
Remote Sensing and Ecological Variables Related to Influenza A Prevalence and Subtype Diversity in Wild Birds in the Lluta Wetland of Northern Chile
 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Sample Size
- n = required sampling size;
- N = population size;
- = 1-confidence level;
- D = estimated minimum number of positive samples.
2.3. Influenza A virus Detection
2.4. Sequencing and Phylogenic Analysis
2.5. Ecological and Environmental Variables
2.6. Statistical Analysis
3. Results
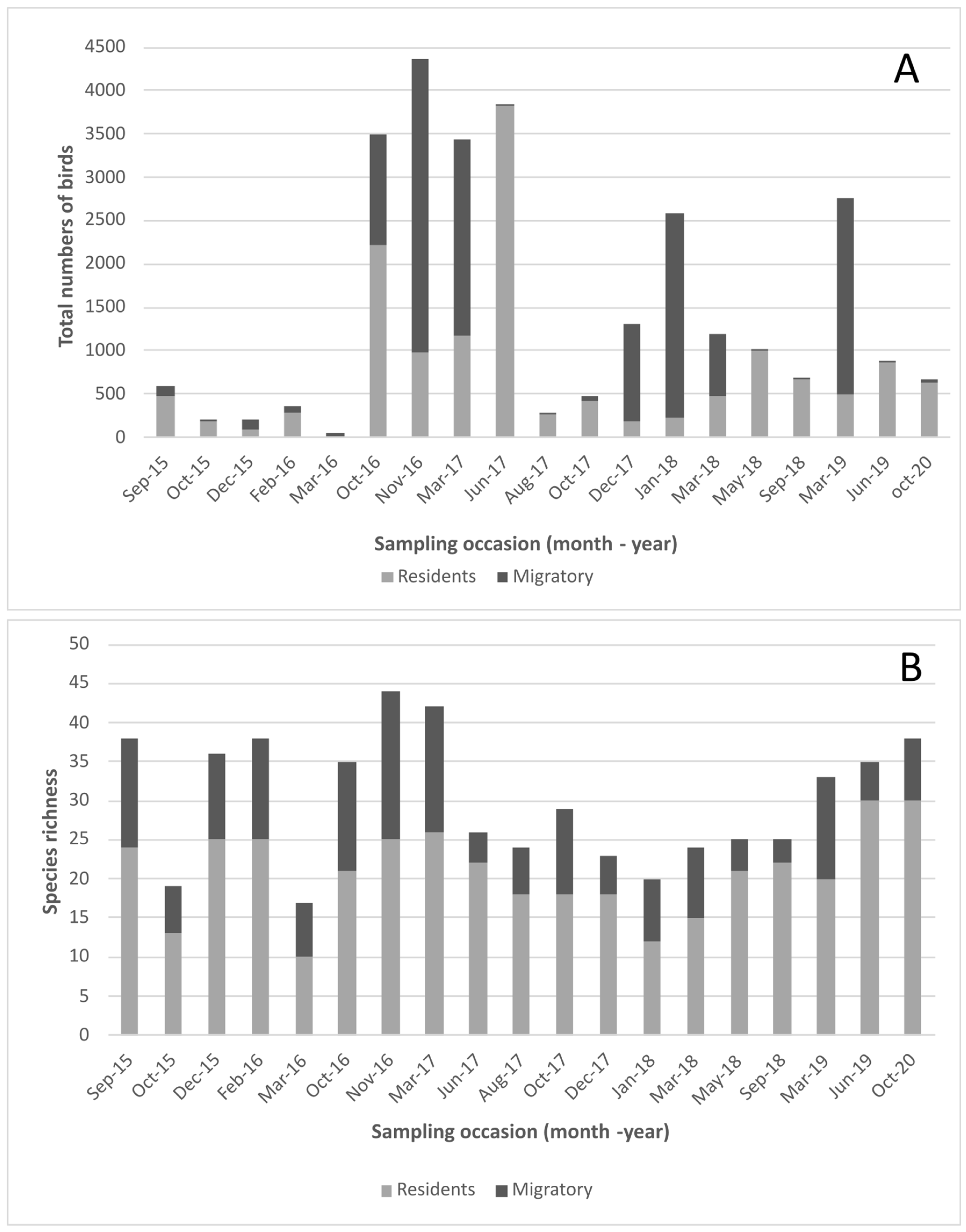
3.1. Influenza Prevalence and Species Richness
3.2. Environmental Variables
3.3. Subtype Diversity and Sequence Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olsen, B.; Munster, V.J.; Wallensten, A.; Waldenström, J.; Osterhaus, A.D.; Fouchier, R.A. Global patterns of influenza A virus in wild birds. Science 2006, 312, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.; Hulse, D. Microbial adaptation and change: Avian influenza. Rev. Sci. Technol.-Off. Int. Epizoot. 2004, 23, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Caliendo, V.; Lewis, N.; Pohlmann, A.; Baillie, S.; Banyard, A.; Beer, M.; Brown, I.; Fouchier, R.; Hansen, R.; Lameris, T. Transatlantic spread of highly pathogenic avian influenza H5N1 by wild birds from Europe to North America in 2021. Sci. Rep. 2022, 12, 11729. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, B.; Raghwani, J.; Vrancken, B.; Jia, R.; Hill, S.C.; Fournié, G.; Cheng, Y.; Yang, Q.; Wang, Y. Bidirectional movement of emerging H5N8 avian influenza viruses between Europe and Asia via migratory birds since early 2020. Mol. Biol. Evol. 2023, 40, msad019. [Google Scholar] [CrossRef]
- Gorsich, E.E.; Webb, C.T.; Merton, A.A.; Hoeting, J.A.; Miller, R.S.; Farnsworth, M.L.; Swafford, S.R.; DeLiberto, T.J.; Pedersen, K.; Franklin, A.B. Continental-scale dynamics of avian influenza in US waterfowl are driven by demography, migration, and temperature. Ecol. Appl. 2021, 31, e2245. [Google Scholar] [CrossRef] [PubMed]
