E.; Yang, Y. Structure-based design of a fusion glycoprotein vaccine for respiratory syncytial virus. Science 2013, 342, 592–598. [Google Scholar] [CrossRef] [PubMed]
- McGinnes, L.W.; Morrison, T.G. Newcastle Disease Virus-Like Particles: Preparation, Purification, Quantification, and Incorporation of Foreign Glycoproteins. In Current Protocols in Microbiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Cullen, L.M.; Boukhvalova, M.S.; Blanco, J.C.G.; Morrison, T.G. Comparisons of Antibody Populations in Different Pre-Fusion F VLP Immunized Cotton Rat Dams and Their Offspring. Vaccines 2020, 8, 133. [Google Scholar] [CrossRef]
- Cullen, L.M.; Schmidt, M.R.; Morrison, T.G. The importance of RSV F protein conformation in VLPs in stimulation of neutralizing antibody titers in mice previously infected with RSV. Hum. Vaccines Immunother. 2017, 13, 2814–2823. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Green, G.; Johnson, S.M.; Costello, H.; Brakel, K.; Harder, O.; Oomens, A.G.; Peeples, M.E.; Moulton, H.M.; Niewiesk, S.; Heise, M.T. CX3CR1 Is a Receptor for Human Respiratory Syncytial Virus in Cotton Rats. J. Virol. 2021, 95, e00010-21. [Google Scholar] [CrossRef]
- Boukhvalova, M.S.; Blanco, J. The cotton rat Sigmondon Hispidus model of respiratory syncytial virus infection. In Challenges and Opportunities for Respiratory Syncytial Virus Vaccines; Anderson, L.J., Graham, B.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Blanco, J.; Cullen, L.; Kamali, A.; Sylla, F.; Boukhvalova, M.S.; Morrison, T.G. Evolution of protection after maternal immunization for respiratory syncytial virus in cotton rats. PLoS Pathog. 2021, 17, e1009856. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Boukhvalova, M.S.; Prince, G.A.; Blanco, J.C.G. Respiratory Syncytial Virus Infects and Abortively Replicates in the Lungs in Spite of Preexisting Immunity. J. Virol. 2007, 81, 9443–9450. [Google Scholar] [CrossRef]
- Boukhvalova, M.S.; Kevin, C.Y.; Katie, H.K.; John, P.H.; Gregory, A. Prince, David D. Porter, and Jorge C. G. Blanco. Age-Related Differences in Pulmonary Cytokine Response to Respiratory Syncytial Virus Infection: Modulation by Anti-Inflammatory and Antiviral Treatment. J. Infect. Dis. 2007, 195, 511–518. [Google Scholar] [CrossRef]
- Lee, J.; Paparoditis, P.; Horton, A.P.; Frühwirth, A.; McDaniel, J.R.; Jung, J.; Boutz, D.R.; Hussein, D.A.; Tanno, Y.; Pappas, L.; et al. Persistent Antibody Clonotypes Dominate the Serum Response to Influenza over Multiple Years and Repeated Vaccinations. Cell Host Microbe 2019, 25, 367–376.e5. [Google Scholar] [CrossRef]
- Devarajan, P.; Swain, S.L. Original Antigenic Sin: Friend or Foe in Developing a Broadly Cross-Reactive Vaccine to Influenza? Cell Host Microbe 2019, 25, 354–355. [Google Scholar] [CrossRef]
- Hall, C.B.; Simoes, E.A.F.; Anderson, L.J. Clinical and Epidemiologic Features of Respiratory Syncytial Virus. In Challenges and Opportunities for Respiratory Syncytial Virus Vaccines; Anderson, L.J., Graham, B.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 39–58. [Google Scholar]
- Glezen, W.P.; Taber, L.H.; Frank, A.L.; Kasel, J.A. Risk of Primary Infection and Reinfection with Respiratory Syncytial Virus. Am. J. Dis. Child. 1986, 140, 543–546. [Google Scholar] [CrossRef]