- Hoye, B.J.; Munster, V.J.; Nishiura, H.; Klaassen, M.; Fouchier, R.A. Surveillance of wild birds for avian influenza virus. Emerg. Infect. Dis. 2010, 16, 1827. [Google Scholar] [CrossRef]
- Verhagen, J.H.; Fouchier, R.A.; Lewis, N. Highly pathogenic avian influenza viruses at the wild–domestic bird interface in Europe: Future directions for research and surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef]
- Jiménez-Bluhm, P.; Karlsson, E.A.; Freiden, P.; Sharp, B.; Di Pillo, F.; Osorio, J.E.; Hamilton-West, C.; Schultz-Cherry, S. Wild birds in Chile Harbor diverse avian influenza A viruses. Emerg. Microbes Infect. 2018, 7, 1–4. [Google Scholar] [CrossRef]
- Nelson, M.I.; Pollett, S.; Ghersi, B.; Silva, M.; Simons, M.P.; Icochea, E.; Gonzalez, A.E.; Segovia, K.; Kasper, M.R.; Montgomery, J.M. The genetic diversity of influenza A viruses in wild birds in Peru. PLoS ONE 2016, 11, e0146059. [Google Scholar]
- Rimondi, A.; Gonzalez-Reiche, A.S.; Olivera, V.S.; Decarre, J.; Castresana, G.J.; Romano, M.; Nelson, M.I.; van Bakel, H.; Pereda, A.J.; Ferreri, L. Evidence of a fixed internal gene constellation in influenza A viruses isolated from wild birds in Argentina (2006–2016). Emerg. Microbes Infect. 2018, 7, 1–13. [Google Scholar] [CrossRef]
- Ruiz, S.; Jimenez-Bluhm, P.; Di Pillo, F.; Baumberger, C.; Galdames, P.; Marambio, V.; Salazar, C.; Mattar, C.; Sanhueza, J.; Schultz-Cherry, S. Temporal dynamics and the influence of environmental variables on the prevalence of avian influenza virus in main wetlands in central Chile. Transbound. Emerg. Dis. 2021, 68, 1601–1614. [Google Scholar] [CrossRef]
- Xu, K.; Ferreri, L.; Rimondi, A.; Olivera, V.; Romano, M.; Ferreyra, H.; Rago, V.; Uhart, M.; Chen, H.; Sutton, T. Isolation and characterization of an H9N2 influenza virus isolated in Argentina. Virus Res. 2012, 168, 41–47. [Google Scholar] [CrossRef] [PubMed]
- García-Walther, J.; Senner, N.; Norambuena, H.; Schmitt, F. Atlas De Las Aves Playeras de Chile: Sitios Importantes Para Su Conservación; Universidad Santo Tomás: Santiago, Chile, 2017. [Google Scholar]
- Bravo-Vasquez, N.; Di Pillo, F.; Lazo, A.; Jiménez-Bluhm, P.; Schultz-Cherry, S.; Hamilton-West, C. Presence of influenza viruses in backyard poultry and swine in El Yali wetland, Chile. Prev. Vet. Med. 2016, 134, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Bluhm, P.; Bravo-Vasquez, N.; Torchetti, M.K.; Killian, M.L.; Livingston, B.; Herrera, J.; Fuentes, M.; Schultz-Cherry, S.; Hamilton-West, C. Low pathogenic avian influenza (H7N6) virus causing an outbreak in commercial Turkey farms in Chile. Emerg. Microbes Infect. 2019, 8, 479–485. [Google Scholar] [CrossRef]
- Jimenez-Bluhm, P.; Di Pillo, F.; Bahl, J.; Osorio, J.; Schultz-Cherry, S.; Hamilton-West, C. Circulation of influenza in backyard productive systems in central Chile and evidence of spillover from wild birds. Prev. Vet. Med. 2018, 153, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Pedreros, A.; Merino, C. Diversity of aquatic bird species in a wetland complex in southern Chile. J. Nat. Hist. 2014, 48, 1453–1465. [Google Scholar] [CrossRef]
- Sielfeld, W.; Peredo, R.; Fuentes, R.; Malinarich, V.; Olivares, F. Coastal Wetlands of Northern Chile. In The Ecology and Natural History of Chilean Saltmarshes; Springer: Berlin/Heidelberg, Germany, 2017; pp. 105–167. [Google Scholar]
- Navarro, N.; Abad, M.; Bonnail, E.; Izquierdo, T. The Arid Coastal Wetlands of Northern Chile: Towards an Integrated Management of Highly Threatened Systems. J. Mar. Sci. Eng. 2021, 9, 948. [Google Scholar] [CrossRef]
- Peredo, R. La Desembocadura Del Río Lluta: Un Humedal Para Las Aves, En El Desierto Costero De Chile. 2007. Available online: https://biblioteca.cehum.org/ (accessed on 9 December 2022).