- Boelen, A.; Andeweg, A.; Kwakkel, J.; Lokhorst, W.; Bestebroer, T.; Dormans, J.; Kimman, T. Both immunisation with a formalin-inactivated respiratory syncytial virus (RSV) vaccine and a mock antigen vaccine induce severe lung pathology and a Th2 cytokine profile in RSV-challenged mice. Vaccine 2000, 19, 982–991. [Google Scholar] [CrossRef]
- Acosta, P.L.; Caballero, M.; Polack, F. Brief History and Characterization of Enhanced Respiratory Syncytial Virus Disease. Clin. Vaccine Immunol. 2016, 23, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Ohrum, K.; Snell Bennett, A.; Rajani Gaurav, M.; Hostetler, L.; Maynard Sean, K.; Lazzaro, M.; Cheng Lily, I.; O’Day, T.; Cayatte, C. CD4+ T Cells Drive Lung Disease Enhancement Induced by Immunization with Suboptimal Doses of Respiratory Syncytial Virus Fusion Protein in the Mouse Model. J. Virol. 2019, 93, e00695-19. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Ohrum, K.; Cayatte, C.; Bennett, A.S.; Rajani, G.M.; McTamney, P.; Nacel, K.; Hostetler, L.; Cheng, L.; Ren, K.; O’Day, T.; et al. Immunization with Low Doses of Recombinant Postfusion or Prefusion Respiratory Syncytial Virus F Primes for Vaccine-Enhanced Disease in the Cotton Rat Model Independently of the Presence of a Th1-Biasing (GLA-SE) or Th2-Biasing (Alum) Adjuvant. J. Virol. 2017, 91, e02180-16. [Google Scholar] [CrossRef] [PubMed]
- Prince, G.A.; Prieels, J.P.; Sllaoui, M.; Porter, D.D. Pulmmonary lesions in primary respiratory syncytial virus infection, reinfection, and vaccine-enhanced disease in cotton rats (Sigmodon hispidus). Lab. Investig. 1999, 79, 1385–1392. [Google Scholar] [PubMed]
- Blanco, J.C.G.; Pletneva, L.M.; Otoa, R.O.; Patel, M.C.; Vogel, S.N.; Boukhvalova, M.S. Preclinical assessment of safety of maternal vaccination against respiratory syncytial virus (RSV) in cotton rats. Vaccine 2017, 35, 3951–3958. [Google Scholar] [CrossRef]
- Anderson, L.J. Respiratory syncytial virus vaccine development. Semin. Immunol. 2013, 25, 160–171. [Google Scholar] [CrossRef]
- Anderson, L.J.; Dormitzer, P.R.; Nokes, D.J.; Rappuoli, R.; Roca, A.; Graham, B.S. Strategic priorities for respiratory syncytial virus (RSV) vaccine development. Vaccine 2013, 31 (Suppl. S2), B209–B215. [Google Scholar] [CrossRef]
- Del Giudice, G.; Weinberger, B.; Grubeck-Loebenstein, B. Vaccines for the Elderly. Gerontology 2015, 61, 203–210. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco, J.C.G.; Cullen, L.M.; Kamali, A.; Sylla, F.Y.D.; Boukhvalova, M.S.; Morrison, T.G. Development of Respiratory Syncytial Virus Vaccine Candidates for the Elderly. Viruses 2023, 15, 1305. https://doi.org/10.3390/v15061305
Blanco JCG, Cullen LM, Kamali A, Sylla FYD, Boukhvalova MS, Morrison TG. Development of Respiratory Syncytial Virus Vaccine Candidates for the Elderly. Viruses. 2023; 15(6):1305. https://doi.org/10.3390/v15061305
Chicago/Turabian StyleBlanco, Jorge C. G., Lori M. Cullen, Arash Kamali, Fatouomata Y. D. Sylla, Marina S. Boukhvalova, and Trudy G. Morrison. 2023. "Development of Respiratory Syncytial Virus Vaccine Candidates for the Elderly" Viruses 15, no. 6: 1305. https://doi.org/10.3390/v15061305
APA StyleBlanco, J. C. G., Cullen, L. M., Kamali, A., Sylla, F. Y. D., Boukhvalova, M. S., & Morrison, T. G. (2023). Development of Respiratory Syncytial Virus Vaccine Candidates for the Elderly. Viruses, 15(6), 1305. https://doi.org/10.3390/v15061305