- Gaidet, N.; Caron, A.; Cappelle, J.; Cumming, G.S.; Balança, G.; Hammoumi, S.; Cattoli, G.; Abolnik, C.; Servan de Almeida, R.; Gil, P. Understanding the ecological drivers of avian influenza virus infection in wildfowl: A continental-scale study across Africa. Proc. R. Soc. B Biol. Sci. 2012, 279, 1131–1141. [Google Scholar] [CrossRef]
- Cumming, G.S.; Abolnik, C.; Caron, A.; Gaidet, N.; Grewar, J.; Hellard, E.; Henry, D.A.; Reynolds, C. A social–ecological approach to landscape epidemiology: Geographic variation and avian influenza. Landsc. Ecol. 2015, 30, 963–985. [Google Scholar] [CrossRef]
- Ferenczi, M.; Beckmann, C.; Warner, S.; Loyn, R.; O’riley, K.; Wang, X.; Klaassen, M. Avian influenza infection dynamics under variable climatic conditions, viral prevalence is rainfall driven in waterfowl from temperate, south-east Australia. Vet. Res. 2016, 47, 23. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ramírez, E.; Acevedo, P.; Allepuz, A.; Gerrikagoitia, X.; Alba, A.; Busquets, N.; Díaz-Sánchez, S.; Álvarez, V.; Abad, F.X.; Barral, M. Ecological factors driving avian influenza virus dynamics in Spanish wetland ecosystems. PLoS ONE 2012, 7, e46418. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Wang, T.; Skidmore, A.K.; de Boer, W.F.; Li, L.; Prins, H.H. Environmental factors influencing the spread of the highly pathogenic avian influenza H5N1 virus in wild birds in Europe. Ecol. Soc. 2010, 15, 26. [Google Scholar] [CrossRef]
- Torrontegi, O.; Alvarez, V.; Acevedo, P.; Gerrikagoitia, X.; Höfle, U.; Barral, M. Long-term avian influenza virus epidemiology in a small Spanish wetland ecosystem is driven by the breeding Anseriformes community. Vet. Res. 2019, 50, 4. [Google Scholar] [CrossRef] [PubMed]
- Dohoo, I.R.; Martin, W.; Stryhn, H.E. Veterinary Epidemiologic Research; University of Prince Edward Island: Charlottetown, PE, Canada, 2003. [Google Scholar]
- WHO. CDC protocol of realtime RTPCR for swine influenza A (H1N1). In CDC Protocol of Realtime RTPCR for Swine Influenza A (H1N1); World Health Organization (WHO): Geneva, Switzerland, 2009. [Google Scholar]
- Shu, B.; Wu, K.-H.; Emery, S.; Villanueva, J.; Johnson, R.; Guthrie, E.; Berman, L.; Warnes, C.; Barnes, N.; Klimov, A. Design and performance of the CDC real-time reverse transcriptase PCR swine flu panel for detection of 2009 A (H1N1) pandemic influenza virus. J. Clin. Microbiol. 2011, 49, 2614–2619. [Google Scholar] [CrossRef] [PubMed]
- Moresco, K.A.; Stallknecht, D.E.; Swayne, D.E. Evaluation and attempted optimization of avian embryos and cell culture methods for efficient isolation and propagation of low pathogenicity avian influenza viruses. Avian Dis. 2010, 54, 622–626. [Google Scholar] [CrossRef]
- Cheung, P.P.; Leung, Y.C.; Chow, C.-K.; Ng, C.-F.; Tsang, C.-L.; Wu, Y.-O.; Ma, S.-K.; Sia, S.-F.; Guan, Y.; Peiris, J.M. Identifying the species-origin of faecal droppings used for avian influenza virus surveillance in wild-birds. J. Clin. Virol. 2009, 46, 90–93. [Google Scholar] [CrossRef]
- Kaplan, B.S.; Russier, M.; Jeevan, T.; Marathe, B.; Govorkova, E.A.; Russell, C.J.; Kim-Torchetti, M.; Choi, Y.K.; Brown, I.; Saito, T. Novel highly pathogenic avian A (H5N2) and A (H5N8) influenza viruses of clade 2.3. 4.4 from North America have limited capacity for replication and transmission in mammals. Msphere 2016, 1, e00003-16. [Google Scholar] [CrossRef]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes de novo assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Bao, Y.; Bolotov, P.; Dernovoy, D.; Kiryutin, B.; Zaslavsky, L.; Tatusova, T.; Ostell, J.; Lipman, D. The influenza virus resource at the National Center for Biotechnology Information. J. Virol. 2008, 82, 596–601. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Oxford University Press: Oxford, UK; pp. 95–98.
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Rouse, J.W., Jr.; Haas, R.H.; Schell, J.; Deering, D. Monitoring the Vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation; National Space Agency: Greenbelt, MD, USA, 1973. [Google Scholar]
- Xu, H. Modification of normalised difference water index (NDWI) to enhance open water features in remotely sensed imagery. Int. J. Remote Sens. 2006, 27, 3025–3033. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing. 2018. Available online: http://www.r-project.org/index.html (accessed on 7 December 2022).
- Empres-i, F. Diseases. 2022. Available online: https://empres-i.apps.fao.org/diseases (accessed on 25 November 2022).
- Wahis. Sistema Mundial de Información Zoosanitaria. 2022. Available online: https://wahis.woah.org/#/home (accessed on 25 November 2022).
- Jones, Y.L.; Swayne, D.E. Comparative pathobiology of low and high pathogenicity H7N3 Chilean avian influenza viruses in chickens. Avian Dis. 2004, 48, 119–128. [Google Scholar] [CrossRef]
- Youk, S.; Leyson, C.; Killian, M.L.; Torchetti, M.K.; Lee, D.-h.; Suarez, D.L.; Pantin-Jackwood, M.J. Evolution of the North American Lineage H7 Avian Influenza Viruses in Association with H7 Virus’s Introduction to Poultry. J. Virol. 2022, 96, e00278-22. [Google Scholar] [CrossRef]
- Dietze, K.; Graaf, A.; Homeier-Bachmann, T.; Grund, C.; Forth, L.; Pohlmann, A.; Jeske, C.; Wintermann, M.; Beer, M.; Conraths, F.J. From low to high pathogenicity—Characterization of H7N7 avian influenza viruses in two epidemiologically linked outbreaks. Transbound. Emerg. Dis. 2018, 65, 1576–1587. [Google Scholar] [CrossRef]
- Caron, A.; Abolnik, C.; Mundava, J.; Gaidet, N.; Burger, C.E.; Mochotlhoane, B.; Bruinzeel, L.; Chiweshe, N.; de Garine-Wichatitsky, M.; Cumming, G.S. Persistence of low pathogenic avian influenza virus in waterfowl in a Southern African ecosystem. EcoHealth 2011, 8, 109–115. [Google Scholar] [CrossRef]
- Van Dijk, J.G.; Hoye, B.J.; Verhagen, J.H.; Nolet, B.A.; Fouchier, R.A.; Klaassen, M. Juveniles and migrants as drivers for seasonal epizootics of avian influenza virus. J. Anim. Ecol. 2014, 83, 266–275. [Google Scholar] [CrossRef]
- Kouba, M.; Bartoš, L.; Tomášek, V.; Popelková, A.; Šťastný, K.; Zárybnická, M. Home range size of Tengmalm’s owl during breeding in Central Europe is determined by prey abundance. PLoS ONE 2017, 12, e0177314. [Google Scholar] [CrossRef]
- Tucker, M.A.; Alexandrou, O.; Bierregaard, R.O., Jr.; Bildstein, K.L.; Böhning-Gaese, K.; Bracis, C.; Brzorad, J.N.; Buechley, E.R.; Cabot, D.; Calabrese, J.M. Large birds travel farther in homogeneous environments. Glob. Ecol. Biogeogr. 2019, 28, 576–587. [Google Scholar] [CrossRef]
- Canavan, B.C. Opening Pandora’s Box at the roof of the world: Landscape, climate and avian influenza (H5N1). Acta Trop. 2019, 196, 93–101. [Google Scholar] [CrossRef]
- Cai, W.; McPhaden, M.J.; Grimm, A.M.; Rodrigues, R.R.; Taschetto, A.S.; Garreaud, R.D.; Dewitte, B.; Poveda, G.; Ham, Y.-G.; Santoso, A. Climate impacts of the El Niño–southern oscillation on South America. Nat. Rev. Earth Environ. 2020, 1, 215–231. [Google Scholar] [CrossRef]
- Di Pillo, F.; Baumberger, C.; Salazar, C.; Galdames, P.; Ruiz, S.; Sharp, B.; Freiden, P.; Tan, S.; Schultz-Cherry, S.; Hamilton-West, C. Novel Low Pathogenic Avian Influenza H6N1 in Backyard Chicken in Easter Island (Rapa Nui), Chilean Polynesia. Viruses 2022, 14, 718. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, C.; Moreno, V.; Pedersen, J.; Jeria, J.; Agredo, M.; Gutiérrez, C.; García, A.; Vásquez, M.; Avalos, P.; Retamal, P. Avian influenza in wild birds from Chile, 2007–2009. Virus Res. 2015, 199, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Ghersi, B.M.; Blazes, D.L.; Icochea, E.; Gonzalez, R.I.; Kochel, T.; Tinoco, Y.; Sovero, M.M.; Lindstrom, S.; Shu, B.; Klimov, A. Avian influenza in wild birds, central coast of Peru. Emerg. Infect. Dis. 2009, 15, 935. [Google Scholar] [CrossRef]
- Farnsworth, M.L.; Miller, R.S.; Pedersen, K.; Lutman, M.W.; Swafford, S.R.; Riggs, P.D.; Webb, C.T. Environmental and demographic determinants of avian influenza viruses in waterfowl across the contiguous United States. PLoS ONE 2012, 7, e32729. [Google Scholar]
- Fuller, T.; Ducatez, M.; Njabo, K.; Couacy-Hymann, E.; Chasar, T.; Aplogan, G.; Lao, S.; Awoume, F.; Tehou, A.; Langeois, Q. Seasonality dynamics of avian influenza occurrences in Central and West Africa. bioRxiv 2014. [Google Scholar] [CrossRef]
- Ashok, A.; Rani, H.P.; Jayakumar, K. Monitoring of dynamic wetland changes using NDVI and NDWI based landsat imagery. Remote Sens. Appl. Soc. Environ. 2021, 23, 100547. [Google Scholar] [CrossRef]
- Wu, X.; Lv, M.; Jin, Z.; Michishita, R.; Chen, J.; Tian, H.; Tu, X.; Zhao, H.; Niu, Z.; Chen, X. Normalized difference vegetation index dynamic and spatiotemporal distribution of migratory birds in the Poyang Lake wetland, China. Ecol. Indic. 2014, 47, 219–230. [Google Scholar] [CrossRef]
- Gaidet, N. Ecology of avian influenza virus in wild birds in tropical Africa. Avian Dis. 2016, 60, 296–301. [Google Scholar] [CrossRef]
- Mathieu, C.; Moreno, V.; Retamal, P.; Gonzalez, A.; Rivera, A.; Fuller, J.; Jara, C.; Lecocq, C.; Rojas, M.; García, A. Pandemic (H1N1) 2009 in breeding turkeys, Valparaiso, Chile. Emerg. Infect. Dis. 2010, 16, 709. [Google Scholar] [CrossRef] [PubMed]
- Pereda, A.J.; Uhart, M.; Perez, A.A.; Zaccagnini, M.E.; La Sala, L.; Decarre, J.; Goijman, A.; Solari, L.; Suarez, R.; Craig, M.I. Avian influenza virus isolated in wild waterfowl in Argentina: Evidence of a potentially unique phylogenetic lineage in South America. Virology 2008, 378, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, B.L.; Wood, C.L.; Iliff, M.J.; Bonney, R.E.; Fink, D.; Kelling, S. eBird: A citizen-based bird observation network in the biological sciences. Biol. Conserv. 2009, 142, 2282–2292. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Definition | |
|---|---|---|
| Wild bird community | Total abundance | Total number of birds present at the time of sampling |
| Species richness | Number of species present at the time of sampling | |
| Abundance of migrants | Number of migratory birds present at the time of sampling | |
| Landscape | Vegetation coverage | Mean NDVI for the month of sampling, one month before, two months before, and three months before sampling. |
| Water body size (km2) | Water body size for the month of sampling, one month before, two months before, and three months before sampling. | |
| Meteorological data | Maximum monthly temperature (°C) | Monthly mean of maximum daily temperature for the month of sampling, one month before, two months before, and three months before sampling. |
| Minimum monthly temperature (°C) | Monthly mean of minimum daily temperature for the month of sampling, one month before, two months before, and three months before sampling. | |
| Total monthly rainfall (mm) | Total rainfall at the month of sampling, one month before, two months before, and three months before sampling. | |
| Humidity (%) | Relative air humidity at the month of sampling, one month before, two months before, and three months before sampling. | |
| Variables | Categories | Estimate | p-Value | OR | 95% CI |
|---|---|---|---|---|---|
| (Intercept) | −5.5872 | <0.001 | 0.003 | (0.001–0.01) | |
| NDVI | Low (<0.27) | Reference | |||
| High (≥0.27) | 1.2949 | 0.00230 | 3.65 | (1.58–8.39) | |
| Abundance of Migrants | Low (<113) | Reference | |||
| High (≥113) | 1.2727 | 0.00733 | 3.57 | (1.41–9.05) |
| Strain Name | Year | Isolate Subtype | Host Species | Order | Accession Number |
|---|---|---|---|---|---|
| A/American oystercatcher/Chile/C1307/2015 | 2015 | H9N2 | Haematopus palliatus | Charadriiformes | KX185901 |
| A/Grey plover/Chile/C1313/2015 | 2015 | H9N7 | Pluvialis squatarola | Charadriiformes | KX185918 |
| A/American oystercatcher/Chile/C17359/2016 | 2016 | H3N8 | Haematopus palliatus | Charadriiformes | MH499035 |
| A/American oystercatcher/Chile/C17400/2016 | 2016 | H3N8 | Haematopus palliatus | Charadriiformes | MH498968 |
| A/Franklin’s gull/Chile/C17421/2016 | 2016 | H13N9 | Larus pipixcan | Charadriiformes | MH498978 |
| A/Franklin’s gull/Chile/C17422/2016 | 2016 | H13N9 | Larus pipixcan | Charadriiformes | MH499057 |
| A/Kelp gull/Chile/C27733/2017 | 2017 | H13N8 | Larus dominicanus | Charadriiformes | MH499142 |
| A/Kelp gull/Chile/C35523/2017 | 2017 | H5N3 | Larus dominicanus | Charadriiformes | MK164009 |
| A/blackish oystercatcher/Chile/C40194/2018 | 2018 | H13N2 | Haematopus ater | Charadriiformes | OP888556 |
| A/Chilean flamingo/Chile/C52796/2020 | 2020 | H7N9 | Phoenicopterus chilensis | Phoenicopteriformes | OQ820949 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz, S.; Galdames, P.; Baumberger, C.; Gonzalez, M.A.; Rojas, C.; Oyarzun, C.; Orozco, K.; Mattar, C.; Freiden, P.; Sharp, B.; et al. Remote Sensing and Ecological Variables Related to Influenza A Prevalence and Subtype Diversity in Wild Birds in the Lluta Wetland of Northern Chile. Viruses 2023, 15, 1241. https://doi.org/10.3390/v15061241
Ruiz S, Galdames P, Baumberger C, Gonzalez MA, Rojas C, Oyarzun C, Orozco K, Mattar C, Freiden P, Sharp B, et al. Remote Sensing and Ecological Variables Related to Influenza A Prevalence and Subtype Diversity in Wild Birds in the Lluta Wetland of Northern Chile. Viruses. 2023; 15(6):1241. https://doi.org/10.3390/v15061241
Chicago/Turabian StyleRuiz, Soledad, Pablo Galdames, Cecilia Baumberger, Maria Antonieta Gonzalez, Camila Rojas, Cristobal Oyarzun, Katherinne Orozco, Cristian Mattar, Pamela Freiden, Bridgette Sharp, and et al. 2023. "Remote Sensing and Ecological Variables Related to Influenza A Prevalence and Subtype Diversity in Wild Birds in the Lluta Wetland of Northern Chile" Viruses 15, no. 6: 1241. https://doi.org/10.3390/v15061241
APA StyleRuiz, S., Galdames, P., Baumberger, C., Gonzalez, M. A., Rojas, C., Oyarzun, C., Orozco, K., Mattar, C., Freiden, P., Sharp, B., Schultz-Cherry, S., Hamilton-West, C., & Jimenez-Bluhm, P. (2023). Remote Sensing and Ecological Variables Related to Influenza A Prevalence and Subtype Diversity in Wild Birds in the Lluta Wetland of Northern Chile. Viruses, 15(6), 1241. https://doi.org/10.3390/v15061241